

Abstract
Purpose
To investigate the role of the group III/IV muscle afferents in the bioenergetics of exercising skeletal muscle beyond constraining the magnitude of metabolic perturbation.
Methods
Eight healthy men, performed intermittent isometric knee-extensor exercise to task failure at ~58% maximal voluntary contraction (MVC) under control conditions (CTRL) and with lumbar intrathecal fentanyl to attenuate group III/IV leg muscle afferents (FENT). Intramuscular concentrations of phosphocreatine (PCr), inorganic phosphate (Pi), diprotonated phosphate (H2PO4−), adenosine triphosphate (ATP), and pH were determined using phosphorous magnetic resonance spectroscopy (31P-MRS).
Results
The magnitude of metabolic perturbation was significantly greater in FENT compared to CTRL for [Pi] (37.8 ± 16.8 vs. 28.6 ± 8.6 mM), [H2PO4−] (24.3 ± 12.2 vs. 17.9 ± 7.1 mM), and [ATP] (75.8 ± 17.5 vs. 81.9 ± 15.8 % of baseline), while there was no significant difference in [PCr] (4.5 ± 2.4 vs. 4.4 ± 2.3 mM) or pH (6.51 ± 0.10 vs. 6.54 ± 0.14). The rate of perturbation in [PCr], [Pi], [H2PO4−], and pH was significantly faster in FENT compared to CTRL. Oxidative ATP synthesis was not significantly different between conditions. However, anaerobic ATP synthesis, through augmented creatine kinase and glycolysis reactions, was significantly greater in FENT than CTRL, resulting in a significantly greater ATP cost of contraction (0.049 ± 0.016 vs. 0.038 ± 0.010 mM·min−1·N−1).
Conclusion
Not only do group III/IV muscle afferents constrain magnitude of perturbation in intramuscular Pi, H2PO4−, and ATP during small muscle mass exercise, but also appear to play a role in maintaining efficient skeletal muscle contractile function in men.
Keywords: 31P-MRS, ATP cost of contraction, intrathecal fentanyl, skeletal muscle metabolism
Introduction
During exhaustive exercise, the intramuscular metabolic perturbation is constrained, such that task failure within a given exercise modality is associated with consistent intramuscular metabolite concentrations (e.g. PCr, Pi, H2PO4−, and pH) across varying conditions (11, 12, 22, 43, 44). Mechanistically, this constraint appears to be mediated by group III/IV muscle afferents, which innervate the extracellular space of skeletal muscle and respond in a dose-dependent manner to the mechanical and metabolic stimuli associated with muscle contractions to provide feedback to the brain and spinal cord (25). Specifically, [PCr], [Pi], adenosine diphosphate ([ADP]), [lactate], and pH were perturbed to a greater extent with the fentanyl-induced attenuation of feedback from these leg muscle afferents (7). However, the influence of group III/IV muscle afferents on exercising skeletal muscle bioenergetics (i.e. sources and rates of ATP synthesis) remains unknown.
Interestingly, although not highlighted in our group’s previous work (7), the attenuation of group III/IV muscle afferent feedback appeared to impair muscle efficiency during cycling exercise. This was evinced by a greater PCr depletion and hydrogen ion accumulation in the vastus lateralis, despite an unchanged mean power output, following the fentanyl-induced attenuation of feedback from leg muscle afferents (7). However, given the large variation in power output during these cycling time trials and utilization of highly localized, periodic biopsies, such an observation needs to be investigated using an experimental design better suited to assess such a phenomenon (i.e. constant force exercise) coupled with high-resolution measurements of skeletal muscle bioenergetics (31P-MRS).
Additionally, determining the direct influence of group III/IV muscle afferents on the skeletal muscle bioenergetics and exercise tolerance with pharmacological blockade has been complicated by experimental limitations. Oxygen delivery may be attenuated as a secondary consequence of the pharmacological blockade of group III/IV muscle afferents (3). This would exacerbate the rate of intramuscular metabolic perturbation and confound the direct assessment of group III/IV muscle afferent feedback on exercise tolerance, which may explain why exercise tolerance is unchanged or diminished despite the ability to attain a greater intramuscular metabolic perturbation (1, 2, 7). Such oxygen delivery limitations can likely be circumvented with specific exercise modalities (i.e. intermittent isometric exercise) due to the substantial hyperemia between contractions (34, 45, 46). Furthermore, the degree to which the group III/IV muscle afferents influence the physiological responses to exercise (i.e. circulatory and ventilatory) may differ with exercising muscle mass (1, 2, 36, 37). A small muscle mass exercise paradigm focused more on the muscle itself (i.e. isolated muscle group), would minimize the pulmonary or cardiovascular limitations that arise from the attenuation of group III/IV muscle afferents during large muscle mass exercise paradigms (i.e. whole-body). Importantly, modifying the experimental design based upon these observations will provide a greater understanding of the mechanistic role group III/IV muscle afferents play in the physiological response to exercise in humans.
Accordingly, the purpose of this investigation was to assess the influence of the group III/IV muscle afferents on the intramuscular metabolic perturbation and skeletal muscle bioenergetics during high-intensity intermittent isometric single-leg knee-extensor exercise, using lumbar intrathecal fentanyl and 31P-MRS. We hypothesized that, compared to control conditions, exercise with attenuated group III/IV muscle afferent feedback would 1) result in a greater magnitude of intramuscular metabolic perturbation, but 2) not alter the time-course of the bioenergetic response and, therefore, 3) prolong exercise tolerance.
Methods
Subjects
Eight healthy, recreationally active men (age: 28 ± 5 yrs, stature: 178 ± 4 cm, and body mass: 77 ± 8 kg) volunteered to participate in this study and provided written informed consent prior to testing. All experimental procedures were approved by the Institutional Review Boards of the University of Utah and the Salt Lake City Veteran’s Affairs Medical Center and were conducted in accordance with the Declaration of Helsinki. Subjects were instructed to abstain from vigorous activity for the 24 h preceding each visit to the laboratory and to arrive at the laboratory having abstained from food and caffeine for the preceding 3 h. Subjects visited the laboratory a minimum of four times with at least 72 h between each visit.
Experimental design and exercise modality
For all exercise protocols, subjects performed single-leg intermittent isometric knee-extensor exercise in a semi-recumbent position (~15° elevation of the trunk) with the knee situated over a custom-built knee support (~45° knee joint angle) and the ankle fixed to an immovable strain gauge (SSM-AJ-250, Interface). Non-elastic straps were positioned over the hips and thigh to minimize extraneous movements. For each protocol, intermittent isometric quadriceps exercise was performed with 3 s contraction and 2 s relaxation durations, which were cued by an audio recording. The instantaneous force and the target force were projected on a screen for the subjects to visualize and no information regarding the duration of exercise was provided to the subjects. The exercising leg was balanced with respect to leg dominance (i.e. MVC force) across subjects.
During the first preliminary visit, subjects performed 60 intermittent MVCs of the quadriceps over 5 min. For this protocol, peak MVC force was determined as the highest force attained during the 60 MVCs and the end-test force was determined as the mean force of the final 6 MVCs (10). These facilitated the determination of the target force to be used for the experimental testing, which was calculated as the end-test force plus 20% of the difference between the end-test force and the peak MVC force (11), chosen to elicit task failure in ~5 min. During the second preliminary visit, subjects were familiarized with the time to task failure exercise protocol that would be employed in the subsequent experimental visits. For this protocol, subjects maintained the target force for each 3 s contraction and continued until task failure, which was defined as the inability to attain the target force during three consecutive contractions.
For the two experimental visits, subjects performed the same time to task failure exercise protocol inside a whole body magnetic resonance imaging (MRI) system under both CTRL and FENT conditions. Exercise in these experimental visits was followed a 5 min rest period in order to measure intramuscular metabolic recovery kinetics. The experimental conditions (CTRL or FENT) were conducted in a balanced order across subjects. Prior to positioning the subject inside the MRI, two 5 s MVCs were performed to determine peak force production. Immediately at task failure, a rating of perceived exertion (RPE) was obtained from the subjects using the Borg modified CR10 (8).
Intrathecal fentanyl administration
Subjects were seated in an upright position with a slight flexion of the torso. The skin and subcutaneous tissue at the L3-L4 vertebral interspace were anesthetized using 1% lidocaine (10 mg·ml−1). A 20-gauge spinal introducer needle was then inserted at the L3-L4 vertebral interspace through which a 25-gauge Whitacre needle was advanced into the intrathecal space, which was confirmed by free-flowing cerebrospinal fluid. A small amount of cerebrospinal fluid was aspirated and 0.025 mg of fentanyl (0.050 mg·ml−1) was injected into the intrathecal space in order to attenuate feedback arising from μ-opioid receptor-sensitive group III/IV leg muscle afferents without affecting motor nerve activity or maximal force output (2, 16, 40, 47). Subjects remained in an upright seated position for at least 5 min prior to being positioned for the exercise protocol. Previous studies have documented that this intrathecal fentanyl administration protocol does not result in fentanyl migration to the brain (1, 2, 7, 20, 38, 39).
31P-MRS
MRS was performed using a clinical 2.9 Tesla MRI system (Tim-Trio, Siemens Medical Systems) operating at 49.9 MHz for 31P resonance. A dual 31P-1H surface coil (110 mm 1H coil loop surrounded by the 31P single-loop coil with a diameter of 125 mm) with linear polarization was used to acquire 31P-MRS data (Rapid Biomedical GmbH). In order to sample all major muscles of the quadriceps, the surface coil was secured at mid-thigh level with elastic straps and advanced localized volume shimming was performed after a three-plane scout proton image was acquired. Prior to each experiment, two fully relaxed spectra were acquired at rest with three averages per spectrum and a repetition time of 30 s. Throughout the exercise and recovery portions of the protocol, MRS data acquisition was performed using a free-induction decay pulse sequence with a 2.56 ms adiabatic-half-passage excitation RF pulse, a repetition time of 2.5 s, a receiver bandwidth of 5 kHz, 1024 data points, and 2 averages per spectrum. Saturation factors were quantified by the comparison between fully relaxed (repetition time = 30 s) and partially relaxed (repetition time = 2.5 s) spectra.
Absolute and relative concentrations of intramuscular PCr, Pi, and ATP were determined by a time-domain fitting routine using the AMARES algorithm (42) incorporated into the CSAIPO software (31, 33), assuming a resting [ATP] of 8.2 mM (17). Intracellular pH was calculated from the chemical shift difference between the Pi and PCr signals. The free cytosolic [ADP] was calculated from [PCr] and pH using the creatine kinase (CK) equilibrium constant ( ), with the assumption that PCr represents 85% of the total creatine content (23). Resting concentrations were calculated from the average peak areas of the two fully relaxed spectra. When Pi splitting was evident, the pH corresponding to each Pi pool was calculated separately as pH1 and pH2 on the basis of the chemical shift of each peak relative to PCr, such that the overall pH was then calculated as:
The concentration of the H2PO4− was calculated as (30):
Free cytosolic adenosine monophosphate (AMP) was calculated based on the equilibrium of the adenylate kinase reaction corrected for the effects of pH, assuming a free magnesium concentration of 1 mM (15). In all cases, relative amplitudes were corrected for partial saturation due to the repetition time relative to T1 via the fully relaxed spectra acquired at rest.
ATP synthesis rates and ATP cost of contraction
The rate of ATP production from the breakdown of PCr through the CK reaction (ATPCK, mM·min−1) was calculated from the change in [PCr] for each time point of the exercise period (26):
Based on the sigmoid relationship between the oxidative ATP production rate (ATPOX, mM·min−1) and free cytosolic [ADP], the rate of mitochondrial ATP production was calculated as:
in which Km (the [ADP] at half-maximal oxidation rate) is ~30 μM in skeletal muscle (26), 2.2 is the Hill coefficient for a sigmoid function (24), and Vmax is the peak rate of in vivo oxidative ATP synthesis.
During exercise, changes in intramuscular pH result from glycogen breakdown to pyruvate and lactate, proton efflux, buffering capacity, protons produced by oxidative phosphorylation, and the consumption of protons by the CK reaction (26). Assuming that the glycogenolytic production of 1 mole of H+, when coupled to ATP hydrolysis, yields 1.5 mole of ATP, the ATP production from anaerobic glycolysis (ATPGLY) can be deduced from the total number of protons ( ) produced throughout exercise (21, 26, 27):
where
(in mM·min−1) was calculated from the time-dependent changes in [PCr] and from the stoichiometric coefficient (γ):
where γ is the proton stoichiometric coefficient of the coupled Lohmann reaction as previously described (29). (in mM·min−1) was calculated from the apparent buffering capacity βtotal (in Slykes, millimoles acid added/unit change in pH) and from the rate of pH changes:
where
where
in which βa was determined from the initial change in PCr (ΔPCri) and alkalinization of pH (ΔpH) (13):
βPi was determined based on the dissociation constant of the buffer (K) according to the standard formula (14):
where K = 1.77 × 10−7. In agreement with previous studies and assuming that muscle is a closed system during exercise (14, 27), βbicarbonate was set to zero. (in mM·min−1) was calculated from the factor m = 0.16/[1 + 10(6.1 − pH)], which accounts for the amount of protons produced through oxidative ATP production (27, 28):
(in mM·min−1) was calculated for each time point of exercise using the proportionality constant λ relating proton efflux rate to ΔpH (27, 28):
This proportionality constant λ (in mM·min−1·pH unit−1) was calculated during the recovery period:
During the recovery period, PCr is regenerated throughout the CK reaction as the consequence of oxidative ATP production in mitochondria. Thus, can be calculated from the rates of proton production from the CK reaction ( , in mM·min−1) and mitochondrial ATP production ( , in mM·min−1) on one side and the rate of pH changes on the other side. At this time, ATP production is exclusively aerobic and lactate production is considered negligible:
To improve precision, a modified version of this calculation was used (27, 28), in which the total proton disappearance (i.e. ∫Edt) is estimated cumulatively from the start of recovery and then fitted to an exponential function to obtain the initial recovery rate (Vefflux).
The total ATPase rate (ATPTOTAL, in mM·min−1) was calculated for each time point as:
and the anaerobic ATPase rate (ATPANA, mM·min−1) was calculated for each time point as:
The ATP cost of contraction (in mM·N−1) was calculated as the ratio between ATPTOTAL and the force integral. All values were calculated for the first and final minutes of exercise.
PCr recovery kinetics
The [PCr] kinetics during the recovery period were described by a mono-exponential curve:
where [PCr](t) is the [PCr] at a given time (t), [PCr]end is the [PCr] at task failure, Δ[PCr] is the amount of PCr resynthesized during the recovery period, and τ represents the time constant of the PCr offset kinetics. The initial rate of PCr resynthesis (ViPCr) was calculated from the derivative of the mono-exponential equation at the onset of recovery:
where Δ[PCr] is the amount of PCr resynthesized during the recovery period and the rate constant (26). Vmax was calculated using ViPCr and the [ADP] at task failure (41):
where Km (the [ADP] at half the highest oxidative rate) is ~30 μM in skeletal muscle (26).
Model variables were determined with an iterative process by minimizing the sum of squared residuals (RSS) between the fitted function and the observed values. Goodness of fit was assessed by visual inspection of the residual plot and the frequency plot distribution of the residuals, with the Chi square values and the coefficient of determination (r2) calculated as follows:
Statistical analysis
The integral force per contraction and 31P-MRS data were analyzed using two-way ANOVAs (condition × time) with repeated measures. Tukey’s post hoc analyses were conducted when a significant main effect was detected. The MVC force, mean force of the exercise, cumulative force integral, time to task failure, peak rate of mitochondrial ATP synthesis, and RPE data were compared between conditions using Student’s paired t tests. Pearson product moment correlation analyses were utilized to assess relationships between the percent change between FENT and CTRL in intramuscular metabolite concentration at task failure. Significance for the statistical analysis was accepted at p ≤ 0.05. Results are presented as means ± SD, except for in Figures where SEM is used for clarity.
Results
Fentanyl blockade, maximal force production, exercise tolerance, and perception of effort
Fentanyl blockade did not significantly alter quadriceps MVC force prior to the start of exercise (FENT: 591.6 ± 153.6 vs. CTRL: 574.4 ± 113.3 N, p = 0.41). There was no significant difference in the force integral per contraction over time or between conditions (p > 0.68). For the entire exercise bout, there were no significant differences between conditions for the cumulative force integral (FENT: 45381 ± 8472 vs. CTRL: 47982 ± 7832 N, p = 0.25), mean force (FENT: 329 ± 23 vs. CTRL: 331 ± 81 N, p = 0.80), %MVC force (FENT: 57 ± 9 vs. CTRL: 58 ± 11 %MVC, p = 0.77), or time to task failure (FENT: 239 ± 73 vs. CTRL: 257 ± 79 s, p = 0.12) (Figure 1). The RPE, assessed at task failure, was significantly lower (~50 %) with fentanyl blockade (FENT: 4.5 ± 2.6 vs. CTRL: 9.7 ± 0.5).
Figure 1. Force development and exercise tolerance during high-intensity intermittent isometric single-leg knee-extensor exercise.
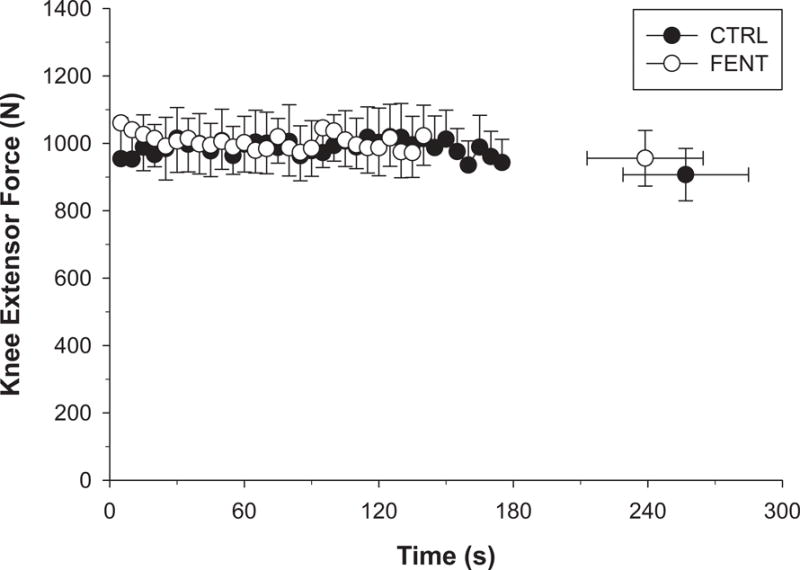
Subjects were required to attain a target force during each 3 s contraction and the force integral of each contraction was calculated during control (CTRL) and fentanyl (FENT) conditions. Values are mean ± SEM for each contraction up to the last common time point prior to task failure for all participants and at task failure. The time to task failure, force integral per contraction, and the cumulative force integral were not significantly different between experimental conditions.
Fentanyl blockade, intramuscular metabolites, and intracellular pH
Fentanyl blockade did not significantly alter baseline [PCr] (FENT: 26.8 ± 3.0 vs. CTRL: 25.3 ± 1.5 mM, p = 0.22), [Pi] (FENT: 1.3 ± 0.4 vs. CTRL: 0.9 ± 0.4 mM, p = 0.75), H2PO4‒ (FENT: 0.5 ± 0.1 vs. CTRL: 0.3 ± 0.1 mM, p = 0.81), or pH (FENT: 7.00 ± 0.02 vs. CTRL: 7.01 ± 0.03, p = 0.64). The intramuscular metabolites and pH data during exercise are illustrated in Figure 2. Specifically, there was a significantly greater magnitude of perturbation in [Pi], [H2PO4−], and [ATP] for FENT than CTRL, while the degree of perturbation in [PCr] (p = 0.95) and pH (p = 0.24) were not significantly altered by fentanyl blockade (Table 1). The greater depletion of [ATP] in FENT compared to CTRL was significantly correlated with a greater accumulation of [Pi] (r = −0.70) and [H2PO4−] (r = −0.79), while there was no significant correlation with [PCr] (r = −0.18, p = 0.67). The rate of perturbation in [PCr], [Pi], [H2PO4−], and pH was significantly faster for FENT than CTRL (Figure 2).
Figure 2. Intramuscular metabolic perturbation during high-intensity intermittent isometric single-leg knee-extensor exercise.
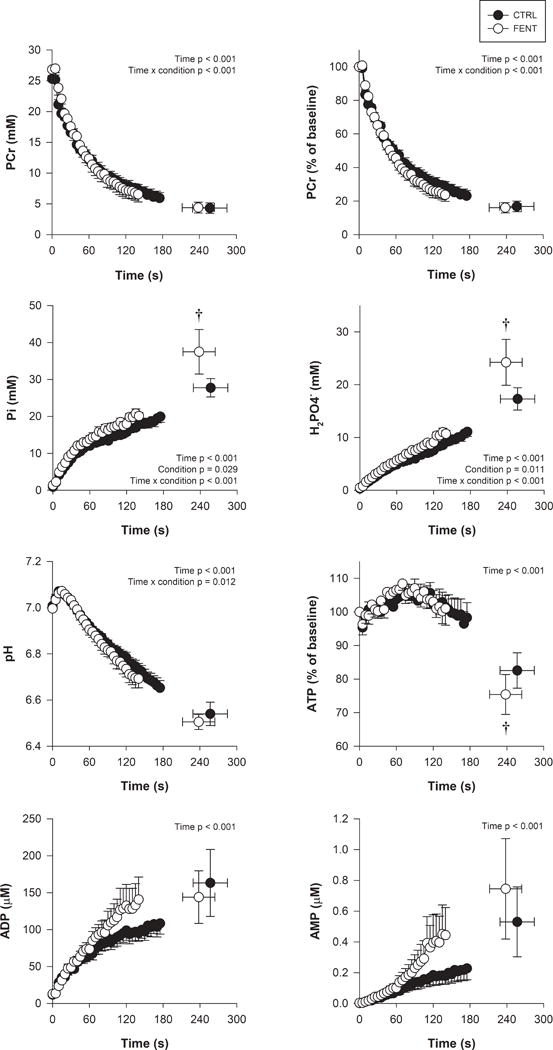
Intramuscular metabolite concentrations were obtained using 31P-MRS during control (CTRL) and fentanyl (FENT) conditions. Main effects and interactions are indicated in the graphs. Values are mean ± SEM for each contraction up to the last common time point prior to task failure for all participants and at task failure. † significantly different from CTRL at specific time point.
Table 1.
Intramuscular metabolite concentrations at task failure for high-intensity intermittent isometric single-leg knee-extensor exercise
| CTRL
|
FENT
|
|
|---|---|---|
| [PCr] (mM) | 4.3 ± 2.4 | 4.4 ± 2.5 |
| [PCr] (%) | 16.8 ± 8.9 | 16.1 ± 8.7 |
| [Pi] (mM) | 27.7 ± 7.0 | 37.5 ± 17.1† |
| [H2PO4−] (mM) | 17.3 ± 6.0 | 24.2 ± 12.3† |
| pH | 6.54 ± 0.14 | 6.51 ± 0.09 |
| [ATP] (%) | 82.6 ± 14.9 | 75.4 ± 16.8† |
Values are mean ± SD. Intramuscular metabolite concentrations were determined using phosphorous magnetic resonance spectroscopy in control (CTRL) and fentanyl (FENT) conditions. PCr, phosphocreatine; Pi, inorganic phosphate; H2PO4−, diprotonated Pi; ATP, adenosine triphosphate. PCr and ATP are presented as a percentage of baseline values.
significantly different from CTRL.
ATP synthesis rates, the ATP cost of contraction, and PCr recovery kinetics
The ATP synthesis rates and the ATP cost of contraction data are illustrated in Figure 3. ATPTOTAL was significantly greater for FENT than CTRL. ATPOX was not significantly different between conditions (p = 0.14), but ATPANA was significantly greater in FENT than in CTRL. This was the result of a greater ANACK and ANAGLY for FENT than CTRL. The percentage contribution of ATPOX (FENT: 56 ± 11 vs. CTRL: 52 ± 7 %, p = 0.44) and ATPANA (FENT: 44 ± 11 vs. CTRL: 48 ± 7 %, p = 0.42) to ATPTOTAL over the 1st min of exercise was not significantly different between conditions. In contrast, over the final min of exercise, there was a significantly greater percentage contribution of ATPANA (FENT: 35 ± 12 vs. CTRL: 27 ± 11 %), and therefore a significantly lower contribution of ATPOX (FENT: 65 ± 12 vs. CTRL: 73 ± 11 %), to ATPTOTAL for FENT compared to CTRL (Figure 3). The greater ATPTOTAL for FENT than CTRL, with no significant differences in force production, resulted in a greater ATP cost of contraction in FENT compared to CTRL. The variables describing PCr recovery kinetics were not significantly different between conditions (Table 2). Over the final min of exercise, ATPOX was not significantly different from Vmax.
Figure 3. Adenosine triphosphate (ATP) synthesis rates and ATP cost of contraction during high-intensity intermittent isometric single-leg knee-extensor exercise.
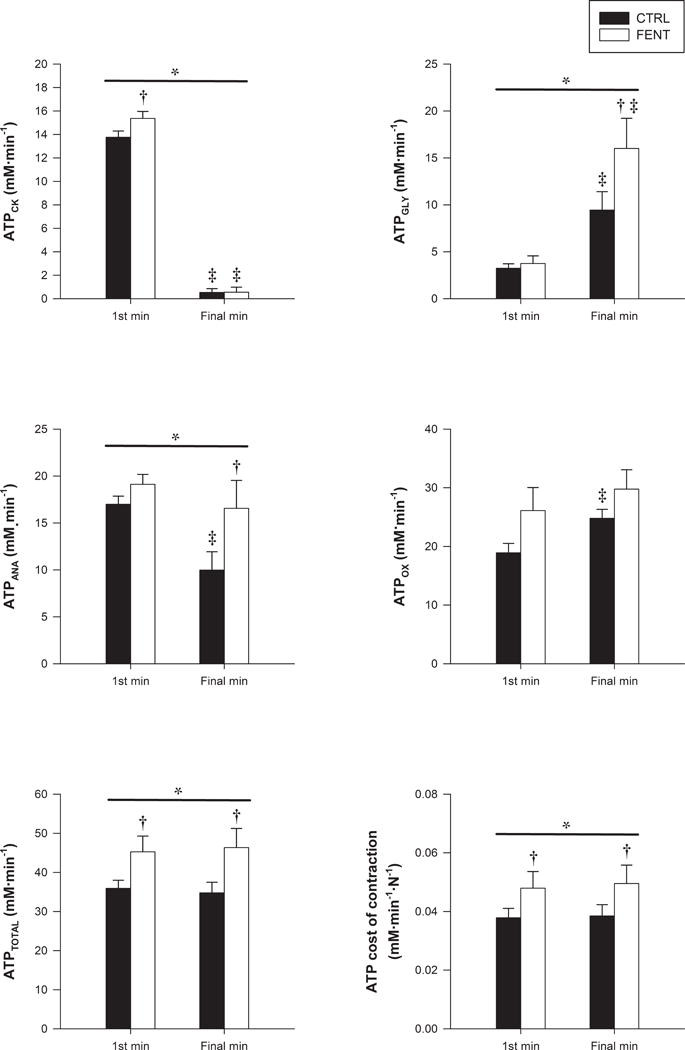
The rate of ATP synthesis through the creatine kinase reaction (ATPCK), anaerobic glycolysis (ATPGLY), cumulative anaerobic metabolism (ATPANA), oxidative phosphorylation (ATPOX), and total ATPase rate (ATPTOTAL) and the ATP cost of contraction for the 1st and final minutes of exercise between fentanyl (FENT) and control (CTRL) conditions. Values are mean ± SEM. * significant main effect for condition, † significantly different from CTRL, and ‡ significantly different from 1st min.
Table 2.
Parameters of PCr dynamics during recovery from high-intensity intermittent isometric single-leg knee-extensor exercise
| CTRL
|
FENT
|
|
|---|---|---|
| Δ[PCr] (mM) | 23.3 ± 2.6 | 26.0 ± 4.3 |
| Tau (s) | 58.4 ± 11.2 | 62.2 ± 7.7 |
| (mM·min−1) | 26.6 ± 4.7 | 28.1 ± 3.1 |
| (mM·min−1) | 24.4 ± 4.1 | 25.3 ± 4.5 |
| (mM·min−1) | 5.1 ± 3.3 | 8.1 ± 3.5 |
Values are mean ± SD. Intramuscular phosphocreatine (PCr) dynamics during 5 min of recovery were fit with a monoexponential model in control (CTRL) and fentanyl (FENT) conditions. Δ[PCr], amount of PCr resynthesized during the recovery period; Tau, time constant of PCr resynthesis; Vmax, peak rate of mitochondrial respiration; ViPCr, initial rate of PCr resynthesis; H+ efflux, proton efflux from muscle.
Discussion
This investigation utilized lumbar intrathecal fentanyl administration and 31P-MRS to elucidate the influence of the lower limb group III/IV muscle afferents on the intramuscular metabolic perturbation and skeletal muscle bioenergetics during high-intensity intermittent isometric single-leg knee-extensor exercise. In agreement with our first hypothesis, a greater magnitude of perturbation in intramuscular Pi, H2PO4−, and ATP was incurred with attenuated group III/IV muscle afferent feedback. However, the attenuation of group III/IV muscle afferent feedback resulted in a faster rate of perturbation in intramuscular PCr, Pi, H2PO4−, pH, and ATP and, therefore, did not prolong exercise tolerance, which is in disagreement with our second and third hypotheses. Importantly, the faster rates of intramuscular metabolic perturbation appear not to have been a consequence of limited oxygen delivery, as oxidative ATP synthesis, , and the peak rate of mitochondrial ATP synthesis were not affected by fentanyl, but rather a greater ATP cost of contraction. Collectively, these findings further support a critical role of the group III/IV muscle afferents in constraining the magnitude of intramuscular metabolic perturbation, even during small muscle mass exercise, but, additionally, unveil that these muscle afferents also appear to play a critical role in maintaining efficient skeletal muscle contractile function during exercise in men.
Group III/IV muscle afferents and the degree of intramuscular metabolic perturbation during exercise
It is now apparent that specific concentrations of intramuscular metabolites are attained at task failure during both large and small muscle mass exercise, which are robust across varying conditions within a given exercise modality (11, 12, 22, 43, 44). It was recently demonstrated, that the physiological mechanism responsible for this constraint on the magnitude of intramuscular metabolic perturbation during large muscle mass exercise is, at least in part, mediated by the group III/IV muscle afferents (7). Consistent with this finding, a greater magnitude of perturbation in intramuscular Pi, H2PO4−, and ATP was incurred during small muscle mass exercise with the attenuation of group III/IV muscle afferent feedback in the current study. Collectively, the observations from both large and small muscle mass exercise document a critical role of the group III/IV muscle afferents in constraining the magnitude of intramuscular metabolic perturbation attained at exercise task failure.
The generally similar influence of group III/IV muscle afferents on the magnitude of intramuscular metabolic perturbation during large and small muscle mass exercise are, however, not without some noteworthy metabolic differences that may indicate disparate determinants of exercise tolerance. Specifically, in CTRL conditions, the intramuscular [PCr] depletion in the current investigation (~80 %, Figure 2) was greater than that previously reported for large muscle mass exercise (~60 %), which also resulted in the attainment of a lower pH (~6.5) compared to large muscle mass exercise (~6.7) (7, 43). Of note, these CTRL [PCr] and pH values at task failure are similar to previous reports using small muscle mass exercise (9, 11). These findings are consistent with the typically greater level of peripheral fatigue developed, and therefore intramuscular metabolic perturbation (7), during small muscle mass exercise compared to large muscle mass exercise (36, 37). Furthermore, the attenuation of group III/IV muscle afferent feedback did not alter the end-exercise intramuscular PCr depletion or pH in the current study (Figure 2), while this exacerbated the PCr depletion (~27 %) and pH during large muscle mass exercise (7). These findings may be indicative of a nadir degree of [PCr] depletion (~80 %) within the quadriceps muscle group, that is only attained during large muscle mass exercise when feedback from the group III/IV muscle afferents is attenuated, but this is actually attained during CTRL when exercise is confined to a small muscle mass where such a greater degree of intramuscular metabolic perturbation is tolerated (36, 37). Contrary to the observations on PCr and pH, the attenuation of the group III/IV muscle afferent feedback resulted in a greater ATP depletion in the current investigation (~7 %, Figure 2), but did not alter intramuscular [ATP] depletion during large muscle mass exercise (7). Interestingly, this greater [ATP] depletion likely acounted for the greater accumulation of [Pi] and [H2PO4−] with the attenuation of group III/IV muscle afferent feedback, as supported by the correlation between these data and the lack of an effect of this attenuation on PCr depletion. Thus, while the general influence of the group III/IV muscle afferents on the magnitude of intramuscular metabolic perturbation during large and small muscle mass exercise are similar, these findings reveal important differences that further highlight the important impact of active muscle mass and afferent activity.
Group III/IV muscle afferents, ATP synthesis, the ATP cost of contraction, and mitochondrial function
The 31P-MRS technique used in this study allowed the time-course of the exercise-induced intramuscular metabolic perturbation to be assessed in a large portion of the active muscle. Interestingly, despite similar force production, a faster rate of change in intramuscular [PCr], [Pi], [H2PO4−], and pH was evident with the attenuation of group III/IV muscle afferent feedback. It could be postulated that this faster rate of change may have been the result of an attenuated oxygen delivery compared to CTRL, which has been demonstrated for dynamic knee-extensor exercise (3). However, this likely was not the case as the oxidative ATP synthesis, the , and the peak rate of mitochondrial ATP synthesis, all of which are sensitive to alterations in oxygen delivery (18, 19, 27, 28, 32), were similar between conditions. Rather than an oxygen delivery limitation, the faster rate of exercise-induced metabolic perturbation appears to have been the result of an increased ATP cost of contraction when group III/IV muscle afferent feedback was attenuated (Figure 3). Specifically, this attenuation of group III/IV muscle afferent feedback did not alter force production, but did increase ATPTOTAL due to an augmented ATPANA (creatine kinase and glycolysis reactions) without an increase in ATPOX, likely due to the attainment of the peak rate of oxidative ATP synthesis in vivo (i.e. ATPOX = Vmax). Additionally, it is interesting to note that the similar PCr recovery kinetics between conditions suggests that the greater magnitude and rate of metabolic perturbation, achieved under conditions of limited muscle afferent feedback, had no impact on skeletal muscle respiratory function. In light of the potentially protective role of skeletal muscle afferent feedback, this interesting observation deserves a more comprehensive investigation.
The definitive identification of the mechanisms responsible for the increased ATP cost of contraction with attenuation of the group III/IV muscle afferent feedback are beyond the scope of the current investigation. However, it is likely that muscle afferent feedback is important for efficient force production during exercise. For example, the motor unit rotation that has been demonstrated to occur during exercise (5) may rely on feedback from the group III/IV muscle afferents. Additionally, the group III/IV muscle afferents appear to be involved in the alteration of motor unit firing rate during exercise to align with changes in the tetanic fusion frequency (6). Without feedback from group III/IV muscle afferents, the efficacious recruitment of motor units and their firing rate may be limited throughout exercise, resulting in compromised muscle function and an increased ATP cost of contraction. With such a hypothesis, it is important to note that the potential influence of the group III/IV muscle afferents on skeletal muscle metabolism is through an indirect mechanism rather than a direct role of these afferents on the intracellular metabolic processes. The increased ATP cost of contraction in FENT compared to CTRL may also contribute to the discrepant findings related to exercise tolerance across previous studies, in addition to the presumed alterations in oxygen delivery (1–3, 7). Thus, by both interrogating the bioenergetic time course during exercise and potentially limiting the impact of group III/IV muscle afferents on oxygen delivery, this study has unveiled a potential role for this afferent feedback in the economy of skeletal muscle contraction during exercise.
Group III/IV muscle afferents and the perception of effort during exercise
Although controversial, it has been postulated that RPE during exercise is exclusively determined by the corollary discharge associated with central motor drive and is not influenced by sensory feedback from the periphery, including skeletal muscle and organs (35). Contrary to this theory, there is experimental evidence that muscle afferent feedback modulates RPE during exercise (4). Remarkably, in the current investigation, RPE at task failure was ~50 % lower in FENT than CTRL, despite similar exercise durations and cumulative force integrals across conditions. Based on corollary discharge as the sole determinant of RPE, this reduction in RPE would necessitate a reduction in central motor drive. However, as a decrease in central motor drive during fatiguing exercise at a constant force is highly unlikely, the observed reduction in RPE in the face of a similar, or potentially higher (1), neural drive during FENT compared to CTRL suggests a dissociation between RPE and central motor drive. These findings provide additional evidence supporting the concept that group III/IV muscle afferents modulate RPE during exercise.
Conclusion
The findings of the current investigation further document the important role of the group III/IV muscle afferents in the exercising human. Specifically, consistent with evidence from large muscle mass exercise, the muscle afferents appear to constrain the magnitude of perturbation in intramuscular Pi, H2PO4−, and ATP during small muscle mass exercise. Additionally, the muscle afferents also increase the rate of intramuscular metabolic perturbation during small muscle mass exercise, which appears to be mediated by an increase ATP cost of muscle contraction. Collectively, these findings document an important role for the group III/IV muscle afferents in the bioenergetics and economy of exercising skeletal muscle in men.
Acknowledgments
The results of the present study are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation. The results of the present study do not constitute endorsement by ACSM.
Funding
This study was supported by NIH Heart, Lung, and Blood Institute grants (HL-103786, HL-116579, HL-091830, and K99HL125756), Veterans Affairs Rehabilitation Research and Development Merits (E6910-R and E1697-R), Spires (E1572-P and E1433-P), Senior Research Career Scientist award (E9275-L), and the Flight Attendant Medical Research Institute (YFEL141011).
Footnotes
Disclosures
The authors declare no conflict of interest, financial or otherwise.
The results of the present study are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation. The results of the present study do not constitute endorsement by ACSM.
Author contributions
The work was conducted at the Utah Vascular Research Laboratory (UVRL) housed in the Salt Lake City Veterans Affairs Hospital, Utah. All authors contributed to data collection/analysis and manuscript preparation. All authors approved the final version of the manuscript. All persons designated as authors qualify for authorship, and all those who qualify for authorship are listed.
References
- 1.Amann M, Blain G, Proctor LT, Sebranek JJ, Pegelow DF, Dempsey JA. Implications of group III and IV muscle afferents for high-intensity endurance exercise performance in humans. J Physiol. 2011;589(21):5299–309. doi: 10.1113/jphysiol.2011.213769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Amann M, Proctor LT, Sebranek JJ, Pegelow DF, Dempsey JA. Opiod-mediated muscle afferents inhibit central motor drive and limit peripheral muscle fatigue development in humans. J Physiol. 2009;587(1):271–83. doi: 10.1113/jphysiol.2008.163303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Amann M, Runnels S, Morgan DE, et al. On the contribution of group III and IV muscle afferents to the circulatory response to rhythmic exercise in humans. J Physiol. 2011;589(15):3855–66. doi: 10.1113/jphysiol.2011.209353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Amann M, Venturelli M, Ives SJ, et al. Peripheral fatigue limits endurance exercise via a sensory feedback-mediated reduction in spinal motoneuronal output. J Appl Physiol. 2013;115:355–64. doi: 10.1152/japplphysiol.00049.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bawa P, Murnaghan C. Motor unit rotation in a variety of human muscles. J Neurophysiol. 2009;102:2265–72. doi: 10.1152/jn.00278.2009. [DOI] [PubMed] [Google Scholar]
- 6.Bigland-Ritchie BR, Dawson NJ, Johansson RS, Lippold OCJ. Reflex origin for the slowing of motoneurone firing rates in fatigue of human voluntary contractions. J Physiol. 1986;379:451–9. doi: 10.1113/jphysiol.1986.sp016263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Blain GM, Mangum TS, Sidhu SK, et al. Group III/IV muscle afferents limit the intramuscular metabolic perturbation during whole body exercise in humans. J Physiol. 2016;594(18):5303–15. doi: 10.1113/JP272283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Borg G. Borg’s perceived exertion and pain scales. Champaign, IL: Human Kinetics; 1998. [Google Scholar]
- 9.Broxterman RM, Layec G, Hureau TJ, Amann M, Richardson RS. Skeletal muscle bioenergetics during all-out exercise: mechanistic insight into the oxygen uptake slow component and neuromuscular fatigue. J Appl Physiol. 2017 doi: 10.1152/japplphysiol.01093.2016. jap 01093 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Burnley M. Estimation of critical torque using intermittent isometric maximal voluntary contractions of the quadriceps in humans. J Appl Physiol. 2009;106:975–83. doi: 10.1152/japplphysiol.91474.2008. [DOI] [PubMed] [Google Scholar]
- 11.Burnley M, Vanhatalo A, Fulford J, Jones AM. Similar metabolic perturbations during all-out and constant force exhaustive exercise in humans: a 31P magnetic resonance spectroscopy study. Exp Physiol. 2010;95(7):798–807. doi: 10.1113/expphysiol.2010.052688. [DOI] [PubMed] [Google Scholar]
- 12.Chidnok W, Fulford J, Bailey SJ, et al. Muscle metabolic determinants of exercise tolerance following exhaustion: relationship to the “critical power”. J Appl Physiol. 2013;115:243–50. doi: 10.1152/japplphysiol.00334.2013. [DOI] [PubMed] [Google Scholar]
- 13.Conley KE, Blei ML, Richards TL, Kushmerick MJ, Jubrias SA. Activation of glycolysis in human muscle in vivo. American Journal of Physiology Cell Physiology. 1997;42:C306–C15. doi: 10.1152/ajpcell.1997.273.1.C306. [DOI] [PubMed] [Google Scholar]
- 14.Conley KE, Kushmerick MJ, Jubrias SA. Glycolysis is independent of oxygenation state in stimulated human skeletal muscle in vivo. J Physiol. 1998;511(3):935–45. doi: 10.1111/j.1469-7793.1998.935bg.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Golding EM, Teague WE, Dobson GP. Adjustment of Kʹ to varying pH and pMg for the creatine kinase, adenylate kinase and ATP hydrolysis equilibria permitting quantitative bioenergetic assessment. The Journal of Experimental Biology. 1995;198:1775–82. doi: 10.1242/jeb.198.8.1775. [DOI] [PubMed] [Google Scholar]
- 16.Grant GJ, Susser L, Cascio M, Moses M, Zakowski MI. Hemodynamic effects of intrathecal fentanyl in nonlaboring term parturients. J Clin Anesth. 1996;8(2):99–103. doi: 10.1016/0952-8180(95)00174-3. [DOI] [PubMed] [Google Scholar]
- 17.Harris RC, Hultman E, Nordesjo LO. Glycogen, glycolytic intermediates and high-energy phosphates determined in biopsy samples of musculus quadriceps femoris of man at rest. Methods and variance of values. Scand J Clin Lab Invest. 1974;33(2):109–20. [PubMed] [Google Scholar]
- 18.Haseler LJ, Hogan MC, Richardson RS. Skeletal muscle phosphocreative recovery in exercise-trained humans is dependent on O2 availability. J Appl Physiol. 1999;86:2013–8. doi: 10.1152/jappl.1999.86.6.2013. [DOI] [PubMed] [Google Scholar]
- 19.Haseler LJ, Lin A, Hoff J, Richardson RS. Oxygen availability and PCr recovery rate in untrained human calf muscle: evidence of metabolic limitation in normoxia. American Journal of Physiology Regulatory, Integrative and Comparative Physiology. 2007;293:R2046–R51. doi: 10.1152/ajpregu.00039.2007. [DOI] [PubMed] [Google Scholar]
- 20.Hilty L, Lutz K, Maurer K, et al. Spinal opiod receptor-sensitive muscle afferents contribute to the fatigue-induced increase in intracortical inhibition in healthy humans. Exp Physiol. 2011;96(5):505–17. doi: 10.1113/expphysiol.2010.056226. [DOI] [PubMed] [Google Scholar]
- 21.Hochachka PW, Mommsen TP. Protons and anaerobiosis. Science. 1983;219:1391–7. doi: 10.1126/science.6298937. [DOI] [PubMed] [Google Scholar]
- 22.Hogan MC, Richardson RS, Haseler LJ. Human muscle performance and PCr hydrolysis with varied inspired oxygen fractions: a 31P-MRS study. J Appl Physiol. 1999;86(4):1367–73. doi: 10.1152/jappl.1999.86.4.1367. [DOI] [PubMed] [Google Scholar]
- 23.Jeneson JA, Westerhoff HV, Brown TR, Van Echteld CJA, Berger R. Quasi-linear relationship between Gibbs free energy of ATP hydrolysis and power output in human forearm muscle. American journal of Physiology Cell Physiology. 1995;37:C1474–C84. doi: 10.1152/ajpcell.1995.268.6.C1474. [DOI] [PubMed] [Google Scholar]
- 24.Jeneson JA, Wiseman RW, Westerhoff HV, Kushmerick MJ. The signal transduction function for oxidative phosphorylation is at least second order in ADP. The Journal of Biological Chemistry. 1996;271(45):27995–8. doi: 10.1074/jbc.271.45.27995. [DOI] [PubMed] [Google Scholar]
- 25.Kaufman MP, Hayes SG, Adreani CM, Pickar JG. Discharge properties of group III and IV muscle afferents. Adv Exp Med Biol. 2002;508:25–32. doi: 10.1007/978-1-4615-0713-0_4. [DOI] [PubMed] [Google Scholar]
- 26.Kemp GJ, Radda GK. Quantitative interpretation of bioenergetic data from 31P and 1H magnetic resonance spectroscopic studies of skeletal muscle: an analytical review. Magn Reson Q. 1994;10(1):43–63. [PubMed] [Google Scholar]
- 27.Kemp GJ, Taylor DJ, Styles P, Radda GK. The production, buffering and efflux of protons in human skeletal muscle during exercise and recovery. NMR Biomed. 1993;6:73–83. doi: 10.1002/nbm.1940060112. [DOI] [PubMed] [Google Scholar]
- 28.Kemp GJ, Thompson CH, Taylor DJ, Radda GK. Proton efflux in human skeletal muscle during recovery from exercise. Eur J Appl Physiol Occup Physiol. 1997;76:462–71. doi: 10.1007/s004210050276. [DOI] [PubMed] [Google Scholar]
- 29.Kushmerick MJ. Multiple equilibria of cations with metabolites in muscle bioenergetics. American Journal of Physiology Cell Physiology. 1997;41:C1739–C47. doi: 10.1152/ajpcell.1997.272.5.C1739. [DOI] [PubMed] [Google Scholar]
- 30.Lanza IR, Wigmore DM, Befroy DE, Kent-Braun JA. In vivo ATP production during free-flow and ischaemic muscle contractions in humans. J Physiol. 2006;577(1):353–67. doi: 10.1113/jphysiol.2006.114249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Layec G, Bringard A, Vilmen C, et al. Accurate work-rate measurements during in vivo MRS studies of exercising human quadriceps. Magnetic Resonance Materials in Physics Biology and Medicine. 2008;21(3):227–35. doi: 10.1007/s10334-008-0117-3. [DOI] [PubMed] [Google Scholar]
- 32.Layec G, Haseler LJ, Trinity JD, et al. Mitochondrial function and increased convective O2 transport: implications for the assessment of mitochondrial respiration in vivo. J Appl Physiol. 2013;115:803–11. doi: 10.1152/japplphysiol.00257.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Le Fur Y, Nicoli F, Guye M, Confort-Gouny S, Cozzone PJ, Kober F. Grid-free interactive and automated data processing for MR chemical shift imaging data. Magnetic Resonance Materials in Physics Biology and Medicine. 2010;23(1):23–30. doi: 10.1007/s10334-009-0186-y. [DOI] [PubMed] [Google Scholar]
- 34.Lind AR, Williams CA. The control of blood flow through human forearm muscles following brief isometric contractions. J Physiol. 1979;288:526–47. [PMC free article] [PubMed] [Google Scholar]
- 35.Marcora S. Perception of effort during exercise is independent of afferent feedback from skeletal muscles, heart, and lungs. J Appl Physiol. 2009;106:2060–2. doi: 10.1152/japplphysiol.90378.2008. [DOI] [PubMed] [Google Scholar]
- 36.Rossman MJ, Garten RS, Venturelli M, Amann M, Richardson RS. The role of active muscle mass in determining the magnitude of peripheral fatigue during dynamic exercise. American Journal of Physiology Regulatory, Integrative and Comparative Physiology. 2014;306:R934–R40. doi: 10.1152/ajpregu.00043.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rossman MJ, Venturelli M, McDaniel J, Amann M, Richardson RS. Muscle mass and peripheral fatigue: a potential role for afferent feedback? Acta Physiol. 2012;206(4):242–50. doi: 10.1111/j.1748-1716.2012.02471.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sidhu SK, Weavil JC, Venturelli M, et al. Spinal μ-opioid receptor-sensitive lower limb muscle afferents determine corticospinal responsiveness and promote central fatigue in upper limb muscle. J Physiol. 2014;592(22):5011–24. doi: 10.1113/jphysiol.2014.275438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sidhu SK, Weavil JC, Venturelli M, et al. Aging alters muscle reflex control of autonomic cardiovascular responses to rhythmic contractions in humans. American journal of physiology Heart and circulatory physiology. 2015;309(9):H1479–89. doi: 10.1152/ajpheart.00433.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Standl TG, Horn E, Luckmann M, Burmeister M, Wilhelm S, Schulte am Esch J. Subarachnoid sufentanil for early postoperative pain management in orthopedic patients: a placebo-controlled, double-blind study using spinal microcatheters. Anesthesiology. 2001;94(2):230–8. doi: 10.1097/00000542-200102000-00011. [DOI] [PubMed] [Google Scholar]
- 41.Trenell MI, Sue CM, Kemp GJ, Sachinwalla T, Thompson CH. Aerobic exercise and muscle metabolism in patients with mitochondrial myopathy. Muscle Nerve. 2006;33(4):524–31. doi: 10.1002/mus.20484. [DOI] [PubMed] [Google Scholar]
- 42.Vanhamme L, van den Boogart A, Van Huffel S. Improved method for accurate and efficient quantification of MRS data with use of prior knowledge. J Magn Reson. 1997;129:35–43. doi: 10.1006/jmre.1997.1244. [DOI] [PubMed] [Google Scholar]
- 43.Vanhatalo A, Black MI, DiMenna FJ, et al. The mechanistic bases of the power-time relationship: muscle metabolic responses and relationships to muscle fibre type. J Physiol. 2016;594(1):4407–23. doi: 10.1113/JP271879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Vanhatalo A, Fulford J, DiMenna FJ, Jones AM. Influence of hyperoxia on muscle metabolic responses and the power-duration work relationship during severe-intensity exercise in humans: a 31P magnetic resonance spectroscopy study. Exp Physiol. 2010;95(4):528–40. doi: 10.1113/expphysiol.2009.050500. [DOI] [PubMed] [Google Scholar]
- 45.Wigmore DM, Damon BM, Pober DM, Kent-Braun JA. MRI measures of perfusion-related changes in human skeletal muscle during progressive contractions. J Appl Physiol. 2004;97:2385–94. doi: 10.1152/japplphysiol.01390.2003. [DOI] [PubMed] [Google Scholar]
- 46.Wigmore DM, Propert K, Kent-Braun JA. Blood flow does not limit skeletal muscle force production during incremental isometric contractions. Eur J Appl Physiol. 2006;96:370–8. doi: 10.1007/s00421-005-0037-0. [DOI] [PubMed] [Google Scholar]
- 47.Wilson LB, Hand GA. The pressor reflex evoked by static contraction: neurochemistry at the site of the first synapse. Brain research Brain research reviews. 1997;23(3):196–209. doi: 10.1016/s0165-0173(96)00019-7. [DOI] [PubMed] [Google Scholar]