

Abstract
Background
Childhood obesity prevalence has increased worldwide and is an important risk factor for type 2 diabetes (T2D) and cardiovascular disease (CVD). The production of inflammatory adipokines by obese adipose tissue contributes to the development of T2D and CVD. While levels of circulating adipokines such as adiponectin and leptin have been established in obese children and adults, the expression of adiponectin and leptin receptors on circulating immune cells can modulate adipokine signalling, but has not been studied so far. Here, we aim to establish the expression of adiponectin and leptin receptors on circulating immune cells in obese children pre and post-lifestyle intervention compared to normal weight control children.
Methods
13 obese children before and after a 1-year lifestyle intervention were compared with an age and sex-matched normal weight control group of 15 children. Next to routine clinical and biochemical parameters, circulating adipokines were measured, and flow cytometric analysis of adiponectin receptor 1 and 2 (AdipoR1, AdipoR2) and leptin receptor expression on peripheral blood mononuclear cell subsets was performed.
Results
Obese children exhibited typical clinical and biochemical characteristics compared to controls, including a higher BMI-SD, blood pressure and circulating leptin levels, combined with a lower insulin sensitivity index (QUICKI). The 1-year lifestyle intervention resulted in stabilization of their BMI-SD. Overall, circulating leukocyte subsets showed distinct adipokine receptor expression profiles. While monocytes expressed high levels of all adipokine receptors, NK and iNKT cells predominantly expressed AdipoR2, and B-lymphocytes and CD4+ and CD8+ T-lymphocyte subsets expressed AdipoR2 as well as leptin receptor. Strikingly though, leukocyte subset numbers and adipokine receptor expression profiles were largely similar in obese children and controls. Obese children showed higher naïve B-cell numbers, and pre-intervention also higher numbers of immature transition B-cells and intermediate CD14++CD16+ monocytes combined with lower total monocyte numbers, compared to controls. Furthermore, adiponectin receptor 1 expression on nonclassical CD14+CD16++ monocytes was consistently upregulated in obese children pre-intervention, compared to controls. However, none of the differences in leukocyte subset numbers and adipokine receptor expression profiles between obese children and controls remained significant after multiple testing correction.
Conclusions
First, the distinct adipokine receptor profiles of circulating leukocyte subsets may partly explain the differential impact of adipokines on leukocyte subsets. Second, the similarities in adipokine receptor expression profiles between obese children and normal weight controls suggest that adipokine signaling in childhood obesity is primarily modulated by circulating adipokine levels, instead of adipokine receptor expression.
Introduction
Obese children often remain obese in adulthood, and are at risk for metabolic syndrome and cardiovascular disease later in life [1–3]. Over the last few decades, enhanced excretion of inflammatory adipose tissue derived proteins (adipokines) emerged as one of the mechanisms underlying the cardiometabolic sequelae in obesity [4,5]. The adipokine profile in childhood and adulthood obesity includes increased levels of the inflammatory adipokines chemerin and leptin, and decreased levels of the anti-inflammatory adiponectin, which together propagates systemic inflammation, insulin resistance and vascular dysfunction, as a precursor for cardiovascular disease [4,6–11].
The impact of circulating adipokines is not merely determined by plasma levels, but also orchestrated by differential adipokine receptor expression on target organs. For example, adiponectin receptor 1 (AdipoR1) and receptor 2 (AdipoR2) are significantly homologous (67% amino acid identity), and both serve as a receptor for globular and full-length adiponectin [12]. However, AdipoR1 is predominantly expressed in liver, skeletal muscle, macrophages and hypothalamus, while AdipoR2 is most abundant in liver, white adipose tissue, and the vasculature. The differential tissue distribution and downstream signalling pathways of AdipoR1 and AdipoR2 importantly contribute to the plethora of adiponectin’s biological actions [10]. Next to differences in tissue distribution, up/downregulation of adipokine receptor expression under specific conditions can also modulate adipokine effects. Natural Killer (NK) cells, for instance, critically depend on leptin receptor expression for their activation and function [13]. While leptin receptor-deficient mice showed impaired NK cell activity, leptin receptor expression was upregulated in rats with diet-induced obesity, apparently to compensate for decreased downstream signalling [14]. Taken together, both tissue distribution and disease-specific up/downregulation of adipokine receptors can modulate adipokine effects.
Whereas studying adipose tissue, liver and vascular distribution of adipokine receptors is precluded for medical ethical reasons, circulating leukocytes are readily available and play a pivotal role in systemic inflammation in obesity [15–18]. Here, we used recently available flow cytometry antibodies to study the expression of AdipoR1, AdipoR2 and leptin receptor on circulating leukocyte subsets, in addition to measuring circulating adipokine levels. Considering the pivotal role of adipokine signalling in obesity, we included obese children pre and post-lifestyle intervention, next to normal weight control children. This study thus aims to unravel the differential impact of circulating adipokine levels and adipokine receptor expression on adipokine signalling in childhood obesity.
Materials and methods
Patients
This observational cohort study included 15 lean control children and 13 obese children aged 4–18 years, all patients of the pediatric outpatient department of Meander Medical Centre in Amersfoort and Hospital Gelderse Vallei Hospital in Ede, The Netherlands. Body Mass Index standard deviation (BMI-SD) values were calculated based on results of the Fifth Dutch Growth Study [19]. Obesity was defined using established international age and sex-specific BMI cut-off points [20]. The obese patients were enrolled in an established 1-year multidisciplinary, multicomponent, family-based treatment programme developed in the Gelderse Vallei Hospital in Ede, The Netherlands [21]. Patients were included from 2010 to 2015. Anthropometric measurements, blood pressure (BP) measurements and laboratory samples were collected at baseline (lean controls, obese children), and at the end of the 1-year intervention program (obese children). The study was approved by the ethical committee of Wageningen UR (METC 12/26) and the University Medical Center Utrecht (METC 09/217K). Written informed consent was obtained from children older than 12 years and from the parents.
Clinical variables
Waist circumference was used as a marker of central adiposity and measured with a flexible tape to the nearest 0.1 cm at umbilicus height. Blood pressure (BP) was measured in supine position with an automated blood pressure monitor (Welch Allyn VSM 300, Skaneateles Falles, NY, USA) after 5 minutes of rest during a well visit in the outpatient clinic. A minimum of two BP measurements was performed, with an interval of at least one minute between the measurements. The mean of these two measurements was collected for data analysis. Blood pressure percentile scores were obtained according to the Fourth Report on BP in children [22].
Routine laboratory measurements
Routine laboratory testing included fasting glucose, insulin levels and lipid profiles (total cholesterol, high-density lipoprotein [HDL] cholesterol, low-density lipoprotein [LDL], and triglycerides), as well as samples for leukocyte differentiation, and alanine-aminotranspherase (ALT). The insulin sensitivity index (QUICKI) was calculated according to international standards [23].
Flow cytometry
Whole blood samples collected in sodium heparin tubes (BD vacutainer 367876) where spun down at room temperature, 160g for 10 minutes. Plasma was subsequently removed and stored at -80°C awaiting further use. Peripheral Blood mononuclear cells (PBMC) were isolated using Ficoll-Paque density gradient centrifugation and samples were stored in fetal bovine serum (FBS) (Biowest) supplemented with 10% DMSO (Sigma-Aldrich) at -150°C. All children donated 6–12ml whole blood, with 6–10 million PBMC per 6ml whole blood sample. Upon preparation for flow cytometry, the stored PBMC samples where thawed and washed in medium comprising of RPMI1640 supplemented with l-glutamate and 25 mM HEPES (Gibco), containing 2% FBS (Biowest) and penicillin/streptomycin (100 U/mL) (Invitrogen). Cells were spun down for 10 minutes at 280g at room temperature. All samples were stained for 20 minutes in the dark at 4°C and subsequently analyzed on the BD LSR Fortessa. For monocyte, NK cell, B-cell, T-helper, T-effector/memory and Treg phenotyping, 200.000 PBMCs were analyzed per sample. For iNKT cell phenotyping, 750.000 PBMCs were analyzed per sample. The following antibodies were used: CD3 AF700 (Biolegend, clone UCHT1), CD4 PerCP-Cy5.5 (BD, clone SK3), CD25 PE-Cy7 (BD, clone M-A251), CD45RO BV711 (Biolegend, clone UCHL1), CD127 BV421 (BD Horizon, clone HIL-7R-M21), CD8 V500 (BD, clone RPA-T8), Leptin receptor Alexaflour647 (BD,clone 52263), ADIPOR1 FITC (USBio, rabbit polyclonal antibody). ADIPOR2 PE (USBio, rabbit polyclonal antibody), CD27 APC-eFluor780 (eBioscience, clone O323), CD28 BV421 (BD Horizon, clone CD28.2), CCR6 PE-Cy7 (eBiosciences, clone R6H1), CXCR3 BV510 (Biolegend, clone GO25H7), CD16 V500 (BD Horizon, clone 3G8), CD56 PE-Cy7 (BD, clone NCAM16.2), CD1d tetramer BV421 (NIH, hCD1d-PBS-57), CD10 PE-Cy7 (BD, clone HI10A), CD19 APC-efluor 780 (eBioscience, clone HIB19), CD21 BV711 (BD, clone B-ly4), CD27 BV510 (BD Horizon, clone L128), CD38 PerCP-Cy5.5 (BD Pharmigen, clone HIT2). Leukocyte subset numbers were calculated using the differential blood count (e.g. number of CD14++CD16+ monocytes = (fraction of CD14++CD16+ monocytes / total monocytes) x differential blood count monocyte number). Gating strategy of the leukocyte subsets is shown in the S1 Fig.
Multiplex immune assay (MIA)
Plasma levels of adiponectin, chemerin and leptin were measured by a MIA using Luminex xMAP technology (xMAP, Luminex Austin TX USA) validated by the Laboratory of Translational Immunology, University Medical Center Utrecht [24]. Biorad FlexMAP3D (Biorad laboratories. Hercules USA) and xPONENT software version 4.2 (Luminex) were used for acquisition and data was analyzed by 5-parametric curve fitting using Bio-Plex Manager software, version 6.1.1 (Biorad).
Statistical analysis
Differences between groups were studied with an independent-sample Student’s t-test for normally distributed data, and with a Mann-Whitney U test for non-parametric comparisons. Multiple testing correction using the Benjamini and Hochberg False Discovery Rate (FDR) procedure was applied when assessing leukocyte subset numbers and comparing adipokine receptor expression of leukocyte subsets in lean versus obese children pre and post-intervention. Pearson’s correlation coefficients were calculated to determine the correlation between levels of circulating adipokines and the expression of adiponectin and leptin receptors on circulating immune cells. Statistical analyses were performed with the SPSS 22 statistical package (IBM SPSS Statistics Inc, Chicago, IL, USA).
Results
Circulating leukocyte subset numbers
Obese children exhibited typical clinical and biochemical characteristics compared to age and sex-matched normal weight controls, with a higher BMI-SD, waist circumference and blood pressure, combined with a lower insulin sensitivity index (QUICKI) and higher plasma levels of alanine aminotransferase, chemerin and leptin (Table 1). The obese children participated in an established lifestyle intervention program [21], which resulted in stabilization of their BMI-SD and other clinical and biochemical characteristics, and enabled sampling pre and post-lifestyle intervention (Table 1).
Table 1. Patient characteristics.
| Lean controls (n = 15) | Obese—pre (n = 13) | Obese—post (n = 13 | |
|---|---|---|---|
| Girls (number, %) | 8 (53.3) | 7 (53.8) | 7 (53.8) |
| Age (years) | 11.7 ± 2.9 | 10.7 ± 3.9 | 12.0 ± 3.9 |
| BMI | 18.5 ± 2.6 *# | 27.9 ± 5.2 * | 28.2 ± 5.5 # |
| BMI-SD | 0.5 ± 0.9 *# | 3.3 ± 0.7 * | 3.1 ± 0.7 # |
| Waist (cm) | 64.5 ± 7.5 *# | 94.4 ± 16.8 * | 98.4 ± 17.0 # |
| Systolic blood pressure (SBP) | 102.2 ± 12.7 *# | 113.2 ± 12.2 * | 114.8 ± 11.7 # |
| SBP percentile | 39.9 ± 23.6 *# | 65.7 ± 25.1 * | 65.5 ± 28.4 # |
| Diastolic blood pressure (DBP) | 57.7 ± 8.2 *# | 67.2 ± 9.5 * | 69.4 ± 10.5 # |
| DBP percentile | 35.9 ± 19.9 *# | 62.6 ± 23.9 * | 65.4 ± 27.1 # |
| QUICKI | 0.4 ± 0.0 *# | 0.3 ± 0.0 * | 0.3 ± 0.0 # |
| Alanine aminotransferase (U/l) | 18.0 (14.5–21.8) *# | 29.5 (22.2–53.9) * | 30.3 (22.9–41.2) # |
| Triacylglycerol (mmol/l) | 0.6 ± 0.3 * | 1.0 ± 0.4 * | 1.0 ± 0.7 |
| Total cholesterol (mmol/l) | 4.0 ± 0.8 | 4.0 ± 0.8 | 3.9 ± 0.9 |
| HDL-cholesterol (mmol/l) | 1.7 ± 0.9 | 1.2 ± 0.3 | 1.3 ± 0.2 |
| LDL-cholesterol (mmol/l) | 2.2 ± 0.7 | 2.3 ± 0.7 | 2.2 ± 0.8 |
| Adiponectin (ug/ml) | 18.9 (15.6–26.5) | 12.9 (9.5–21.3) | 13.6 (10.4–19.8) |
| Chemerin (ug/ml) | 1.2 (1.0–1.7) *# | 1.9 (1.8–2.7) * | 2.3 (1.8–2.5) # |
| Leptin (ng/ml) | 125 (16–195) *# | 399 (219–692) * | 411 (308–600) # |
Clinical characteristics and laboratory parameters for lean controls versus obese children pre-lifestyle intervention (pre) and post-lifestyle intervention (post). Normally distributed data are shown as mean ± SD, non-parametric data as median (interquartile range).
* p<0.05 for lean controls compared to obese-pre.
# p<0.05 for lean controls versus obese-post.
Circulating leukocytes were analysed with multi-parameter flow cytometry, and leukocyte subsets were gated according to international standards (Fig 1, S1 Fig) [25]. Overall, leukocyte subset numbers were comparable between obese children and lean controls. However, obese children pre and post-intervention showed higher naïve B-cell numbers, and obese children pre-intervention showed higher numbers of immature transition B-cells and intermediate CD14++CD16+ monocytes than lean controls (Table 2). Obese children pre-intervention additionally showed lower total monocyte numbers than lean controls. Notably, differences in leukocyte subset numbers did not survive multiple testing correction.
Fig 1. Adipokine receptor expression on leukocyte subsets.

The first column illustrates the gating strategy for the leukocyte subsets. The other columns show representative histograms of the AdipoR1, AdipoR2 and leptin receptor expression of the different leukocyte subsets.
Table 2. Leukocyte subset numbers.
| Lean controls | Obese-pre | Obese-post | |
|---|---|---|---|
| Innate immunity | |||
| Monocytes (total) | 400 (300–600) * | 300 (300–400) * | 400 (300–500) |
| CD14++CD16- | 352 (255–542) | 244 (196–303) | 330 (245–374) |
| CD14++CD16+ | 15 (11–19)* | 16 (13–22)* | 18 (13–39) |
| CD14+CD16++ | 37 (33–94) | 39 (30–79) | 40 (36–69) |
| Natural Killer cells (CD16+CD56+) | 150 (80–211) | 224 (116–321) | 226 (134–287) |
| CD16+CD56++ | 8.7 (5.5–12) | 8.0 (4.9–13) | 9.3 (5.8–12) |
| CD16-CD56++ | 4.9 (2.8–8.0) | 5.5 (3.4–8.1) | 5.0 (4.4–8.9) |
| Bridging immunity | |||
| Natural Killer T cells | 1.2 (0.6–2.2) | 1.4 (0.9–3.9) | 1.2 (1.0–3.6) |
| Adaptive immunity | |||
| B cells | |||
| Naive (CD10-CD27-) | 178 (132–270) *# | 255 (189–454) * | 316 (224–454) # |
| Memory (CD10-CD27+) | 48 (41–73) | 69 (45–113) | 74 (56–117) |
| Immature transition (CD10+CD27+) | 10 (5.4–17) * | 23 (9.3–39) * | 15 (10–31) |
| CD4+ T helper cells | |||
| CD45RO- CXCR3- | 511 (451–700) | 639 (401–750) | 624 (334–937) |
| CD45RO- CXCR3+ | 58 (40–176) | 58 (32–110) $ | 85 (68–162) $ |
| CD45RO+ CXCR3- | 140 (115–196) | 151 (125–227) | 207 (103–251) |
| CD45RO+ CXCR3+ | 111 (84–134) | 67 (43–115) | 84 (57–117) |
| CD8+ cytotoxic T cells | |||
| CD45RO- CCR7- | 76 (37–138) | 76 (43–178) | 94 (29–161) |
| CD45RO- CCR7+ | 284 (275–358) | 355 (272–434) | 294 (238–477) |
| CD45RO+ CCR7- | 82 (44–85) | 56 (38–91) | 52 (42–73) |
| CD45RO+ CCR7+ | 15 (12–19) | 13 (8.6–30) | 18 (13–22) |
| Regulatory T cells (CD25+CD127-) | 26 (14–35) | 21 (16–28) | 22 (13–33) |
Leukocyte subset numbers (x 106) of lean controls compared to obese children pre-lifestyle intervention (pre) and post-lifestyle intervention (post). Data are presented as median (interquartile range).
* p<0.05 for lean controls compared to obese-pre.
# p<0.05 for lean controls versus obese-post.
$ p<0.05 for obese-pre compared to obese-post.
Adipokine receptor expression on leukocyte subsets
Focussing on the innate immune cells, monocytes expressed high levels of adipokine receptors compared to other circulating leukocyte subsets (Fig 1). Whereas nonclassical CD14+CD16++ monocytes particularly expressed high levels of AdipoR1, all monocyte subsets expressed AdipoR2, and classical CD14++CD16- and intermediate CD14++CD16+ monocytes particularly expressed high leptin receptor levels. In contrast, Natural Killer (NK) cells and invariant Natural Killer T-cells (iNKT) predominantly showed AdipoR2 expression, next to discrete AdipoR1 expression by CD16+ NK cells (Fig 1, Tables 3, 4 and 5).
Table 3. Adiponectin receptor 1 expression.
| Lean controls | Obese-pre | Obese-post | |
|---|---|---|---|
| Innate immunity | |||
| Monocytes (total) | 21 (18–31) | 31 (18–36) | 25 (20–29) |
| CD14++CD16- | 10 (8.8–16) # | 8.2 (4.6–27) | 7.7 (3.7–11) # |
| CD14++CD16+ | 82 (68–88) | 90 (81–94) | 84 (73–93) |
| CD14+CD16++ | 84 (71–92) * | 94 (77–98) * | 91 (85–96) |
| Natural Killer cells (CD16+CD56+) | 82 (56–92) | 86 (76–95) | 88 (78–95) |
| CD16+CD56++ | 41 (29–63) | 51 (39–58) | 56 (44–64) |
| CD16-CD56++ | 1.6 (0.5–3.1) | 0.9 (0.1–4.6) | 0.8 (0.4–1.8) |
| Bridging immunity | |||
| Natural Killer T cells | 7.6 (4.1–25) | 12 (4.1–18) | 6.9 (5.0–15) |
| Adaptive immunity | |||
| B cells | |||
| Naive (CD10-CD27-) | 1.1 (0.6–2.2) # | 0.8 (0.5–2.0) | 0.5 (0.4–1.0) # |
| Memory (CD10-CD27+) | 5.9 (3.8–6.6) | 5.6 (3.8–8.8) | 6.4 (3.5–9.6) |
| Immature transition (CD10+CD27+) | 0.0 (0.0–0.2) * | 0.7(0.3–1.0) *$ | 0.1 (0.0–0.5) $ |
| CD4+ T helper cells | |||
| CD45RO- CXCR3- | 5.2 (1.6–21) | 3.3 (1.0–22) | 3.6 (1.6–19) |
| CD45RO- CXCR3+ | 64 (45–92) | 47 (37–71) | 63 (49–81) |
| CD45RO+ CXCR3- | 6.1 (2.1–12) | 4.7 (1.6–12) | 6.6 (2.2–11) |
| CD45RO+ CXCR3+ | 24 (12–38) | 21 (12–42) | 41 (16–56) |
| CD8+ cytotoxic T cells | |||
| CD45RO- CCR7- | 46 (41–55) | 51 (42–61) | 50 (42–61) |
| CD45RO- CCR7+ | 60 (55–74) | 54 (45–66) | 53 (46–68) |
| CD45RO+ CCR7- | 34(30–43) | 36 (31–52) | 38 (34–43) |
| CD45RO+ CCR7+ | 70 (66–77) | 68 (52–90) | 72 (67–87) |
| Regulatory T cells (CD25+CD127-) | 3.9 (1.6–14) | 4.0 (1.0–9.0) | 4.8 (1.6–9.7) |
Percentage expression of adiponectin receptor 1 on leukocyte subsets of lean controls compared to obese children pre-lifestyle intervention (pre) and post-lifestyle intervention (post). Data are presented as median (interquartile range).
* p<0.05 for lean controls compared to obese-pre.
# p<0.05 for lean controls versus obese-post.
$ p<0.05 for obese-pre compared to obese-post.
Table 4. Adiponectin receptor 2 expression.
| Lean controls | Obese-pre | Obese-post | |
|---|---|---|---|
| Innate immunity | |||
| Monocytes (total) | 60 (32–67) | 68 (46–80) | 57 (43–70) |
| CD14++CD16- | 59 (29–66) | 60 (45–80) | 54 (41–67) |
| CD14++CD16+ | 79 (60–88) | 80 (67–93) | 75 (59–86) |
| CD14+CD16++ | 60 (44–70) | 63 (46–80) | 62 (44–68) |
| Natural Killer cells (CD16+CD56+) | 67 (62–80) # | 61 (53–72) | 62 (53–66) # |
| CD16+CD56++ | 98 (97–100) | 97 (95–99) | 98 (93–99) |
| CD16-CD56++ | 99 (96–100) | 97 (95–99) | 98 (97–100) |
| Bridging immunity | |||
| Natural Killer T cells | 77 (70–89) | 70 (61–74) | 77 (62–82) |
| Adaptive immunity | |||
| B cells | |||
| Naive (CD10-CD27-) | 38 (35–46) | 36 (32–41) | 35 (24–40) |
| Memory (CD10-CD27+) | 45 (39–52) | 43 (36–46) | 41 (19–47) |
| Immature transition (CD10+CD27+) | 74 (61–81) | 72 (58–87) | 77 (57–82) |
| CD4+ T helper cells | |||
| CD45RO- CXCR3- | 27 (25–32) # | 25 (20–38) | 25 (13–27) # |
| CD45RO- CXCR3+ | 54 (46–88) | 49 (40–58) | 50 (43–67) |
| CD45RO+ CXCR3- | 34 (31–39) | 32 (28–41) | 33 (14–37) |
| CD45RO+ CXCR3+ | 43 (36–62) | 40 (33–54) | 47 (33–60) |
| CD8+ cytotoxic T cells | |||
| CD45RO- CCR7- | 29 (11–34) | 31 (24–44) | 29 (12–33) |
| CD45RO- CCR7+ | 26 (22–48) | 24 (19–44) | 25 (20–38) |
| CD45RO+ CCR7- | 31 (13–41) | 36 (30–44) | 30 (7.6–35) |
| CD45RO+ CCR7+ | 58 (52–76) | 55 (35–80) | 58 (52–77) |
| Regulatory T cells (CD25+CD127-) | 23 (21–27) | 20 (15–27) | 22 (19–28) |
Percentage expression of adiponectin receptor 2 on leukocyte subsets of lean controls compared to obese children pre-lifestyle intervention (pre) and post-lifestyle intervention (post). Data are presented as median (interquartile range).
# p<0.05 for lean controls versus obese-post.
Table 5. Leptin receptor expression.
| Lean controls | Obese-pre | Obese-post | |
|---|---|---|---|
| Innate immunity | |||
| Monocytes (total) | 96 (95–98) | 97 (92–98) | 97 (94–98) |
| CD14++CD16- | 99 (99–100) | 100 (99–100) | 100 (99–100) |
| CD14++CD16+ | 100 (99–100) | 100 (99–100) | 100 (99–100) |
| CD14+CD16++ | 70 (61–81) | 75 (65–84) | 77 (77–81) |
| Natural Killer cells (CD16+CD56+) | 8.0 (5.3–12) | 8.8 (3.3–18) | 6.1 (4.4–10) |
| CD16+CD56++ | 7.2 (5.3–9.6) | 5.7 (3.0–14) | 6.2 (3.0–9.7) |
| CD16-CD56++ | 6.6 (5.9–7.7) | 5.3 (3.1–7.1) | 6.4 (2.9–7.8) |
| Bridging immunity | |||
| Natural Killer T cells | 11 (5.8–19) | 13 (5.2–32) | 9.8 (8.7–18) |
| Adaptive immunity | |||
| B cells | |||
| Naive (CD10-CD27-) | 51 (44–56) | 51 (46–58) | 47 (46–55) |
| Memory (CD10-CD27+) | 60 (57–65) | 63 (54–65) | 57 (50–62) |
| Immature transition (CD10+CD27+) | 46 (39–54) | 50 (35–57) | 46 (39–51) |
| CD4+ T helper cells | |||
| CD45RO- CXCR3- | 31 (28–40) | 29 (24–41) | 28 (24–36) |
| CD45RO- CXCR3+ | 69 (59–93) | 62 (50–73) | 71 (64–80) |
| CD45RO+ CXCR3- | 26 (22–29) | 25 (21–26) | 24 (21–29) |
| CD45RO+ CXCR3+ | 42 (29–50) | 41 (28–49) $ | 55 (44–66) $ |
| CD8+ cytotoxic T cells | |||
| CD45RO- CCR7- | 22 (16–26) | 22 (17–34) | 25 (19–29) |
| CD45RO- CCR7+ | 41 (31–52) | 39 (31–52) | 38 (32–48) |
| CD45RO+ CCR7- | 21 (19–32) | 24 (19–32) | 28 (24–31) |
| CD45RO+ CCR7+ | 65 (56–72) | 67 (48–87) | 69 (61–80) |
| Regulatory T cells (CD25+CD127-) | 10 (7.8–17) | 12 (5.9–15) | 13 (11–17) |
Percentage expression of leptin receptor on leukocyte subsets of lean controls compared to obese children pre-lifestyle intervention (pre) and post-lifestyle intervention (post). Data are presented as median (interquartile range).
$ p<0.05 for obese-pre compared to obese-post.
Considering the adaptive immune cells, AdipoR1 expression appeared to be low, in general. Only part of the CD4+ CXCR3+ T-cells and CD8+ T-cells and a discrete subset of regulatory T-cells showed AdipoR1 expression. In contrast, AdipoR2 was particularly expressed by B-cells, next to a subset of CD4+ CXCR3+ T-cells and a subset of CD8+ CD45RO+ (memory) T-cells. Finally, leptin receptor was predominantly expressed by B-cells, and a subset of CD4+ CD45RO- (non-memory) CXCR3+ T-cells and of CD8+ CD45RO+ (memory) CCR7+ T-cells and regulatory T-cells (Fig 1, Tables 3, 4 and 5).
Taken together, circulating leukocyte subsets showed distinct adipokine receptor expression profiles. Monocytes generally expressed high levels of all adipokine receptors, while other leukocyte subsets showed a different pattern. NK and iNKT cells predominantly expressed AdipoR2, while B-lymphocytes and CD4+ and CD8+ T-lymphocyte subsets expressed AdipoR2 as well as leptin receptor. Notably, regulatory T-cells showed little adipokine receptor expression, but a discrete subset of regulatory T-cells expressed a combination of adipokine receptors (Figure in S2 Fig).
Adipokine receptor expression in lean and obese children
In order to establish differences in adipokine receptor expression between obese children pre and post-intervention and normal weight controls, we studied differences in expression percentages (Tables 3, 4 and 5), as well as median fluorescence intensities (MFI) (S1–S3 Tables). In general, obese children pre and post-intervention and normal weight controls showed similar adipokine receptor expression on leukocyte subsets. In fact, none of the observed differences in adipokine receptor expression between obese children and lean controls remained significant after multiple testing correction.
As an alternative strategy to discriminate between random deviations and potentially relevant differences, we focused on differences in adipokine receptor expression that were consistent in percentages as well as fluorescence intensities. AdipoR1 expression on nonclassical CD14+CD16++ monocytes was consistently upregulated in percentages and MFI in obese children pre-intervention, compared to lean controls (Table 3 and S1 Table). The other leukocyte subsets did not show consistent differences in AdipoR1, AdipoR2 or leptin receptor expression between obese children and lean controls.
Circulating adipokines and adipokine receptor expression
Finally, we wondered whether high circulating adipokine levels were associated with alterations in adipokine receptor expression, which could modulate adipokine signalling. First, we studied the relationship between circulating adipokine levels and adipokine receptor expression on monocytes. Neither did we observe a correlation between circulating adiponectin levels and AdipoR1/AdipoR2 expression (Fig 2A and 2B), nor a correlation between circulating leptin levels and leptin receptor expression (Fig 2C, Fig B in S3 Fig). Second, screening other leukocyte subsets did not yield a correlation between circulating adipokine levels and adipokine receptor expression either (Fig A in S3 Fig).
Fig 2. Circulating adipokine levels and monocyte adipokine receptor expression.
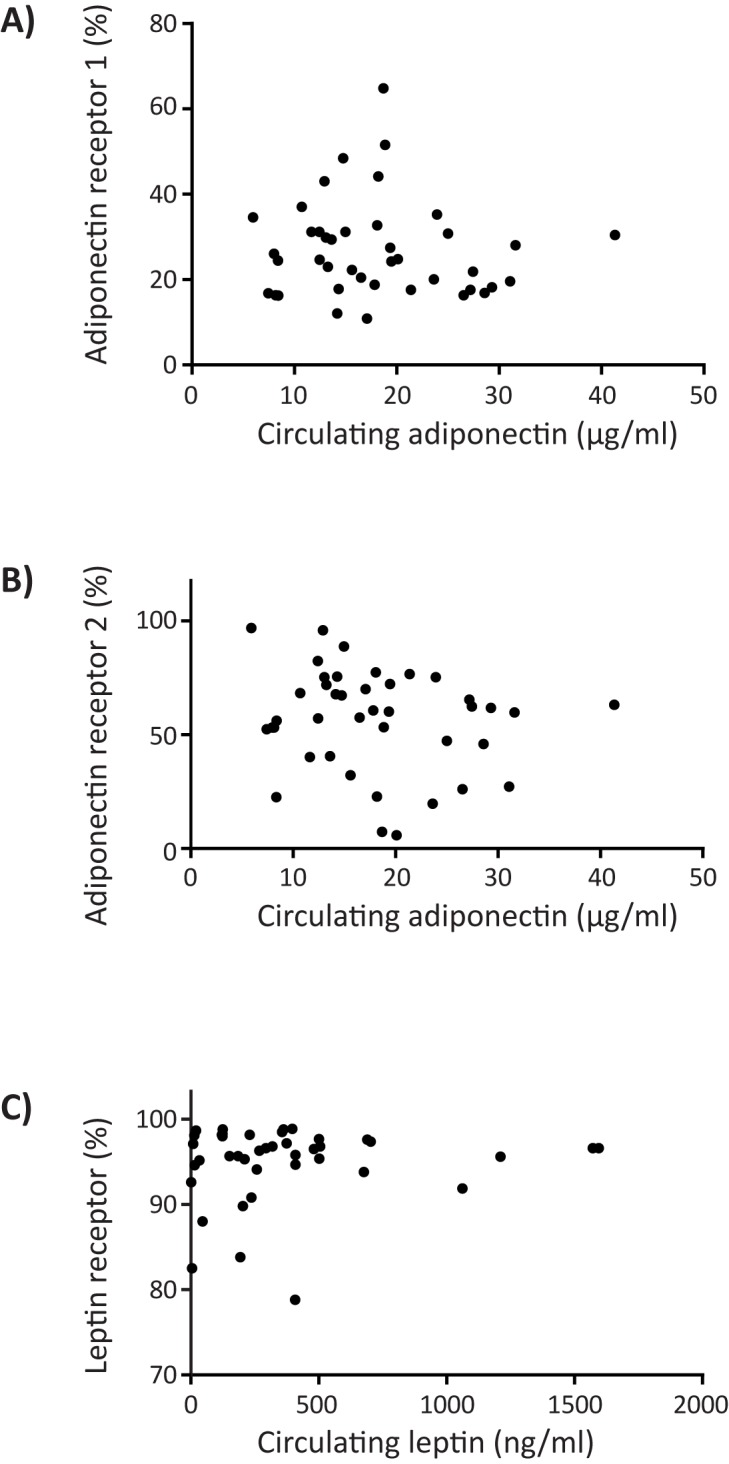
(A) Circulating adiponectin levels versus percentage of AdipoR1 expressing monocytes (Pearson’s correlation coefficient -0.079, p = 0.623). (B) Circulating adiponectin levels versus percentage of AdipoR2 expressing monocytes (Pearson’s correlation coefficient -0.178, p = 0.265). (C) Circulating leptin levels versus percentage of leptin receptor expressing monocytes (Pearson’s correlation coefficient 0.107, p = 0.505).
Discussion
Over the last decades, enhanced circulating leptin levels and decreased adiponectin levels emerged as one of the mechanisms underlying the cardiometabolic sequelae in obesity [4,5,11]. This study aimed to establish the expression of adiponectin and leptin receptors on circulating immune cell subsets in obese children and normal weight controls, since adipokine receptor expression on circulating leukocytes can modulate adipokine signalling and obesity-induced systemic inflammation [15,16]. Here, we will discuss our two main findings.
First, circulating leukocyte subsets showed distinct adipokine receptor expression profiles, which may partly explain the differential impact of adipokines on leukocyte subsets. Whereas enhanced AdipoR1/R2 expression on the myeloid cell lineage has for example been associated with anti-inflammatory (M2) macrophage polarization and suppression of foam cell formation [26,27], the decreased AdipoR1 expression on classical CD14++CD16- monocytes may be involved in their inflammatory fate and pivotal role in the development of cardiovascular disease [28,29]. Another example is the discrete subpopulation of Tregs expressing a combination of leptin receptor and adiponectin receptors (S2 Fig). Considering the role of peroxisome-proliferator-activated receptor γ (PPARγ) induced adiponectin signalling in adipose tissue differentiation of Tregs [30,31], and the pivotal role of leptin in T-cell differentiation [32–34], it is tempting to speculate that circulating leptin and adiponectin receptor positive Tregs represent recirculating adipose tissue Tregs. Taken together, our data provide an exciting starting point for future studies to the role of adipokine receptors in leukocyte differentiation and function.
Second, we observed largely similar leukocyte subset numbers and adipokine receptor expression profiles in obese children and controls. The high naive CD10-CD27- B-cell numbers in obese children pre and post-lifestyle intervention appear to be an exception to that. Interestingly, our findings correspond with recent studies observing high naive B-cell numbers in obese adults [35,36]. The high naïve B-cell numbers may be explained by leptin-induced B-cell hyperstimulation, which can impair B-cell function [35,36]. Indeed, B-cell responses to vaccination can be impaired in obesity [35,37]. With respect to the similar adipokine receptor expression profiles of obese children and lean controls, our findings suggest that in childhood obesity, adipokine signalling in circulating leukocytes is primarily modulated by circulating adipokine levels, instead of adipokine receptor expression.
Our study has a few limitations that have to be taken into account. Storage or freeze-thawing of the peripheral blood mononuclear cells may have neutralized differences in adipokine receptor expression between obese children and controls. Notably, differences in adipokine receptor expression between leukocyte subsets were preserved, which argues against significant storage or freeze-thawing effects. Next, while adipokine receptor expression in liver, skeletal muscle, adipose tissue and other tissues plays in important role in adipokine signalling as well, medical ethical reasons precluded tissue collection. Importantly, our observations in circulating leukocyte subsets do not extend to other tissues. Finally, our study may have been underpowered to identify subtle differences in adipokine receptor expression due to relatively small patient numbers. In obese adults, lymphocyte AdipoR1 and AdipoR2 mRNA expression was reduced compared to anorexic adults [38]. Likewise, reduced monocyte AdipoR1 and AdipoR2 protein expression was observed in obese adults with coronary artery disease, compared to obese adults without cardiovascular disease [39]. However, the results in these studies may have been distorted due to the fact that multiple testing corrections were not applied and nor differentiation was made between functionally distinct leukocyte subsets.
In conclusion, our results cannot exclude subtle differences in adipokine receptor expression, but suggest that adipokine signalling in circulating leukocytes in childhood obesity is primarily modulated by altered adipokine levels.
Supporting information
First, forward scatter (FSC) and sideward scatter (SSC) profiles were used to roughly distinguish monocytes and lymphocytes/NK cells. Second, doublet cells were excluded using FSC-Area (FSC-A) and FSC-Height (FSC-H) gating. Finally, leukocyte subsets were gated on their marker expression.
(TIF)
Focussing on the CD25hiCD127lowCD4+ T-cells (regulatory T-cells), the adiponectin receptor 1-positive subset also expresses the leptin receptor.
(TIF)
(A) Circulating adiponectin levels versus percentage of adiponectin receptor 2 expressing iNKT cells (Pearson’s correlation coefficient 0.048, p = 0.764), and circulating leptin levels versus percentage of leptin receptor expressing regulatory T-cells (Pearson’s correlation coefficient -0.130, p = 0.416). (B) Circulating adiponectin levels versus median fluorescence intensity (MFI) of adiponectin receptor 1 and 2 expression (Pearson’s correlation coefficient AdipoR1 0.052, p = 0.746; AdipoR2–0.172, p = 0.284), and circulating leptin levels versus median fluorescence intensity (MFI) of leptin receptor expression (Pearson’s correlation coefficient 0.126, p = 0.431).
(TIF)
(DOC)
(DOC)
(DOC)
(SAV)
Acknowledgments
The authors thank the paediatricians of the Meander Medical Center (Amersfoort, the Netherlands) and Hospital Gelderse Vallei (Ede, the Netherlands) for their help with patient recruitment.
The SPSS-database used for this study is attached as supplemental material (S1 Data).
Abbreviations
- AdipoR1/2
Adiponectin receptor 1/2
- BMI-SD
Body mass index standard deviation for age and sex
- BP
Blood pressure
- CVD
Cardiovascular disease
- HDL
High-density lipoprotein
- iNKT cells
invariant Natural Killer T-cells
- LDL
Low-density lipoprotein
- MFI
Median fluorescence intensity
- NK cells
Natural Killer cells
- PBMC
Peripheral blood mononuclear cells
- QUICKI
Quantitative insulin sensitivity index
- T2D
Type 2 diabetes
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This research was performed within the framework of the Center for Translational Molecular Medicine (CTMM; www.ctmm.nl), project Circulating Cells (grant 01C-102), and supported by the Dutch Heart Foundation, the UMC Utrecht Vascular Prevention Project and the Wilhelmina Children’s Hospital Research Fund. Henk Schipper was supported by a Fellowship Clinical Research Talent of the University Medical Center Utrecht. Genoveva Keustermans was supported by the Technology Foundation STW (Stichting voor de Technische Wetenschappen).
References
- 1.Twig G, Yaniv G, Levine H, Leiba A, Goldberger N, Derazne E, et al. (2016) Body-Mass Index in 2.3 Million Adolescents and Cardiovascular Death in Adulthood. N Engl J Med 374: 2430–2440. doi: 10.1056/NEJMoa1503840 [DOI] [PubMed] [Google Scholar]
- 2.Baker JL, Olsen LW, Sorensen TI (2007) Childhood body-mass index and the risk of coronary heart disease in adulthood. N Engl J Med 357: 2329–2337. doi: 10.1056/NEJMoa072515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lloyd LJ, Langley-Evans SC, McMullen S (2012) Childhood obesity and risk of the adult metabolic syndrome: a systematic review. Int J Obes (Lond) 36: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ouchi N, Parker JL, Lugus JJ, Walsh K (2011) Adipokines in inflammation and metabolic disease. Nat Rev Immunol 11: 85–97. doi: 10.1038/nri2921 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Freitas Lima LC, Braga VA, do Socorro de Franca Silva M, Cruz JC, Sousa Santos SH, de Oliveira Monteiro MM, et al. (2015) Adipokines, diabetes and atherosclerosis: an inflammatory association. Front Physiol 6: 304 doi: 10.3389/fphys.2015.00304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Schipper HS, Nuboer R, Prop S, van den Ham HJ, de Boer FK, Kesmir C, et al. (2012) Systemic inflammation in childhood obesity: circulating inflammatory mediators and activated CD14(++) monocytes. Diabetologia 55: 2800–2810. doi: 10.1007/s00125-012-2641-y [DOI] [PubMed] [Google Scholar]
- 7.Bonomini M, Pandolfi A (2016) Chemerin in renal dysfunction and cardiovascular disease. Vascul Pharmacol 77: 28–34. doi: 10.1016/j.vph.2015.10.007 [DOI] [PubMed] [Google Scholar]
- 8.Rourke JL, Dranse HJ, Sinal CJ (2013) Towards an integrative approach to understanding the role of chemerin in human health and disease. Obes Rev 14: 245–262. doi: 10.1111/obr.12009 [DOI] [PubMed] [Google Scholar]
- 9.Abella V, Scotece M, Conde J, Pino J, Gonzalez-Gay MA, Gomez-Reino JJ, et al. (2017) Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat Rev Rheumatol 13: 100–109. doi: 10.1038/nrrheum.2016.209 [DOI] [PubMed] [Google Scholar]
- 10.Yamauchi T, Kadowaki T (2013) Adiponectin receptor as a key player in healthy longevity and obesity-related diseases. Cell Metab 17: 185–196. doi: 10.1016/j.cmet.2013.01.001 [DOI] [PubMed] [Google Scholar]
- 11.Ohashi K, Shibata R, Murohara T, Ouchi N (2014) Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol Metab 25: 348–355. doi: 10.1016/j.tem.2014.03.009 [DOI] [PubMed] [Google Scholar]
- 12.Yamauchi T, Iwabu M, Okada-Iwabu M, Kadowaki T (2014) Adiponectin receptors: a review of their structure, function and how they work. Best Pract Res Clin Endocrinol Metab 28: 15–23. doi: 10.1016/j.beem.2013.09.003 [DOI] [PubMed] [Google Scholar]
- 13.Tian Z, Sun R, Wei H, Gao B (2002) Impaired natural killer (NK) cell activity in leptin receptor deficient mice: leptin as a critical regulator in NK cell development and activation. Biochem Biophys Res Commun 298: 297–302. [DOI] [PubMed] [Google Scholar]
- 14.Nave H, Mueller G, Siegmund B, Jacobs R, Stroh T, Schueler U, et al. (2008) Resistance of Janus kinase-2 dependent leptin signaling in natural killer (NK) cells: a novel mechanism of NK cell dysfunction in diet-induced obesity. Endocrinology 149: 3370–3378. doi: 10.1210/en.2007-1516 [DOI] [PubMed] [Google Scholar]
- 15.Procaccini C, De Rosa V, Galgani M, Carbone F, La Rocca C, Formisano L, et al. (2013) Role of adipokines signaling in the modulation of T cells function. Front Immunol 4: 332 doi: 10.3389/fimmu.2013.00332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sanchez-Margalet V, Martin-Romero C, Santos-Alvarez J, Goberna R, Najib S, Gonzalez-Yanes C (2003) Role of leptin as an immunomodulator of blood mononuclear cells: mechanisms of action. Clin Exp Immunol 133: 11–19. doi: 10.1046/j.1365-2249.2003.02190.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pang TT, Narendran P (2008) The distribution of adiponectin receptors on human peripheral blood mononuclear cells. Ann N Y Acad Sci 1150: 143–145. doi: 10.1196/annals.1447.021 [DOI] [PubMed] [Google Scholar]
- 18.Tsiotra PC, Pappa V, Raptis SA, Tsigos C (2000) Expression of the long and short leptin receptor isoforms in peripheral blood mononuclear cells: implications for leptin's actions. Metabolism 49: 1537–1541. doi: 10.1053/meta.2000.18519 [DOI] [PubMed] [Google Scholar]
- 19.Schonbeck Y, Talma H, van Dommelen P, Bakker B, Buitendijk SE, Hirasing RA, et al. (2011) Increase in prevalence of overweight in Dutch children and adolescents: a comparison of nationwide growth studies in 1980, 1997 and 2009. PLoS One 6: e27608 doi: 10.1371/journal.pone.0027608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cole TJ, Bellizzi MC, Flegal KM, Dietz WH (2000) Establishing a standard definition for child overweight and obesity worldwide: international survey. BMJ 320: 1240–1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.van Hoek E, Bouwman LI, Koelen MA, Lutt MA, Feskens EJ, Janse AJ (2016) Development of a Dutch intervention for obese young children. Health Promot Int. [DOI] [PubMed] [Google Scholar]
- 22.National High Blood Pressure Education Program Working Group on High Blood Pressure in C, Adolescents (2004) The fourth report on the diagnosis, evaluation, and treatment of high blood pressure in children and adolescents. Pediatrics 114: 555–576. [PubMed] [Google Scholar]
- 23.Katz A, Nambi SS, Mather K, Baron AD, Follmann DA, Sullivan G, et al. (2000) Quantitative insulin sensitivity check index: a simple, accurate method for assessing insulin sensitivity in humans. J Clin Endocrinol Metab 85: 2402–2410. doi: 10.1210/jcem.85.7.6661 [DOI] [PubMed] [Google Scholar]
- 24.Schipper HS, de Jager W, van Dijk ME, Meerding J, Zelissen PM, Adan RA, et al. (2010) A multiplex immunoassay for human adipokine profiling. Clin Chem 56: 1320–1328. doi: 10.1373/clinchem.2010.146118 [DOI] [PubMed] [Google Scholar]
- 25.Maecker HT, McCoy JP, Nussenblatt R (2012) Standardizing immunophenotyping for the Human Immunology Project. Nat Rev Immunol 12: 191–200. doi: 10.1038/nri3158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tian L, Luo N, Zhu X, Chung BH, Garvey WT, Fu Y (2012) Adiponectin-AdipoR1/2-APPL1 signaling axis suppresses human foam cell formation: differential ability of AdipoR1 and AdipoR2 to regulate inflammatory cytokine responses. Atherosclerosis 221: 66–75. doi: 10.1016/j.atherosclerosis.2011.12.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.van Stijn CM, Kim J, Lusis AJ, Barish GD, Tangirala RK (2015) Macrophage polarization phenotype regulates adiponectin receptor expression and adiponectin anti-inflammatory response. FASEB J 29: 636–649. doi: 10.1096/fj.14-253831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Berg KE, Ljungcrantz I, Andersson L, Bryngelsson C, Hedblad B, Fredrikson GN, et al. (2012) Elevated CD14++CD16- monocytes predict cardiovascular events. Circ Cardiovasc Genet 5: 122–131. doi: 10.1161/CIRCGENETICS.111.960385 [DOI] [PubMed] [Google Scholar]
- 29.Weber C, Shantsila E, Hristov M, Caligiuri G, Guzik T, Heine GH, et al. (2016) Role and analysis of monocyte subsets in cardiovascular disease. Joint consensus document of the European Society of Cardiology (ESC) Working Groups "Atherosclerosis & Vascular Biology" and "Thrombosis". Thromb Haemost 116: 626–637. doi: 10.1160/TH16-02-0091 [DOI] [PubMed] [Google Scholar]
- 30.Cipolletta D (2014) Adipose tissue-resident regulatory T cells: phenotypic specialization, functions and therapeutic potential. Immunology 142: 517–525. doi: 10.1111/imm.12262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cipolletta D, Feuerer M, Li A, Kamei N, Lee J, Shoelson SE, et al. (2012) PPAR-gamma is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature 486: 549–553. doi: 10.1038/nature11132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Saucillo DC, Gerriets VA, Sheng J, Rathmell JC, Maciver NJ (2014) Leptin metabolically licenses T cells for activation to link nutrition and immunity. J Immunol 192: 136–144. doi: 10.4049/jimmunol.1301158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Reis BS, Lee K, Fanok MH, Mascaraque C, Amoury M, Cohn LB, et al. (2015) Leptin receptor signaling in T cells is required for Th17 differentiation. J Immunol 194: 5253–5260. doi: 10.4049/jimmunol.1402996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Naylor C, Petri WA Jr. (2016) Leptin Regulation of Immune Responses. Trends Mol Med 22: 88–98. doi: 10.1016/j.molmed.2015.12.001 [DOI] [PubMed] [Google Scholar]
- 35.Frasca D, Ferracci F, Diaz A, Romero M, Lechner S, Blomberg BB (2016) Obesity decreases B cell responses in young and elderly individuals. Obesity (Silver Spring) 24: 615–625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kosaraju R, Guesdon W, Crouch MJ, Teague HL, Sullivan EM, Karlsson EA, et al. (2017) B Cell Activity Is Impaired in Human and Mouse Obesity and Is Responsive to an Essential Fatty Acid upon Murine Influenza Infection. J Immunol. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fan W, Chen XF, Shen C, Guo ZR, Dong C (2016) Hepatitis B vaccine response in obesity: A meta-analysis. Vaccine 34: 4835–4841. doi: 10.1016/j.vaccine.2016.08.027 [DOI] [PubMed] [Google Scholar]
- 38.Alberti L, Gilardini L, Girola A, Moro M, Cavagnini F, Invitti C (2007) Adiponectin receptors gene expression in lymphocytes of obese and anorexic patients. Diabetes Obes Metab 9: 344–349. doi: 10.1111/j.1463-1326.2006.00614.x [DOI] [PubMed] [Google Scholar]
- 39.Kollias A, Tsiotra PC, Ikonomidis I, Maratou E, Mitrou P, Kyriazi E, et al. (2011) Adiponectin levels and expression of adiponectin receptors in isolated monocytes from overweight patients with coronary artery disease. Cardiovasc Diabetol 10: 14 doi: 10.1186/1475-2840-10-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
First, forward scatter (FSC) and sideward scatter (SSC) profiles were used to roughly distinguish monocytes and lymphocytes/NK cells. Second, doublet cells were excluded using FSC-Area (FSC-A) and FSC-Height (FSC-H) gating. Finally, leukocyte subsets were gated on their marker expression.
(TIF)
Focussing on the CD25hiCD127lowCD4+ T-cells (regulatory T-cells), the adiponectin receptor 1-positive subset also expresses the leptin receptor.
(TIF)
(A) Circulating adiponectin levels versus percentage of adiponectin receptor 2 expressing iNKT cells (Pearson’s correlation coefficient 0.048, p = 0.764), and circulating leptin levels versus percentage of leptin receptor expressing regulatory T-cells (Pearson’s correlation coefficient -0.130, p = 0.416). (B) Circulating adiponectin levels versus median fluorescence intensity (MFI) of adiponectin receptor 1 and 2 expression (Pearson’s correlation coefficient AdipoR1 0.052, p = 0.746; AdipoR2–0.172, p = 0.284), and circulating leptin levels versus median fluorescence intensity (MFI) of leptin receptor expression (Pearson’s correlation coefficient 0.126, p = 0.431).
(TIF)
(DOC)
(DOC)
(DOC)
(SAV)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.