
Fig. 1.
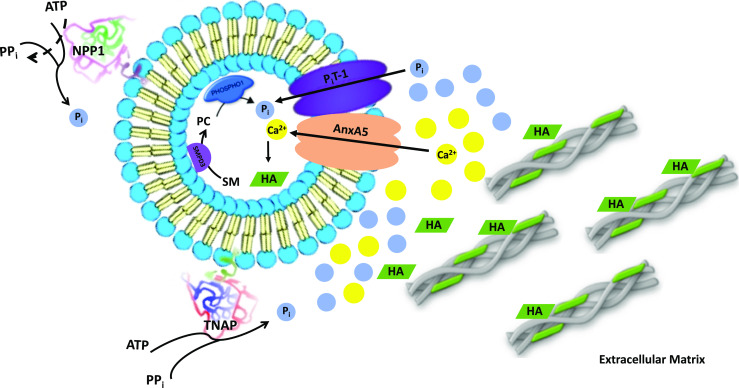
Proteoliposomes as matrix vesicles (MVs) biomimetic systems. The construction of MV models has the aim to provide an environment that would allow the initial nucleation of apatite inside of liposomes as well as to provide the surface nucleation that can also help in the propagation of apatite onto the extracellular matrix. The proteoliposomes internal nucleation can be initiated by orphan phosphatase 1 (PHOSPHO1) action producing inorganic phosphate (Pi) from the hydrolysis of phosphocoline (PC), which itself is derived from sphingomyelin (SM) by the action of sphingomyelin phosphodiesterase 3 (SMPD3) located in the inner surface of the MV membrane. In addition, protein transporters, such as annexin A5 (AnxA5) and phosphate transporter 1 (PiT-1), provide ions for nucleation inside the proteoliposomes. The events outside the vesicles are studied by the incorporation of the regulated proteins, tissue nonspecific alkaline phosphatase (TNAP) and ectonucleotide pyrophosphatase/phosphodiesterase (NPP1), which play a crucial role controlling appropriate inorganic pyrophosphate (PPi)/Pi ratio. Furthermore, calcium channels accelerate the nucleational activity of calcium–phosphate–lipid complexes (CPLX) inside the MVs. AnxA5 also shows binding affinity to type II collagen and when present in the proteoliposome membrane can drive the vesicles’ attachment onto to collagenous extracellular matrix stimulating mineral propagation. Hydroxyapatite (HA)