

Abstract
Phytochrome B (phyB) is the primary red light photoreceptor in plants, and regulates both growth and development. The relative levels of phyB in the active state are determined by the light conditions, such as direct sunlight or shade, but are also affected by light-independent dark reversion. Dark reversion is a temperature-dependent thermal relaxation process, by which phyB reverts from the active to the inactive state. Here, we show that the homologous phyB-binding proteins PCH1 and PCHL suppress phyB dark reversion, resulting in plants with dramatically enhanced light sensitivity. Moreover, far-red and blue light upregulate the expression of PCH1 and PCHL in a phyB independent manner, thereby increasing the response to red light perceived by phyB. PCH1 and PCHL therefore provide a node for the molecular integration of different light qualities by regulation of phyB dark reversion, allowing plants to adapt growth and development to the ambient environment.
Dark reversion of the red-light receptor phytochrome B (phyB) converts the receptor to the inactive state. Here, Enderle et al. show that PCH1 and PCHL bind phyB and suppress dark reversion, and that far-red and blue-light induced upregulation of PCH1 and PCHL increases red light sensitivity in Arabidopsis.
Introduction
Phytochromes are red/far-red photoreceptors in plants that play a critical role in the adaptation of growth and development to the environment1–3. Gene duplication events during evolution of seed plants resulted in small gene families coding for different types of phytochromes, including phytochromes A–E (phyA–E) in Arabidopsis 4. Phytochrome A (phyA) and phytochrome B (phyB) play a dominant role in seed plants2. PhyA is required for sensing far-red light (FR) and weak light of any wavelength, while phyB is the primary receptor for red light (R). Phytochromes exist in two states, an inactive Pr and a biologically active Pfr form that have absorption peaks in R (660 nm) and FR (730 nm), respectively. Pr and Pfr reversibly convert into each other upon absorption of light, resulting in high levels of the active Pfr form in R and low levels in FR5. In addition to light-induced Pfr → Pr reversion, the active Pfr state can also revert to the inactive Pr state in a light-independent thermal relaxation process referred to as dark reversion. The physiological activity of phyB, the primary phytochrome in light-grown and adult plants, is strongly affected by dark reversion6–8. As such, dark reversion is a major factor that determines how plants respond to light. Several phyB mutant alleles with altered dark reversion have been discovered, but despite the strong impact of dark reversion on the activity of phyB, no extragenic mutants with altered phyB dark reversion had been identified yet7–12. It is also unknown, whether plants actively regulate dark reversion of phyB to control photomorphogenic growth and development.
Recently, PHOTOPERIODIC CONTROL OF HYPOCOTYL 1 (PCH1) has been identified as a protein binding to light-activated phyB13,14. Seedlings lacking functional PCH1 have elongated hypocotyls compared to the wild type when grown in short days14. Under these conditions, PCH1 transcript and protein levels peak at dusk, enhancing phyB-dependent inactivation of the growth-promoting transcription factor PHYTOCHROME INTERACTING FACTOR 4 (PIF4)13,14. In contrast, PCH1 levels are low towards the end of the night, leading to increased PIF4 activity and hypocotyl growth14. Thus, PCH1 has been suggested to integrate clock and light signals through modulation of diurnal phyB activity. However, PCH1 is a protein of unknown molecular function and the mechanism by which it enhances phyB-dependent light responses is still unknown.
Here, we show that the pch1 pchl double mutant, which lacks functional PCH1 and a homologue, PCH1-LIKE (PCHL), displays strongly accelerated phyB dark reversion. Moreover, we show that additional signalling pathways control the expression of PCH1 and PCHL, thereby affecting the activity of phyB. Thus, PCH1 and PCHL allow plants to adapt to their environment by acting as a node of signal integration that regulates phyB dark reversion.
Results
PCH1 and PCHL interact with light-activated phyB
Using two independent approaches, yeast two-hybrid (Y2H) screening and MS/MS analyses of immunoprecipitated phyB complexes from extracts of phyB-NLS-GFP expressing Arabidopsis plants, we identified PCH1 as a phyB interacting protein (Fig. 1a, b; Supplementary Fig. 1a). PCH1 has previously been identified as a phyB interaction partner13,14. Based on homology searches, we found one potential homologue of PCH1 in Arabidopsis, PCH1-LIKE (PCHL) that was not identified in the previous reports (Supplementary Fig. 2). PCHL also interacts with phyB in the Y2H system (Fig. 1a, b); moreover, interaction of phyB with PCH1 and PCHL is confirmed by co-immunoprecipitation (Co-IP) from stable transgenic Arabidopsis plants (Fig. 1c, d; Supplementary Fig. 1b), as well as from mammalian cells (Supplementary Fig. 1c, d). Co-IP assays from plants suggest that PCH1 and PCHL preferentially interact with phyB Pfr, and a weak preference for binding to Pfr was also observed in the Y2H system.
Fig. 1.

Light-activated phyB interacts with PCH1 and PCHL. a, b Y2H detection of interaction of phyB with PCH1 and PCHL. The phyB-GAL4-activation domain (phyB-AD) fusion was co-expressed with GAL4-DNA-binding domain (BD-) fusions of PCH1 or PCHL. a X-Gal filter-lift assay. Yeast cells were lifted from chromophore-supplemented plates pre-incubated for 48 h in either constant darkness (D), red (R), or far-red light (FR). b ONPG assay. Yeast cultures supplemented with chromophore were exposed to red light, to convert phyB to Pfr, or to far-red light for phyB Pr. n = 3; error bars indicate ± s.e.m. c, d PCH1 and PCHL co-purify with phyB from native plant extracts. Four-day-old dark-grown seedlings over-expressing HA-YFP-PCH1 (PCH1ox) c or HA-YFP-PCHL (PCHLox) d in the Col-0 wild type background were either treated with red light (R, 7 μmol m−2 s−1) for 10 min or kept in darkness (D). Native total protein extracts were prepared and used for co-immunoprecipitation (Co-IP) assays. IP was performed using α-GFP antibody. α-phyB and α-GFP antibodies were used to detect endogenous phyB and YFP-tagged PCH1 and PCHL, respectively
PCH1 and PCHL extend phyB activity into the dark phase
Although PCH1 was previously shown as a phyB interactor, the molecular mechanism by which it promotes phyB-mediated light responses remained elusive14. Seedlings over-expressing HA-Yellow Fluorescent Protein (YFP)-tagged PCH1 (PCH1ox) or PCHL (PCHLox) are strongly hypersensitive to continuous R (Supplementary Fig. 3a–e), while seedlings lacking functional PCH1 and PCHL are hyposensitive (Supplementary Fig. 3f); PCH1 and PCHL have only a weak effect on hypocotyl growth in FR, consistent with a function of PCH1 and PCHL in phyB signalling (Supplementary Fig. 3g, h)14. Dark-grown PCH1ox and PCHLox seedlings are fully etiolated but we observed slightly reduced hypocotyl elongation in the dark compared to the wild type (Fig. 2a, b; Supplementary Fig. 4). Interestingly, we found that PCH1ox and PCHLox seedlings also respond to daily R pulses similar to seedlings expressing phyB mutant versions with reduced dark reversion, exhibiting short hypocotyls and open cotyledons (Fig. 2a, b; Supplementary Fig. 4)11. In contrast, wild type Col-0 seedlings are insensitive to R pulses and the response in seedlings over-expressing wild type phyB is very weak due to rapid phyB Pfr → Pr dark reversion between light pulses11; therefore, we hypothesised that PCH1 and PCHL might increase the levels of phyB Pfr by inhibiting phyB dark reversion. The total amount of phyB is critical for the responsiveness to R light15, but immunoblot analyses confirm that the amount of phyB is similar in wild type, PCH1ox, PCHLox, pch1, pchl, and pch1 pchl backgrounds (Supplementary Fig. 5). If PCH1 and PCHL inhibit phyB dark reversion but not Pfr → Pr photoconversion, then a long-wavelength FR pulse immediately following a R pulse would be expected to revert the effects of the R pulse. This was indeed the case (Fig. 2a, b; Supplementary Fig. 4), suggesting that over-expression of PCH1 and PCHL does not affect the intrinsic photoreversibility of phyB.
Fig. 2.

PCH1 and PCHL stabilise phyB in the active state in planta. a, b PCH1ox and PCHLox seedlings respond to red light pulse treatments (Rp). Wild type (Col-0) and mutant seedlings expressing either HA-YFP-PCH1 (PCH1ox) (a) or HA-YFP-PCHL (PCHLox) (b) were grown for 4 days in darkness on filter paper soaked with water. The seedlings were either treated with a single red light pulse (Rp, 5 min, 50 μmol m−2 s−1) per day, or a Rp followed by a long-wavelength FR pulse (FRp, 776 nm, 5 min, 50 μmol m−2 s−1) (Rp → FRp). Control seedlings were kept in darkness (D). See Supplementary Fig. 4 for quantification of hypocotyl lengths and experiments with seedlings grown on 0.5× MS medium. c High levels of active phyB are maintained during the dark phase in PCH1ox and PCHLox seedlings. Wild type (Col-0), PCH1ox, PCHLox, and phyB-9 seedlings were grown for 4 days in 8 h red (R, 50 μmol m−2 s−1)/16 h dark (D) cycles and given a long-wavelength far-red light pulse (FRp, 776 nm, 5 min, 50 μmol m−2 s−1) at time points after lights-off. Control seedlings were kept in darkness (D). d The end-of-day far-red (EOD-FR) response requires PCH1 and PCHL. Wild type (Col-0) and mutant seedlings were grown as in c, except either constant red light (Rc), an immediate far-red light pulse (8 h R → FRp → 16 h D), or no far-red light pulse (8 h R → 16 h D) were used. c, d Mean hypocotyl length relative to dark-grown seedlings is shown. Error bars indicate ± s.e.m.; n ≥ 20. e, f Subnuclear localisation of PCH1 and phyB was analysed by fluorescence microscopy. Scale bar = 5 µm. e PCH1 stabilises phyB photobodies. Four-day-old etiolated seedlings expressing phyB-mCer in phyB-9 or HA-YFP-PCH1 (PCH1ox) backgrounds were exposed to red light (R, 10 μmol m−2 s−1) for 8 h, followed either by 0 or 14 h incubation in darkness (D). Data for phyB-mCer single transgenic seedlings are duplicated in Supplementary Fig. 7a. f PhyB photobodies are highly unstable in the pch1 pchl mutant. Four-day-old etiolated seedlings expressing phyB-GFP in wild type or pch1 pchl backgrounds were exposed to red light (R, 50 μmol m−2 s−1) for 12 h followed by incubation in darkness (D)
To further investigate the possibility that PCH1 and PCHL allow maintenance of high levels of phyB Pfr in the dark, we grew wild type, PCH1ox, and PCHLox seedlings in 8 h R/16 h dark cycles and treated them with FR pulses either at the end of the day or 4, 8, or 12 h after light-off to trigger Pfr → Pr reversion (Fig. 2c). Hypocotyl growth in PCH1ox and PCHLox seedlings was more strongly affected than in the wild type by the reverting effect of FR pulses applied during the dark phase, suggesting that higher levels of photoreversible phyB Pfr have been maintained in the dark in seedlings over-expressing PCH1 or PCHL than in wild type seedlings. These data are consistent with the hypothesis that PCH1 and PCHL reduce phyB dark reversion.
To investigate the effect of endogenous levels of PCH1 and PCHL on the activity of phyB we used pch1, pchl, and pch1 pchl mutant seedlings for an end-of-day far-red (EOD-FR) experiment (Fig. 2d; Supplementary Fig. 6). Seedlings were grown in 8 h R/16 h dark cycles with or without a FR pulse at the end of the light phase. Wild type seedlings respond to a FR pulse at light-off with increased hypocotyl growth, which is due to phyB Pfr → Pr photoconversion16. In contrast, pch1 and pch1 pchl seedlings not treated with a FR pulse grew as tall as those exposed to a FR pulse at the end of the light phase, consistent with rapid loss of active phyB after light-off due to dark reversion. Seedlings lacking functional PCHL also exhibited increased hypocotyl growth in light/dark cycles compared to continuous light but were partially responsive to EOD-FR pulses. These data suggest that PCH1 and PCHL are necessary for robust phyB activity in vivo, possibly by reducing dark reversion of phyB and thereby stabilising phyB Pfr in darkness.
Light-activated phyB, and many phyB-binding proteins, assemble into subnuclear structures called photobodies17. Co-expression of phyB-mCerulean (mCer) and HA-YFP-PCH1 or HA-YFP-PCHL revealed that PCH1 and PCHL also co-localise with phyB in light-induced photobodies in stable transgenic Arabidopsis (Fig. 2e; Supplementary Fig. 7a). Only phyB Pfr is retained in photobodies, while phyB Pr rapidly dissociates11,18. Co-expression of PCH1 strongly accelerated the appearance of phyB-mCer photobodies after light-on and prevented dissociation after light-off, similar to previous observations (Fig. 2e; Supplementary Fig. 7b)14. Over-expression of PCHL had a qualitatively similar but weaker effect on the stability of phyB-mCer photobodies, consistent with the hypothesis that PCH1 and PCHL promote retention of phyB Pfr in darkness. Moreover, phyB photobodies were highly unstable in the pch1 pchl double mutant (Fig. 2f). Importantly, over-expression of PCH1 did not prevent the dissociation of phyB-mCer photobodies in response to a FR pulse given at light-off (Supplementary Fig. 7c), further supporting the hypothesis that PCH1 and PCHL reduce phyB dark reversion but do not affect phyB Pfr → Pr photoconversion.
PCH1 and PCHL inhibit phyB dark reversion
To directly measure the effect of PCH1 and PCHL on the in vivo ratio between active and total phyB (Pfr/Ptot), we utilised a dual-wavelength ratiospectrophotometer (hereafter referred to as a ratiospect). Only endogenous phyA levels are sufficiently high in wild type seedlings to be detected by ratiospect measurements, but phyB Pfr/Ptot levels can be measured in seedlings over-expressing phyB-GFP—either in the phyA mutant background or following a 3 h R pre-irradiation to trigger degradation of phyA11. After saturating R treatment and transfer to the dark, phyB Pfr/Ptot levels dropped more rapidly in the pch1 pchl double mutant background than in the wild type, whilst the total amount of phyB did not change (Fig. 3a, b; Supplementary Fig. 8). Moreover, co-expression of c-Myc-mCherry-PCH1 or c-Myc-mCherry-PCHL in phyB-GFP over-expressing seedlings strongly reduced phyB dark reversion following pre-irradiation with R for either 20 min or 3 h (Fig. 3c, d). Intriguingly, phyB dark reversion in PCH1ox and PCHLox seedlings was substantially slower following the 3 h R pre-irradiation in comparison to only 20 min pre-irradiation, despite both treatments establishing equal initial phyB Pfr/Ptot levels. This observation could be explained through increased phyB nuclear transport, and thus binding to the nuclear localised PCH1 and PCHL. Alternatively, more efficient assembly of phyB into photobodies that are known to form after prolonged exposure to R and to reduce phyB dark reversion could also explain the increased Pfr stability11,18–20.
Fig. 3.

PCH1 and PCHL inhibit phyB dark reversion. Dual-wavelength ratiospectrophotometer quantification of the relative abundance of phyB in the active state (Pfr/Ptot) (a, c) and total phyB (Ptot) (b, d) in planta. a, b PhyB dark reversion is accelerated in the absence of PCH1 and PCHL. Four-day-old etiolated seedlings expressing phyB-GFP in PCH1/PCHL wild type (WT; phyA-211 phyB-9 phyB-GFP) or the mutant background (pch1 pchl; phyA-211 pch1 pchl phyB-GFP) were exposed to red light (R, 10 μmol m−2 s−1) for 3 h followed by incubation in darkness (D) for indicated times. c, d PhyB dark reversion is reduced in PCH1ox and PCHLox seedlings. Four-day-old etiolated seedlings expressing c-Myc-mCherry-PCH1 (PCH1ox) or c-Myc-mCherry-PCHL (PCHLox) in the phyB-GFP background were pre-irradiated with red light (R, 20 μmol m−2 s−1) for either 20 min (black lines and symbols) or 3 h (grey lines and symbols) followed by incubation in darkness (D). a–d Data are means of ≥ 4 biological replicates; error bars indicate ±s.e.m.
PCH1 and PCHL function in molecular signal integration
Signal integration is critical for the adaptation of growth and development of plants to diverse and rapidly changing environmental conditions. Given that PCH1 and PCHL regulate the activity of phyB, they may form a node for input for other signalling pathways upon responses to R. Using qPCR analyses we found that both PCH1 and PCHL expression in dark-grown seedlings is transiently upregulated following transition to R, whilst transition to FR or blue light (B) results in sustained upregulation of PCH1 and PCHL transcript levels (Supplementary Fig. 9). This expression pattern is also observed in PCH1 and PCHL promoter-GUS reporter lines, showing PCH1 and PCHL promoter activity primarily in cotyledons and parts of the hypocotyl, and for PCH1 also in roots (Supplementary Fig. 10a). In 14-day-old plants grown in either long-day or short-day conditions, PCH1 and PCHL promoter activity is detected in leaves, and for PCH1 in roots (Supplementary Fig. 10b). Interestingly, PCH1 promoter activity in roots appears not to be regulated by light. Using pch1 seedlings complemented with pPCH1:HA-YFP-PCH1 (Supplementary Fig. 3i), we also confirmed light-induced accumulation of PCH1 protein in FR, B, and R (Fig. 4a, b). Importantly, light regulation of PCH1 and PCHL transcript levels does not require phyB but depends on phyA in R and FR and on both phyA and cryptochromes in B (Fig. 4c, d; Supplementary Figs. 11 and 12). The phyA protein is degraded in R with a half-life of about 30 min, while degradation is much slower in FR21. Thus, rapid degradation of phyA in R might be the reason why upregulation of PCH1 and PCHL in R is only transient. We hypothesise that FR and B signalling may integrate with phyB signalling through PCH1 and PCHL, given that the expression of PCH1 and PCHL is upregulated in response to FR and B.
Fig. 4.

PCH1 and PCHL contribute to molecular signal integration. a, b PCH1 protein accumulates in far-red and blue light. a Four-day-old etiolated pch1 seedlings expressing HA-YFP-PCH1 from its native promoter were either kept in darkness (D), or exposed to far-red (FR, 25 μmol m−2 s−1), blue light (B, 25 μmol m−2 s−1), or red light (R, 25 μmol m−2 s−1) for 8 h and harvested. Etiolated HA-YFP-PCH1 over-expressing seedlings (PCH1ox) were used for comparison. Total protein was extracted and used for immunoblotting; α-HA and α-actin antibodies were used for detection of HA-YFP-PCH1 and actin (loading control). b pch1 seedlings expressing HA-YFP-PCH1 from its native promoter were grown as described in a and used for fluorescence microscopy. Scale bar = 5 µm. c, d Regulation of PCH1 and PCHL expression by red and far-red light requires phyA. Four-day-old dark-grown wild type (Col-0) and mutant seedlings were either kept in darkness (D) or exposed to red (R, 7.5 μmol m−2 s−1) or far-red light (FR, 6.7 μmol m−2 s−1) for 1 h. Total RNA was extracted and qRT-PCR was performed using probes specific for either PCH1 (c) or PCHL (d). ACT1 was used as internal control and expression of PCH1 and PCHL is shown relative to expression of ACT1. Data are means of three biological replicates; error bars indicate ± s.d. Biological replicates are shown in Supplementary Fig. 11; data in c and d correspond to Supplementary Fig. 11d and h. e, f PCH1 and PCHL are required for phyB responsiveness amplification by far-red and blue light. Dark-grown wild type (Col-0) and mutant seedlings were pre-treated with either far-red (FR, 25 μmol m−2 s−1) (e) or blue light (B, 25 μmol m−2 s−1) (f) and given a red light pulse (Rp, 5 min, 50 μmol m−2 s−1), either followed by a long-wavelength far-red light pulse (FRp, 776 nm, 5 min, 50 μmol m−2 s−1) or darkness; these light pulses were repeated after 24 h. Mean hypocotyl length relative to dark-grown seedlings is shown. Error bars indicate ± s.e.m.; n ≥ 20. See Supplementary Fig. 13 for seedling photographs and detailed explanation of light treatments
The responsiveness of etiolated wild type seedlings to R pulses has been reported to be strongly increased by pre-treatments with FR or B for 8 h22,23. In contrast, we found that PCH1ox and PCHLox seedlings are sensitive to R pulses even without FR or B pre-irradiation (Fig. 2a, b; Supplementary Fig. 4), while R pulses did not further promote hypocotyl growth inhibition and cotyledon unfolding in phyA-211 and pch1 pchl mutant seedlings pre-treated with FR or B (Fig. 4e, f; Supplementary Fig. 13). Thus, induction of PCH1 and PCHL expression triggered through FR-induced and B-induced phyA signalling, can enhance phyB-mediated R responses by suppressing phyB dark reversion and thereby preventing inactivation of phyB Pfr (Fig. 5). It is tempting to speculate that also other signalling pathways could control the activity of phyB through regulation of PCH1 and PCHL, which might be critical for the adaptation of plants to changes in the ambient environment.
Fig. 5.
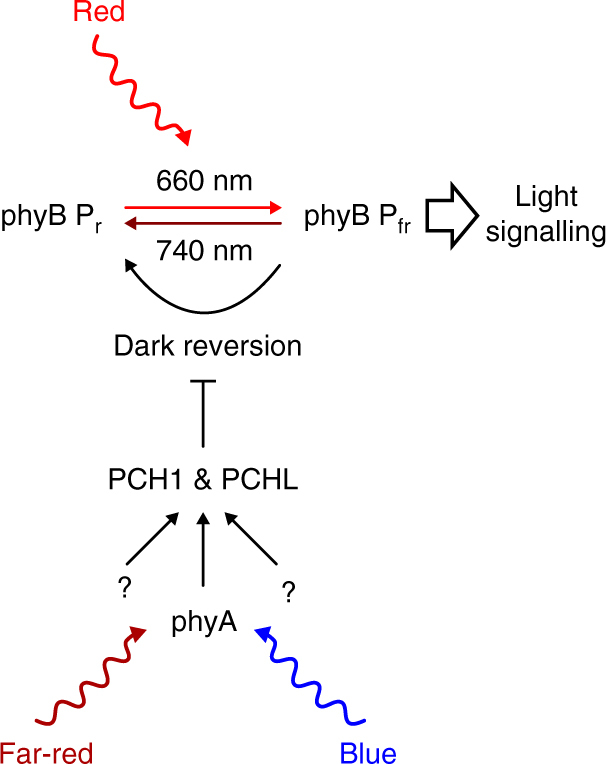
Model for PCH1-dependent and PCHL-dependent regulation of phyB responses. PCH1 and PCHL reduce phyB dark reversion. Blue and far-red light perceived by phyA induce expression of PCH1 and PCHL, and thereby allow seedlings to respond to red light pulses (phyB responsiveness amplification by far-red and blue light). We speculate that also other signalling pathways could control the activity of phyB through regulation of PCH1 and PCHL, which might play an important role in integration of phyB-dependent light signalling with other signalling pathways
Discussion
Dark reversion experiments performed for phyB expressed in E. coli, yeast and Arabidopsis demonstrate that dark reversion can be extensively modulated by external factors although it is an intrinsic property of the phyB molecule8,18,24–26. Here, we provide genetic, photo-biological and biochemical evidence that PCH1 and PCHL suppress phyB dark reversion. Importantly, the effect of PCH1 and PCHL on phyB dark reversion does not depend on over-expression; we show that endogenous levels of PCH1 and PCHL are sufficient to control the activity of phyB through regulation of dark reversion, establishing PCH1 and PCHL as key players in photomorphogenesis and signal integration.
Two fundamentally different models can be envisaged to explain how PCH1 and PCHL suppress dark reversion of phyB. PCH1 and PCHL could either directly stabilise the Pfr conformation of phyB to prevent reversion to the inactive Pr state or they could induce molecular events that indirectly lead to stabilisation of phyB in the Pfr state. Phosphorylation at serine residue 86, for instance, accelerates phyB dark reversion8 and PCH1/PCHL hypothetically could either prevent phosphorylation or promote dephosphorylation. It has also been demonstrated that phyB bound to photobodies is protected from dark reversion18; therefore, another hypothetical mechanism for the PCH1/PCHL-mediated suppression of phyB dark reversion is that PCH1/PCHL promote the association of phyB with photobodies, where other factors could stabilise phyB in the Pfr state.
Over-expression of PCH1 or PCHL strongly suppress dark reversion of phyB. A similar but much weaker effect on phyB dark reversion has been observed for ARABIDOPSIS RESPONSE REGULATOR 4 (ARR4). Seedlings over-expressing ARR4 are slightly hypersensitive to R, possibly due to reduced phyB dark reversion. However, dark reversion has not been investigated in the arr4 mutant; therefore, it is still unknown if endogenous levels of ARR4 are sufficient to regulate dark reversion of phyB26,27. This is in contrast to PCH1 and PCHL, which strongly suppress phyB dark reversion even when expressed at wild type levels. To our knowledge, the pch1 pchl mutant is the first extragenic mutant shown to be affected in phyB dark reversion.
Though both PCH1 and PCHL delay phyB dark reversion, they do not contribute equally to physiological responses to light. It is possible that differences in expression patterns or light regulation of expression could be responsible for these differences. Furthermore, PCH1 and PCHL share only 32% identity in amino acid sequence and PCHL lacks two motifs conserved between PCH1 and sequences found in other species (corresponding to PCH1 amino acid residues 66–76 and 210–224), which could contribute to different biological activity.
Wild type seedlings are normally insensitive to daily R pulses but respond to this treatment upon pre-irradiation with FR or B11,22,23. This responsiveness amplification of phyB by FR and B depends on PCH1 and PCHL. Transcript levels of PCH1 and PCHL are upregulated by phyA in response to FR and B, leading to suppression of phyB dark reversion and increased sensitivity to light (Fig. 5). Hypothetically, phyA-induced expression of PCH1 and PCHL could further promote light responses by enhancing phyB downstream signalling, though experimental evidence is still lacking. The advantage of FR/B-dependent phyB responsiveness amplification for plants growing under natural conditions is unclear; however, this phenomenon demonstrates that plants actively control dark reversion of phyB to regulate the sensitivity to light.
We speculate that other signalling pathways could control PCH1 and PCHL and thereby integrate with phyB-mediated light responses to allow plants to adapt growth and development to the ambient environment. Moreover, phyB dark reversion is temperature-dependent and recent reports have shown that plants use this temperature dependency as a thermo-sensing mechanism28,29. Thus, PCH1 and PCHL could also play a role in integrating ambient light and temperature cues by controlling phyB dark reversion.
Methods
Vectors and cloning
Details on the cloning of plasmid constructs and primers used in this study can be found in the Supplementary Note 1.
Co-immunoprecipitation from mammalian cell culture
HEK293T cells (provided by the BIOSS Toolbox, BIOSS Centre for Biological Signalling Studies, University of Freiburg), were cultivated in DMEM medium (PAN, Aidenbach, Germany) supplemented with 10% FCS (PAN, Aidenbach, Germany), 2 mM l-glutamine, 100 U/ml penicillin, and 0.1 mg/ml streptomycin (PAN, Aidenbach, Germany), at 37 °C, 5% CO2, and high humidity. For transfection cells were seeded at 80–90% confluency in a 1:6 ratio in 10 cm tissue culture plates (Greiner Bio-One) the day prior transfection. Eight micrograms of total DNA was diluted in 560 µl of OptiMEM (Invitrogen), mixed with 24 µl of polyethylene–imine solution (1 mg/ml stock concentration; linear; MW: 25 kDa; Polyscience, Warrington, USA) and incubated for 15 min at room temperature before drop-wise addition to cells. Following 24 h, medium was replaced with fresh medium supplemented with 15 µM phycocyanobilin (Livchem #FSIP14137, Frankfurt/Main, Germany), and returned to incubate in darkness for a further 24 h prior to lysis.
The following steps were performed under green light. Cells were washed with 5 ml of DPBS (PAN, Aidenbach, Germany), and then lysed for 20 min on ice in 2.5 ml lysis buffer (100 mM NaPO4 pH 7.8, 150 mM NaCl, 1 mM KCl, 1 mM EDTA, 1% (w/v) PEG4000, 50 µM MG132 (Sigma-Aldrich), 0.5% Triton-X100, 1 mM Na3VO4, 2 mM Na4P2O7, 10 mM NaF, 1× protease inhibitor mixture (1 tablet per 50 ml; Roche)). Lysate was then centrifuged at 20,000×g for 20 min at 4 °C, and supernatant used immediately for co-immunoprecipitation.
Fifty microlitres of anti-GFP microbeads (Miltenyi Biotec, Bergisch-Gladbach, Germany; Cat-No: 130-091-125) were added to 2.5 ml of lysate, protected from light, and incubated at 4 °C for 2 h. Co-immunoprecipitation was performed following the procedure of the µMACS Anti-GFP Isolation Kit (Miltenyi Biotec, Bergisch-Gladbach, Germany; Cat-No: 130-091-125). Samples were separated by SDS-PAGE, and analysed by immunoblot using a monoclonal α-GFP antibody (Roche; Cat-No: 11814460001, monoclonal, mouse, dilution 1:2000) and a monoclonal α-c-Myc antibody (Abcam, Cambridge, UK; Cat-No: ab32, monoclonal, mouse, dilution 1:5000), respectively, and detected using ECLprime reagent (GE Healthcare).
Plant material
All Arabidopsis thaliana lines used were in Col-0 background. The pch1 (T-DNA insertion line; SALK_024229), phyA-211, phyB-9, cry1-304, and cry2-1 mutants have been described previously14,30–34; the phyA-211 phyB-9 double mutant was generated by crossing the respective single mutants. The pchl mutant line contains a T-DNA insertion within the coding region of PCHL (AT4G34550) and was provided by the Nottingham Arabidopsis Stock Centre (SALK_206946C; #N696800). The pch1 pchl double mutant was generated by crossing the respective single mutants. Genotyping of pch1 and pchl was performed using primers DS245/DS246 (wild type PCH1), DS246/LBb1.3 (T-DNA insertion in pch1), BE078/BE079 (wild type PCHL), and BE078/LBb1.3 (T-DNA insertion in pchl). The sequence of primers used for genotyping pch1 and pchl can be found in the Supplementary Note 1.
The phyB-GFP over-expression line (p35S:PHYB-GFP) used for Pfr and dark reversion measurements in the dual-wavelength ratiospectrophotometer (ratiospect) (Fig. 3) is in phyA-211 phyB-9 double mutant background and has been described previously6. This line was crossed to pch1 pchl to generate phyB-GFP phyA-211 pch1 pchl, which was used for ratiospect measurements (Fig. 3), Western blot quantification of phyB-GFP and endogenous phyB (Supplementary Fig. 8), and analysis of phyB photobodies in pch1 pchl (Fig. 2f). This line still contains endogenous phyB, which considerably contributes to total phyB levels and detection of phyB by ratiospect measurements (Supplementary Fig. 8).
The Agrobacterium tumefaciens floral dip transformation method35,36 was used for generating PCH1ox and PCHLox lines. HA-YFP-PCH1ox (p35S:HA-YFP-PCH1:terRbcS; plasmid pDS366) and HA-YFP-PCHLox (p35S:HA-YFP-PCHL:terRbcS; plasmid pBE52c) were transformed into the Col-0 background, and subsequently crossed into phyA-211, phyB-9, and phyA-211 phyB-9 backgrounds. These lines were used for all physiological experiments, the Co-IP assays shown in Fig. 1, and the Western blots in Supplementary Fig. 5.
PCH1ox (p35S:myc-mCherry-PCH1:terRbcS; plasmid pBE50) and PCHLox (p35S:myc-mCherry-PCHL:terRbcS; plasmid pBE60) were transformed into the above phyB-GFP line6. These lines were used for ratiospect measurements of Pfr levels (Fig. 3) and the Co-IP assay in Supplementary Fig. 1b.
For native level expression of PCH1, pPCH1:HA-YFP-PCH1 (pBE48) was transformed into the pch1 line using the floral dip method35,36. These lines were used in Fig. 4a (line #2) and 4b (line #2), as well as in Supplementary Fig. 3i (lines #1 and #2).
A phyB-mCer (pPHYB:PHYB-mCer:terRbcS, pPPO43B-PHYB) line that over-expresses phyB was generated in the phyB-9 background using the floral dip method35,36, and crossed to PCH1ox phyA-211 and PCHLox phyA-211. These lines were used for the experiments shown in Fig. 2e and Supplementary Fig. 7.
To generate phyB-NLS-GFP lines for purification of phyB complexes used for MS/MS analyses, pART27 p35S:PHYB-NLS-GFP was transformed into phyA-211 phyB-9 double mutant background using the floral dip method35. Homozygous lines were selected on kanamycin for further experiments.
PCH1 and PCHL promoter-GUS reporter constructs (plasmids pBE26new and pBE75) were transformed into Col-0 using the floral dip method35,36. Homozygous lines were selected by spraying T2 seedlings with BASTA. These lines were used for the experiments shown in Supplementary Fig. 10.
Measurement of hypocotyl length
Seedlings were grown in Petri dishes on four sheets of filter paper (Macherey-Nagel; Cat-No: MN 615) soaked with sterile water. Seeds were first stratified at 4 °C in darkness for 4–7 days, then germination induced by incubation in white light (70 µmol m−2 s−1) at 22 °C for 4–8 h. Following 16 h in darkness at 22 °C, seedlings were given a long-wavelength FR pulse (776 nm, 5 min, 50 µmol m−2 s−1), to inactivate all phytochromes before transfer to various conditions, which are described in figure legends. The hypocotyl length of at least 20 individual seedlings was measured at the fourth day following germination induction. Error bars represent the standard error of the population. Seedlings for the experiments shown in Supplementary Fig. 4d and f were grown in Petri dishes on four sheets of filter paper soaked with sterile 0.5× MS medium under the conditions described above.
Histochemical GUS stain of Arabidopsis seedlings and plants
Three-day-old dark-grown seedlings treated for 24 h with either R (660 nm, 25 µmol m−2 s−1), FR (740 nm, 25 µmol m−2 s−1), B (450 nm, 25 µmol m−2 s−1), or white light (100 µmol m−2 s−1), or 14-day-old seedlings grown in either long (16 h white light, 100 µmol m−2 s−1/8 h dark) or short day conditions (8 h white light, 100 µmol m−2 s−1/16 h dark) were submerged in 2 ml fixative solution (0.3 M mannitol, 10 mM MES, 0.3% formaldehyde, pH 5.6) for 45 min. The seedlings were then incubated in washing solution (50 mM Na2HPO4, pH 7) for 45 min on a shaker before transfer to the staining solution (50 mM Na2HPO4, 0.05% X-Gluc [Carl Roth, Germany], 2 mM K3Fe(CN)6, 2 mM K4Fe(CN)6, pH 7). Seedlings were vacuum infiltrated for 20 min and incubated over night at room temperature (22 °C). Then, seedlings were destained stepwise using a dilution series of 50%, 60% and finally 70% ethanol. Pictures were taken using a camera-equipped binocular microscope from Zeiss, Germany. Magnification was 6.4× for whole seedlings, 25.6× for leaves and 40× for roots.
Quantification of transcript levels
Four-day-old dark-grown seedlings were either kept in darkness or exposed to the respective light conditions for the indicated time. Total RNA was extracted using the concert plant RNA purification reagent (Invitrogen) followed by a RNA clean up using the RNAeasy kit (Qiagen). cDNA was prepared using the High Capacity Reverse Transcription kit (Applied Biosystems, Thermo Scientific). qRT-PCR was performed using Taqman probes specific for PCH1, PCHL, or ACT1 (AT1G32200; internal control) and a 2× high ROX master mix (Eurogentec, Belgium). Relative PCH1 or PCHL gene expression was calculated in comparison to ACT1 expression. The sequence of primers and probes used for qRT-PCR can be found in the Supplementary Note 1.
In vivo spectrometric analyses
Dark reversion of phyB over-expression lines was measured in living seedlings in a dual-wavelength ratiospectrophotometer (ratiospect). Four-day-old dark-grown seedlings were irradiated for either 20 min or 3 h with saturating red light (20 µmol m−2 s−1), then returned to darkness (22 °C) for indicated times. Seedlings were transferred to ice and 100–140 mg of seedlings transferred to chilled ratiospect cuvettes. Ratiospect measurements were performed at 0.3–0.8 °C to avoid the effects of temperature. Each time point represents the average of at least four independent biological samples. The total amount of phytochrome (Ptot) and Pfr/Ptot values were calculated as previously described37.
Co-immunoprecipitation from Arabidopsis
Four-day-old dark-grown seedlings were either treated with red light (7 µmol m−2 s−1) for 10 min or kept in the dark. Immunopreciptiations were performed using IP buffer (50 mM Tris/HCl pH 7.5, 150 mM NaCl, 0.1% NP-40, 1× Protease inhibitor cocktail, 1 mM PMSF) and 1 µg α-GFP antibody (Abcam; Cat-No: ab6556, polyclonal, rabbit) bound to 20 µl Dynabeads (Life Technologies; Cat-No: 10002D). Following 1 h incubation in darkness, immunoprecipitated complexes were washed three times with IP buffer. Elutions were analysed by SDS-PAGE, and immunoblotting with α-phyB (monoclonal, mouse, B6-B338, dilution 1:250), α-c-Myc (Invitrogen; Cat-No: MA1-980, monoclonal, mouse, dilution 1:1000), and α-GFP (Invitrogen; Cat-No: A-11120, monoclonal, mouse, dilution 1:1000).
Microscopy
For epifluorescence microscopy seedlings were transferred to glass slides under dim green light. Epifluorescence microscopy was performed using an Axioplan microscope (Zeiss, Oberkochem, Germany) equipped with filter sets for GFP (Z13, excitation 470 nm, emission 493 nm; Zeiss), YFP (F31-028, excitation 500 nm, emission 515 nm; AHF Analysentechnik, Tübingen, Germany), and CFP (F31-044, excitation 436 nm, emission 455 nm; AHF Analysentechnik, Tübingen, Germany).
Arabidopsis protein extraction and immunoblot analysis
Dark-grown seedlings were exposed to light as described in the figure legends. Total protein was extracted from harvested seedlings using preheated (95 °C) denaturing buffer (4 M Urea, 65 mM Tris/HCl pH 7.3, 3% SDS, 10% glycerol, 0.05% bromphenol blue, 10 mM DTT), 40–60 µg protein separated by 10% SDS-PAGE, and transferred to PVDF membranes. Membranes were probed with α-phyB antibody (monoclonal, mouse, B6-B338, 1:1000 dilution), α-HA-tag (Sigma-Aldrich; Cat-No: H3663, monoclonal, mouse, 1:3000 dilution), α-actin (Sigma-Aldrich; Cat-No: A0480, monoclonal, mouse, 1:3000 dilution), and detected with alkaline phosphatase-coupled α-mouse IgG (Vector Labs, Burlingame, CA, USA; Cat-No: AP-2000, 1:10,000 dilution) using CDP-Star substrate (Roche Diagnostics) and a camera detection system (Fusion SL, Vilber-Lourmat, Germany). Quantification of immunblot signals in Supplementary Figs. 5 and 8 was done as previously described (Nicole Smalley; OpenWetWare—Protein Quantification Using ImageJ; http://www.openwetware.org/index.php?title=Protein_Quantification_Using_ImageJ&oldid=700558; 2017).
Mass-spectrometry identification of PCH1
Transgenic plants expressing phyB-NLS-GFP were germinated and grown in the dark for 4 days before irradiating for 10 min with red light (7 µmol m−2 s−1). Seedlings were harvested and ground in liquid nitrogen. Total protein was solubilised and immunoprecipitated in the IP buffer (50 mM Tris/HCl pH 7.5, 150 mM NaCl, 0.1% NP-40, 1× Protease inhibitor cocktail (Sigma-Aldrich, P9599), 1 mM PMSF) by 5 µg α-GFP antibody (Abcam; Cat-No: ab6556, polyclonal, rabbit) bound to 100 µl Dynabeads (Life Technologies; Cat-No: 10002D). Immunoprecipitation was performed with the same method as described in the Co-IP from plants. The collected beads were sent for liquid MS/MS analyses at JadeBio Inc., La Jolla, CA.
Yeast two-hybrid methods
Yeast two-hybrid assays were performed as previously described39. Briefly, yeast were cultured in or on media lacking leucine and tryptophan that was supplemented with 20 µM phycocyanobilin (Livchem #FSIP14137, Frankfurt/Main, Germany). Plates were incubated under constant red light (660 nm, 1.3 µmol m−2 s−1) or far-red light (740 nm, 13 µmol m−2 s−1) for 48 h at 26 °C for X-Gal filter lift assays. o-Nitrophenol-β-d-galactopyranoside assays were performed using yeast log-phase cultures exposed to either 10 min red or far-red light (7 µmol m−2 s−1).
Image processing
Uncropped gel and blot images are shown in Supplementary Figs. 14–22.
Data availability
The authors declare that all data supporting the findings of this study are available within the paper and its supplementary information files.
Electronic supplementary material
Acknowledgements
This study was supported by the Excellence Initiative of the German Federal and State Governments (EXC 294/BIOSS to A.H. and M.H.U. and GSC-4/Spemann Graduate School to P.S.) and grants from the German Research Foundation (HI 1369/5-1 and HI 1369/7-1) to A.H., the Human Frontier Science Programme (HFSP; RGP0025/2013) to A.H. and E.H., and the National Institute of Health (NIH) (1R01 GM-114297) to E.H.; B.E. was supported by an LGFG (Landesgraduiertenförderung Baden-Württemberg) scholarship. We thank Prof. Jorge J. Casal (University of Buenos Aires) and Prof. Eberhard Schäfer (University of Freiburg) for discussion, Martina Krenz (University of Freiburg, Germany) for excellent technical work, and the BIOSS Toolbox for providing HEK293T cells.
Author contributions
B.E. performed all physiological experiments, microscopy, ratiospect measurements, Y2H assays, and generated all transgenic lines and crossings; D.J.S. did the Y2H screen for phytochrome-interacting proteins; B.E. and P.K.K. performed qPCR experiments; I.P. did Co-IPs from plant extracts and MS/MS; P.S. performed Co-IPs from mammalian cells; C.K. did Western blot for endogenous phyB in pch1 pchl; M.H.U., E.H., and A.H. conceived the study; B.E., E.H., and A.H. wrote the manuscript; all authors discussed and commented on the manuscript.
Competing interests
The authors declare no competing financial interests.
Footnotes
Electronic supplementary material
Supplementary Information accompanies this paper at 10.1038/s41467-017-02311-8.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Casal JJ. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant. Biol. 2013;64:403–427. doi: 10.1146/annurev-arplant-050312-120221. [DOI] [PubMed] [Google Scholar]
- 2.Li J, Li G, Wang H, Deng XW. Phytochrome signaling mechanisms. Arab. Book Am. Soc. Plant Biol. 2011;9:e0148. doi: 10.1199/tab.0148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kami C, Lorrain S, Hornitschek P, Fankhauser C. Light-regulated plant growth and development. Curr. Top. Dev. Biol. 2010;91:29–66. doi: 10.1016/S0070-2153(10)91002-8. [DOI] [PubMed] [Google Scholar]
- 4.Li FW, et al. Phytochrome diversity in green plants and the origin of canonical plant phytochromes. Nat. Commun. 2015;6:7852. doi: 10.1038/ncomms8852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mancinelli, A. L. in Photomorphogenesis inPlants (eds Kendrick, R. E. & Kronenberg, G. H. M.) 211–269 (Kluwer Academic Publishers, Dordrecht, 1994).
- 6.Klose C, et al. Systematic analysis of how phytochrome B dimerization determines its specificity. Nat. Plants. 2015;1:15090. doi: 10.1038/nplants.2015.90. [DOI] [PubMed] [Google Scholar]
- 7.Kretsch T, Poppe C, Schäfer E. A new type of mutation in the plant photoreceptor phytochrome B causes loss of photoreversibility and an extremely enhanced light sensitivity. Plant J. 2000;22:177–186. doi: 10.1046/j.1365-313x.2000.00715.x. [DOI] [PubMed] [Google Scholar]
- 8.Medzihradszky M, et al. Phosphorylation of phytochrome B inhibits light-induced signaling via accelerated dark reversion in Arabidopsis. Plant Cell. 2013;25:535–544. doi: 10.1105/tpc.112.106898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nito K, Wong CCL, Yates JR, Chory J. Tyrosine phosphorylation regulates the activity of phytochrome photoreceptors. Cell Rep. 2013;3:1970–1979. doi: 10.1016/j.celrep.2013.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang J, Stankey RJ, Vierstra RD. Structure-guided engineering of plant phytochrome B with altered photochemistry and light signaling. Plant Physiol. 2013;161:1445–1457. doi: 10.1104/pp.112.208892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ádám Eacute, et al. Altered dark- and photoconversion of phytochrome B mediate extreme light sensitivity and loss of photoreversibility of the phyB-401 mutant. PLoS One. 2011;6:e27250. doi: 10.1371/journal.pone.0027250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Elich TD, Chory J. Biochemical characterization of Arabidopsis wild-type and mutant phytochrome B holoproteins. Plant Cell. 1997;9:2271–2280. doi: 10.1105/tpc.9.12.2271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Huang H, et al. Identification of evening complex associated proteins in Arabidopsis by affinity purification and mass spectrometry. Mol. Cell. Proteom. 2016;15:201–217. doi: 10.1074/mcp.M115.054064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huang H, et al. PCH1 integrates circadian and light-signaling pathways to control photoperiod-responsive growth in Arabidopsis. eLife. 2016;5:1–27. doi: 10.7554/eLife.13292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Leivar P, et al. The Arabidopsis phytochrome-interacting factor PIF7, together with PIF3 and PIF4, regulates responses to prolonged red light by modulating phyB levels. Plant Cell. 2008;20:337–352. doi: 10.1105/tpc.107.052142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nagatani A, Reed JW, Chory J. Isolation and initial characterization of Arabidopsis mutants that are deficient in phytochrome A. Plant Physiol. 1993;102:269–277. doi: 10.1104/pp.102.1.269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Van Buskirk EK, Decker PV, Chen M. Photobodies in light signaling. Plant Physiol. 2012;158:52–60. doi: 10.1104/pp.111.186411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rausenberger J, et al. An integrative model for phytochrome B mediated photomorphogenesis: from protein dynamics to physiology. PLoS One. 2010;5:e10721. doi: 10.1371/journal.pone.0010721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kircher S, et al. Nucleocytoplasmic partitioning of the plant photoreceptors phytochrome A, B, C, D, and E is regulated differentially by light and exhibits a diurnal rhythm. Plant Cell. 2002;14:1541–1555. doi: 10.1105/tpc.001156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bauer D, et al. Constitutive Photomorphogenesis 1 and multiple photoreceptors control degradation of Phytochrome Interacting Factor 3, a transcription factor required for light signaling in Arabidopsis. Plant Cell. 2004;16:1433–1445. doi: 10.1105/tpc.021568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hennig L, Büche C, Eichenberg K, Schäfer E. Dynamic properties of endogenous phytochrome A in Arabidopsis seedlings. Plant Physiol. 1999;121:571–578. doi: 10.1104/pp.121.2.571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hennig L, Poppe C, Unger S, Schäfer E. Control of hypocotyl elongation in Arabidopsis thaliana by photoreceptor interaction. Planta. 1999;208:257–263. doi: 10.1007/s004250050557. [DOI] [PubMed] [Google Scholar]
- 23.Casal JJ, Boccalandro H. Co-action between phytochrome B and HY4 in Arabidopsis. Planta. 1995;197:213–218. doi: 10.1007/BF00202639. [DOI] [PubMed] [Google Scholar]
- 24.Oka Y, et al. Functional analysis of a 450-amino acid N-terminal fragment of phytochrome B in Arabidopsis. Plant Cell. 2004;16:2104–2116. doi: 10.1105/tpc.104.022350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Eichenberg K, et al. Arabidopsis phytochromes C and E have different spectral characteristics from those of phytochromes A and B. FEBS Lett. 2000;470:107–112. doi: 10.1016/S0014-5793(00)01301-6. [DOI] [PubMed] [Google Scholar]
- 26.Sweere U, et al. Interaction of the response regulator ARR4 with phytochrome B in modulating red light signaling. Science. 2001;294:1108–1111. doi: 10.1126/science.1065022. [DOI] [PubMed] [Google Scholar]
- 27.Mira-Rodado V, et al. Functional cross-talk between two-component and phytochrome B signal transduction in Arabidopsis. J. Exp. Bot. 2007;58:2595–2607. doi: 10.1093/jxb/erm087. [DOI] [PubMed] [Google Scholar]
- 28.Legris M, et al. Phytochrome B integrates light and temperature signals in Arabidopsis. Science. 2016;354:897–900. doi: 10.1126/science.aaf5656. [DOI] [PubMed] [Google Scholar]
- 29.Jung JH, et al. Phytochromes function as thermosensors in Arabidopsis. Science. 2016;354:886–889. doi: 10.1126/science.aaf6005. [DOI] [PubMed] [Google Scholar]
- 30.Franklin KA, Quail PH. Phytochrome functions in Arabidopsis development. J. Exp. Bot. 2010;61:11–24. doi: 10.1093/jxb/erp304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Reed JW, Nagpal P, Poole DS, Furuya M, Chory J. Mutations in the gene for the red/far-red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell. 1993;5:147–157. doi: 10.1105/tpc.5.2.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Reed JW, Nagatani A, Elich TD, Fagan M, Chory J. Phytochrome A and phytochrome B have overlapping but distinct functions in Arabidopsis development. Plant Physiol. 1994;104:1139–1149. doi: 10.1104/pp.104.4.1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mockler TC, Guo H, Yang H, Duong H, Lin C. Antagonistic actions of Arabidopsis cryptochromes and phytochrome B in the regulation of floral induction. Development. 1999;126:2073–2082. doi: 10.1242/dev.126.10.2073. [DOI] [PubMed] [Google Scholar]
- 34.Guo H, Yang H, Mockler TC, Lin C. Regulation of flowering time by Arabidopsis photoreceptors. Science. 1998;279:1360–1363. doi: 10.1126/science.279.5355.1360. [DOI] [PubMed] [Google Scholar]
- 35.Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16:735–743. doi: 10.1046/j.1365-313x.1998.00343.x. [DOI] [PubMed] [Google Scholar]
- 36.Davis AM, Hall A, Millar AJ, Darrah C, Davis SJ. Protocol: Streamlined sub-protocols for floral-dip transformation and selection of transformants in Arabidopsis thaliana. Plant Methods. 2009;5:3. doi: 10.1186/1746-4811-5-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gross, J., Seyfried, M., Fukshansky, L. & Schäfer, E. in Methods in Photomorphogenesis, BiologicalTechnique Series (eds Smith, H. & Holmes, M.G.) 131–157 (Academic Press, London, 1984).
- 38.Hirschfeld M, Tepperman JM, Clack T, Quail PH, Sharrock RA. Coordination of phytochrome levels in phyB mutants of Arabidopsis as revealed by apoprotein-specific monoclonal antibodies. Genetics. 1998;149:523–535. doi: 10.1093/genetics/149.2.523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sheerin DJ, et al. Light-activated phytochrome A and B interact with members of the SPA family to promote photomorphogenesis in Arabidopsis by reorganizing the COP1/SPA complex. Plant Cell. 2015;27:189–201. doi: 10.1105/tpc.114.134775. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The authors declare that all data supporting the findings of this study are available within the paper and its supplementary information files.