

Abstract
The amygdala is a limbic brain region that plays a key role in emotional processing, neuropsychiatric disorders, and the emotional-affective dimension of pain. Preclinical and clinical studies have identified amygdala hyperactivity as well as impairment of cortical control mechanisms in pain states. Hyperactivity of basolateral amygdala (BLA) neurons generates enhanced feedforward inhibition and deactivation of the medial prefrontal cortex (mPFC), resulting in pain-related cognitive deficits. The mPFC sends excitatory projections to GABAergic neurons in the intercalated cell mass (ITC) in the amygdala, which project to the laterocapsular division of the central nucleus of the amygdala (CeLC; output nucleus) and serve gating functions for amygdala output. Impairment of these cortical control mechanisms allows the development of amygdala pain plasticity. Mechanisms of abnormal amygdala activity in pain with particular focus on loss of cortical control mechanisms as well as new strategies to correct pain-related amygdala dysfunction will be discussed in the present review.
1. The Amygdala and Pain
The amygdala is an almond-shaped limbic structure located in the medial temporal lobe and is well known for its role in conveying emotional significance to a sensory stimulus, emotional and affective states, and related behavioral adaptations in response to changes in the internal and external bodily environment [1–4]. The amygdala has also emerged as an important site in the brain for the emotional-affective dimension of pain and pain modulation [5–12].
A pain-related function was first suggested by the discovery of a dedicated nociceptive pathway from the spinal cord through the external lateral parabrachial (PB) nucleus to the central nucleus of the amygdala [13, 14]. Reevaluation of an historical example of reduced pain sensitivity also suggests amygdala involvement in pain processing. Patient H.M. was a man that underwent bilateral resection of the temporal lobe including the uncus, amygdala, anterior hippocampus, and parahippocampal gyrus to correct severe and intractable epilepsy [15–17]. After the surgery, H.M. did not perceive even the highest thermal stimulus intensity as painful when control groups did. It is now thought that this deficit was likely due to amygdala resection [16, 17], illustrating the importance of the amygdala in pain processing in the brain. Importantly, this deficit in pain perception occurred despite an intact nociceptive system and was not accompanied by the tissue injury characteristic of pain insensitivity disorders, indicating that protective pain functions were intact.
Since the initial discovery of nociceptive pathways to the amygdala, preclinical [5, 7, 8] and clinical [10, 11, 18, 19] studies have provided direct support for amygdala involvement in pain. Electrophysiological recordings in anesthetized rats in vivo and in rodent brain slices in vitro and molecular biological assays showed increased activity markers in response to acute noxious stimuli, including mechanical or thermal stimulation [20, 21], as well as in models of visceral pain [22–28], intraplantar formalin [29–31], acid-induced muscle pain [32], kaolin/carrageenan-induced monoarthritis [33–41], and chronic neuropathic pain [42–44].
The clinical relevance of these findings has been corroborated by human neuroimaging studies that demonstrate amygdala activation in response to experimental noxious stimuli, including mechanical compression, thermal stimulation, and capsaicin application [10], as well as increased amygdala activity in migraineurs compared to healthy controls when presented with negative but not positive or neutral emotional stimuli [45]. In addition, functional connectivity between the left amygdala and the PFC, cingulate cortex, and basal ganglia is different in patients with complex regional pain syndrome (CRPS) [46], and corticolimbic reverberating loops have been implicated in the prediction of and transitioning to chronic pain [11]. Patients with irritable bowel syndrome (IBS) had higher positive resting-state functional connectivity between the amygdala and the insula, pre- and postcentral gyri, and supplementary motor area compared to healthy controls, and this increased connectivity positively correlated to pain intensity [47]. A separate study demonstrated that IBS patients that did not have visceral hypersensitivity had decreased positive resting-state functional connectivity of the amygdala within the default mode network compared to healthy controls as well as IBS patients with visceral hypersensitivity [48]. In female twin pairs with and without chronic pelvic pain, connectivity between the right PAG and the right amygdala, connectivity between the left PAG and the right and left basolateral amygdala, and connectivity of the right basolateral amygdala to the medial orbital frontal cortex, anterior cingulate cortex (ACC), right insula, left thalamus, and hypothalamus differed between the twin with pelvic pain compared to the healthy twin before and after bladder distension by an oral water bolus [49].
2. Amygdala Pain Neurocircuitry
The amygdala receives multiple lines of input (Figure 1) relevant for pain processing, and multiple nuclei in the amygdala are involved in its pain processing functions. These include the lateral-basolateral complex (LA/BLA), the central nucleus (CeA), and the intercalated cell mass (ITC); see Figure 2 and [7–9].
Figure 1.
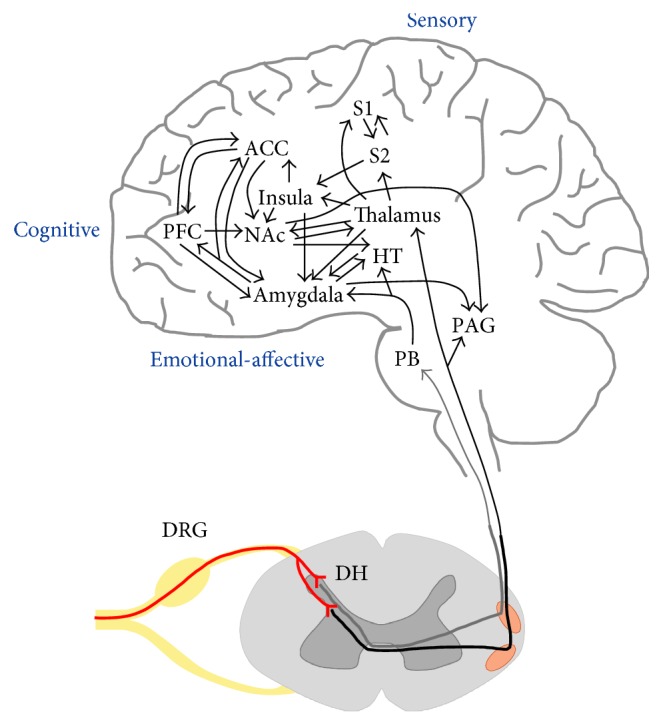
Pain neurocircuitry. Peripheral nociceptive afferent fibers (red lines) form synapses in the dorsal horn of the spinal cord. Axons of spinal dorsal horn neurons decussate in the anterior white commissure and travel in the ventrolateral funiculus (spinothalamic tract; black line) or the dorsolateral funiculus (spino-parabrachio-amygdaloid tract; gray line) to different targets in the brain. Sensory discriminative aspects of pain involve projections from the thalamus to somatosensory cortical areas. Cognitive aspects of pain involve integration within limbic and (prefrontal) cortical regions. Emotional-affective aspects of pain involve integrative processing in the limbic brain regions centered on the amygdala which is a key node. Circuitry is based on [6, 7, 15, 50–58]. Abbreviations: ACC, anterior cingulate cortex; DH, dorsal horn; DRG, dorsal root ganglion; HT, hypothalamus; NAc, nucleus accumbens; PAG, periaqueductal gray; PB, parabrachial nucleus; PFC, prefrontal cortex; S1/2, primary/secondary somatosensory cortex.
Figure 2.
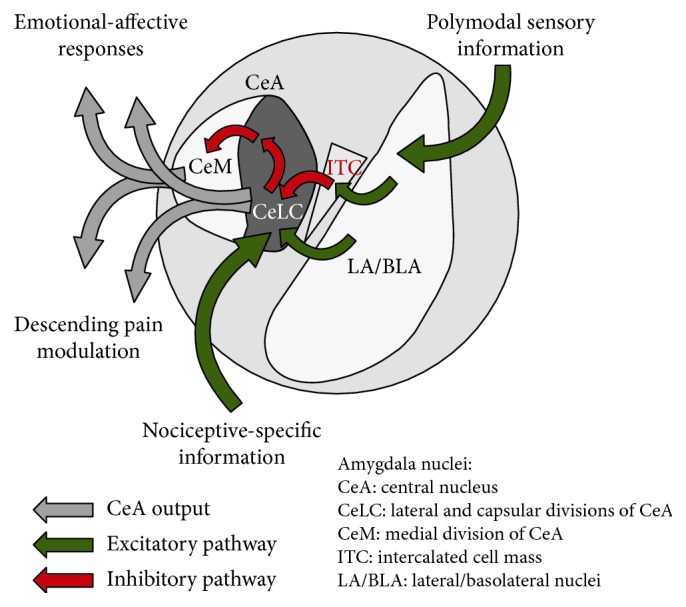
Amygdala pain neurocircuitry. Lateral-basolateral complex (LA/BLA), central nucleus (CeA), and intercalated cell mass (ITC) form the core circuitry involved in amygdala-dependent pain behaviors and pain modulation. The LA/BLA receives polymodal sensory information from cortical and thalamic areas and attaches emotional-affective information, which is then relayed to the CeA. This transmission includes direct excitatory projections to neurons in the lateral and capsular divisions of the CeA (CeLC) as well as feedforward inhibition of CeLC neurons through an LA/BLA projection to the ITC, a group of GABAergic interneurons between LA/BLA and CeA. ITC cells are also the target of cortical control from the mPFC. The CeLC integrates purely nociceptive information received via the spino-parabrachio-amygdaloid tract with highly processed information received from the LA/BLA to generate emotional-affective responses and contribute to top-down pain modulation via projections to the brainstem. This can be done through two types of amygdala outputs: one from CeLC projection neurons and the other from CeM neurons that can be disinhibited by CeLC neurons.
The LA/BLA is predominantly composed of pyramidal glutamatergic projection neurons that receive polymodal sensory, including nociceptive, inputs from the midline and posterior nuclei of the thalamus, insular cortex, and sensory association cortices, as well as inputs from the ACC and medial prefrontal cortex (mPFC) [1, 7–9, 12]. Through associative processing, the LA/BLA attaches emotional-affective content to the sensory inputs and transmits that highly processed information to the amygdala output region in the CeA for further processing as part of amygdala fear and anxiety circuitry [3, 50, 59, 60]. This LA/BLA-CeA projection is now known to generate and modulate pain-related behaviors [7]. The BLA also projects to different cortical areas, including the infra- and prelimbic mPFC, ACC, and perirhinal and insular cortices [51, 61–67]. The BLA-mPFC projection is thought to provide emotional information for value-based executive functions [67–70] and has been implicated in pain-related cortical deactivation and cognitive control [6, 7, 11, 71].
The CeA serves as the major output nucleus for amygdala-driven pain-related functions. The laterocapsular region of the CeA (CeLC, including both the lateral and capsular divisions) receives purely nociceptive information via the spino-parabrachio-amygdaloid tract [72] and possibly via direct projections from the spinal cord [73, 74]. The parabrachial input is characterized by its peptidergic nature and serves as the exclusive source of calcitonin gene-related peptide (CGRP) for the amygdala [75]. The CeLC contains GABAergic projection neurons that also contain peptides such as corticotropin-releasing factor (CRF); they are characterized by their nonaccommodating spike firing pattern [7, 38, 76]. Nearly half of the CRF-CeA neurons receive CGRP input from the PB [77]. This nociceptive information is integrated with polymodal sensory information from the LA/BLA to generate amygdala-mediated responses important for pain behaviors and pain modulation [1, 7–9, 12].
Interposed between LA/BLA and CeLC is a group of GABAergic interneurons in the intercalated cell mass (ITC). ITC cells receive excitatory inputs from the infralimbic mPFC and LA/BLA and are activated during extinction of negative emotional responses [2, 3, 50, 78–82]. Therefore, ITC cells serve a gating function for amygdala output from the CeA through feedforward inhibition that involves activation of neuropeptide S (NPS) receptors on ITC cells [39, 83, 84].
CeA output arises from medial CeA (CeM) and CRF-CeLC projection neurons. These projections target other limbic, hypothalamic, and brainstem regions including the periaqueductal gray (PAG) [3, 60, 85–87]. Projections to hypothalamus and brainstem regions such as the nucleus of the solitary tract (NTS) contribute to amygdala-driven autonomic responses. Projections to the PAG generate or modulate vocalizations and startle responses. CeA projections to other limbic structures such as the bed nucleus of the stria terminalis (BNST) and to monoaminergic regions such as locus coeruleus are thought to drive amygdala influences over anxiety and depression [3, 88, 89]. CeLC neurons can also affect output from the CeM through intra-amygdala connections involving GABAergic protein kinase C delta (PKCδ) positive and other interneurons in the lateral CeA [50, 90–92], but details of this intra-amygdalar circuitry remain to be determined.
CeA neurons expressing PKCδ (PKCδ+) are inhibited by exposure to the fear-conditioned stimulus and have been termed “OFF cells,”, whereas PKCδ-negative neurons are excited when presented with the conditioned stimulus, and reciprocal inhibitory connections exist between these cell types [1, 90, 91]. Inhibition of PKCδ+ cells is believed to contribute to emotional responses such as freezing through disinhibition of CeM output neurons [50]. Importantly, PKCδ+ neurons are distinct from CRF-containing CeLC projection neurons characterized by their nonaccommodating spike firing pattern [87, 91]. And so, it is reasonable to assume that it is the population of PKCδ-negative CRF-CeLC neurons that have been studied for their nociceptive PB input and participation in pain processing [7]. CRF-CeLC neurons are uniquely positioned to provide and regulate amygdala output through their projections to extra-amygdalar targets and through local inhibition of PKCδ+ cells to disinhibit CeM output neurons [1, 3, 60, 85–87] (Figure 2).
3. Pain-Related Changes in Amygdala Neurocircuitry
Importantly, changes in the amygdala neurocircuitry have been detected in different preclinical models of pain and linked mechanistically to pain behaviors, indicating that these maladaptive neuroplastic changes are a brain mechanism of pain. In systems (whole animal) electrophysiology studies of amygdala neurons in anesthetized rats, background activity and responses to mechanical compression of peripheral tissues increased in an arthritis pain model in the CeLC [36, 37, 93–97] and BLA [71], as well as in the CeLC in a neuropathic pain model [42, 44]. These activity changes are not simply a reflection of changes along the pain pathways to the amygdala but arise from synaptic plasticity within the amygdala network. Brain slice physiology studies showed enhanced excitatory transmission at the PB-CeLC and BLA-CeLC synapses [33–35, 38, 40], decreased ITC-mediated synaptic inhibition of CeLC neurons [39, 40], and increased neuronal excitability in the CeLC [33, 34, 38], as well as enhanced excitatory transmission at the LA-BLA synapse and increased excitability of BLA neurons [71] in an arthritis pain model. Enhanced transmission at the PB-CeLC synapse and neuronal excitability [25] and increased levels of neurochemical activity markers (c-Fos and extracellular signal-regulated kinase, ERK) [22, 23, 28, 98] and CRF [24, 26] were found in the CeA in visceral pain models. Enhanced PB-CeLC and BLA-CeLC neurotransmission and excitability [43, 99] and increased expression of CRF and glucocorticoid receptors [100, 101] in the CeA were also observed in neuropathic pain models. It should be noted that not all of these neuroplastic changes have been described in all pain models. It is possible that amygdala neuroplasticity could differ based on time course (acute versus chronic pain) or etiology (inflammatory versus neuropathic) of pain. The full scope of pain-related amygdala plasticity remains an area of active investigation.
The clinical relevance of these neuroplastic changes in preclinical pain models is supported by neuroimaging studies, indicating that amygdala activity is increased in subjects with previously diagnosed pain conditions, including osteoarthritis, IBS, and fibromyalgia compared to matched controls [10].
Interestingly, this amygdala pain-related plasticity exhibits hemispheric lateralization. Under normal conditions, nociceptive inputs into the CeLC are received by and activate both the right and left amygdala [30, 36]. No apparent difference has been detected in background activity or responses to innocuous and noxious mechanical test stimuli between neurons in the right and left amygdala (CeLC), although neurons in the left amygdala appear to have more restricted receptive fields in peripheral tissues [36]. Exogenous activators also can increase neuronal activity in the right and left CeLC [36] or induce hypersensitivity [102]. In inflammatory [36] and neuropathic [42] pain conditions, neurons in the right but not left amygdala exhibit a sustained increase in background and evoked activity irrespective of the side of injury, and the receptive field size of neurons in the right but not left amygdala is increased in an arthritis pain model [36]. In humans, differences in connectivity between the right and left amygdala have been observed in IBS patients compared to healthy controls [47], IBS patients compared to those without visceral hypersensitivity [48], and twin pairs with one twin with and one without chronic pelvic pain before and after bladder distension [49].
Mechanisms of hemispheric lateralization of amygdala plasticity and function are not yet clear. Protein kinase A (PKA) activation has been implicated in the development of central sensitization [103] and synaptic plasticity [33–35, 104] in the CeLC in an arthritis pain model, and application of a PKA inhibitor into the right but not left amygdala resulted in a reduction in neuronal activity in this pain model [36], indicating that PKA is endogenously activated in the right but not left amygdala in the pain state. ERK is also known to play an important role in amygdala plasticity and amygdala-driven behaviors in inflammatory pain models [30, 104], and activation of the metabotropic glutamate receptor 5 (mGluR5) has been shown to increase amygdala ERK activation and ERK-dependent pain behaviors [102, 105]. Application of a blocker of ERK activation [30, 31] and pharmacological blockade or conditional deletion of mGluR5 [102] prevented formalin-induced hypersensitivity when administered into the right but not left amygdala. Chemical (pituitary adenylate cyclase-activating polypeptide) or optogenetic activation of the right but not left CeA increased nociceptive visceromotor responses in a murine urinary bladder distension model, and optogenetic silencing of the left but not right CeA increased bladder distension-induced visceromotor responses [27]. In a visceral pain model cyclophosphamide-induced cystitis, optogenetic activation of the left but not right CeA inhibited abdominal mechanosensitivity, whereas activation of the right CeA further increased visceromotor responses in this model [27] These findings may suggest that while neuroplasticity in the right CeA drives pain-related behaviors, the left CeA may be linked to antinociception.
Evidence from neuroimaging studies in humans also suggests right hemispheric lateralization of amygdala responses to acute experimental pain stimuli [10], whereas in clinical pain conditions, right hemispheric deactivation [106] and left hemispheric signal increases [10] have been reported. This could be due to a compensatory increase in inhibitory transmission in the left amygdala that is not present in the right amygdala in the chronic pain state. Pain-related hemispheric lateralization may reflect a general principle of lateralized emotional processing. Right hemispheric amygdala activation has been found in response to masked fearful faces [107] and was exaggerated in veterans with posttraumatic stress disorder (PTSD) [108]. Enhanced activity of the right, but not the left, amygdala in men was related to encoding and long-term memory of films judged as arousing negative emotions compared to neutral films [109, 110]. Mechanisms and significance of pain-related amygdala lateralization remain to be determined.
4. Pain-Related Amygdala-Centered Corticolimbic Interactions
Information processing in the amygdala can be regulated by inhibitory gating mechanisms centered on ITC cells and their activation by cortical control systems (Figure 2). The mPFC influences amygdala function through feedforward inhibition of CeLC neurons via excitatory projections to ITC cells as an important mechanism of cognitive modulation of emotions such as a fear [2, 3, 50, 78, 79, 81, 82, 88, 111]. Evidence suggests that mPFC-driven feedforward inhibition of CeLC output neurons is impaired in pain [7, 112]. Electrical stimulation of the external capsule, including infralimbic mPFC inputs into the amygdala, resulted in a non-N-methyl-D-aspartate (non-NMDA) receptor-mediated monosynaptic excitatory synaptic response (EPSC) in dorsomedial ITC cells and non-NMDA receptor-driven synaptic inhibition (IPSC) of CeLC neurons [39]. In brain slices from arthritic rats, the monosynaptic EPSC in ITC cells and the glutamate-driven IPSP in CeLC neurons were reduced [39], suggesting pain-related impairment of mPFC-driven feedforward inhibition of amygdala output.
Decreased infralimbic mPFC activity has been implicated in extinction deficits [113–116]. Accumulating evidence points to mPFC deactivation in pain, which could explain impaired control of amygdala processing [7, 112]. Functional and structural abnormalities in the mPFC have been detected in human pain patients [117, 118] and in preclinical pain models [71, 119–121]. As a consequence, activity of output neurons in the infralimbic and prelimbic mPFC is decreased in acute [71, 122, 123] and chronic pain models [120, 124, 125]. Decreased glutamatergic drive of pyramidal cells [120] and abnormally enhanced glutamatergic activation of parvalbumin-expressing GABAergic interneurons [71, 125, 126] have been implicated in the mPFC deactivation in pain.
Hyperactivity in the BLA plays an important role in pain-related mPFC deactivation [7, 112]. The BLA sends glutamatergic projections to the pre- and infralimbic mPFC [61, 66, 67]. Importantly, while some of these BLA axon terminals make direct contact with pyramidal cells, the majority of synapses on neighboring parvalbumin and somatostatin-positive interneurons form GABAergic connections with pyramidal cells, targeting mainly the somatic and proximal axonal regions [62, 127]. This synaptic arrangement was shown to account for amygdala-driven mPFC deactivation by glutamate-driven feedforward inhibition in an arthritis pain model [71, 126]. Feedforward inhibition involves activation of GABAergic interneurons mediated by non-NMDA receptors and mGluR1 but not mGluR5 [122, 128]. It should be noted that mGluR5 in the mPFC is expressed mostly on postsynaptic elements [129] to exert excitatory effects on pyramidal cells [130–132]. In contrast, GABAergic inputs to mGluR5 expressing mPFC pyramidal cells are regulated by cannabinoid CB1 receptors under normal conditions [131, 133]. In brain slices from arthritic rats, IPSCs evoked by electrical or optogenetic activation of BLA axon terminals in pre- and infralimbic mPFC pyramidal cells were increased; IPSCs could be blocked with non-NMDA glutamate receptor and GABAA receptor antagonists [71, 126]. Systems electrophysiology studies in anesthetized rats showed that pain-related decreases in background and evoked activity of prelimbic mPFC pyramidal-like neurons were reversed by a GABAA receptor antagonist and attenuated by an mGluR1 but not mGluR5 antagonist [122]. The decrease in mPFC pyramidal cell activity was causally linked to increased BLA neuronal activity in the arthritis pain model because restoring BLA activity with a CRF1 antagonist increased background and evoked activity of prelimbic mPFC neurons [71].
5. Pharmacological Strategies Targeting Amygdala Pain Neurocircuitry
Interventions that increase amygdala output, even in the absence of acute injury, elicit pain behaviors [27, 30, 102, 134, 135], whereas those that decrease amygdala activity generally inhibit pain behaviors (see [7] for review). Therefore, controlling amygdala activity is a desirable therapeutic strategy for chronic pain. Interventions that were found to have some beneficial effect in preclinical studies include non-NMDA and NMDA receptor antagonists, mGluR1 and mGluR5 antagonists, agonists for group II mGluR2/3 and group III mGluR, including mGluR8, antagonists for CGRP1 and CRF1 receptors, neuropeptide S activating NPS receptors, and inhibitors of ERK and PKA (reviewed in [7]). Here, we will discuss strategies to control amygdala activity by restoring cortical control as well as interventions targeting the amygdala that have emerged from recent studies.
5.1. Strategies Targeting Pain-Related Corticoamygdala Dysfunction
There is good evidence to suggest that mPFC deactivation in pain results in loss of amygdala control (see the Pain-Related Amygdala-Centered Corticolimbic Interactions section). A CRF1 receptor antagonist (NBI27914) inhibited the pain-related increase in synaptic excitation and background and evoked activity of BLA neurons in arthritic rats and increased the background and evoked activity of mPFC neurons that was decreased in the pain model [71]. This intervention also inhibited increased mechanosensitivity (spinal withdrawal reflexes), averse affective responses (audible and ultrasonic vocalizations), and anxiety-like behaviors (measured in the elevated plus maze) and restored normal decision-making on a rodent gambling task in arthritic rats [71].
Another strategy to restore mPFC output used a group II mGluR antagonist (LY341495) to increase synaptically evoked spiking of mPFC pyramidal cells in brain slices from normal and arthritic rats [136]. Effects of a group II agonist (LY379268) showed that these receptors act on glutamatergic synapses from BLA to inhibit direct excitatory transmission and feedforward inhibition onto pyramidal cells, but their net effect is decreased pyramidal cell output, possibly because the effect on EPSCs preceded that on IPSCs. Facilitatory effects of the antagonist suggest that the system may be tonically active to control pyramidal output.
Activation of mGluR5 was tested because of its location on mPFC pyramidal cells (see the Pain-Related Amygdala-Centered Corticolimbic Interactions section). A positive allosteric modulator (PAM) of mGluR5 (VU0360172) increased synaptically evoked spiking in mPFC pyramidal cells using electrical and optogenetic stimulation of BLA inputs [126, 131]. This facilitatory effect on mPFC output involved inhibition of synaptic inhibition by engaging endocannabinoid signaling because CB1 antagonists (AM251 and AM281) and an intracellular inhibitor of diacylglycerol lipase DAGL (tetrahydrolipstatin, THL) blocked the effect of VU0360172 [126, 131]. While this strategy worked under normal conditions, the facilitatory effect of VU0360172 was lost in the arthritis pain model due to a breakdown of mGluR5-driven endocannabinoid signaling in the mPFC resulting in a lack of 2-arachidonoylglycerol (2-AG) [126]. The facilitatory effect of mGluR5 activation on mPFC output was restored with inhibitors of the postsynaptic 2-AG hydrolyzing enzyme ABHD6 (intracellular WWL70) and the monoacylglycerol lipase MGL (JZL184) to increase availability of 2-AG in the postsynaptic cell or with a GABAA receptor blocker (intracellular picrotoxin) [126]. Coapplication of a CB1 receptor agonist (ACEA) with the mGluR5 PAM also increased synaptically evoked spiking of mPFC pyramidal cell neurons in brain slices from arthritic rats by decreasing abnormally enhanced feedforward inhibition from the BLA through depolarization-induced suppression of synaptic inhibition [126]. Systems electrophysiology studies in anesthetized rats with arthritis showed that coadministration of VU0360172 and ACEA into the mPFC increased background and evoked activity of pyramidal-like cells in the mPFC and inhibited the pain-related increase of background and evoked activity in amygdala (CeLC) neurons [123]. This combination strategy also inhibited increased mechanosensitivity (spinal withdrawal reflexes) and audible and ultrasonic vocalizations and mitigated cognitive deficits in the reward-based decision-making in a rodent gambling task in the arthritis pain model [126]. The data further confirm the inverse link between mPFC and amygdala activity and that restoring mPFC output with a combination strategy of mGluR5-CB1 activation can engage cortical control of abnormally enhanced amygdala output to inhibit pain behaviors.
Neuropeptide S (NPS) binds to the Gq/Gs-coupled NPS receptor (NPSR), which is expressed in several brain regions including the dorsomedial ITC cell cluster in the amygdala, and produces anxiolytic effects [84, 137–141]. NPS increased mPFC-driven feedforward inhibition of CeLC neurons by activating ITC cell drive and output in brain slices from arthritic rats through a PKA-dependent mechanism [39]. Intra-ITC as well as nasal application of NPS resulted in decreased background and evoked activity of CeLC neurons in anesthetized rats with arthritis pain, and this effect was blocked by stereotaxic administration of an NPSR antagonist ([D-Cys(tBu)5]NPS or SHA68) into the ITC area [142]. Intra-ITC or nasal application of NPS also inhibited pain-related increases in audible and ultrasonic vocalizations as well as anxiety-like behaviors on the elevated plus maze but had no effect on mechanosensitivity; the inhibitory effects were blocked by stereotaxic administration of [D-Cys(tBu)5]NPS or SHA68 [39, 142].
These studies provide strong evidence for the concept that engaging mPFC control of amygdala processing may be a useful therapeutic strategy for pain management. This concept is supported by studies in humans that have implicated corticolimbic loops, including mPFC-amygdala interactions, in the prediction of and transitioning to chronic pain [11, 143].
5.2. Therapeutic Strategies Targeting Pain-Related Amygdala Hyperactivity
Pharmacological interventions targeting glutamate receptors and neuropeptide systems in the amygdala have been reviewed recently [7]. Here, additional strategies involving the serotonergic system and potassium channels will be discussed.
Serotonergic descending pathways are involved in endogenous antinociceptive signaling from the brain to the spinal cord [52, 144, 145]. However, serotonin (5-HT) actions can be excitatory or inhibitory depending on the specific receptor subtype and its associated neurotransmitter [52]. It is therefore not surprising that selective serotonin reuptake inhibitors (SSRIs) have shown inconsistent efficacy for neuropathic pain treatment [146–148]. One of the at least 14 5-HT receptors, the Gq/11-coupled 5-HT2C receptor, has been implicated in adverse and inconsistent effects of SSRIs for neuropathic pain [149, 150] and, specifically in the BLA, in the generation of anxiogenic behaviors [151–153].
In a rat model of neuropathic pain, viral vector-mediated 5-HT2Creceptor knockdown in the BLA inhibited mechanical hypersensitivity, aversive affective pain behaviors (vocalizations), anxiety-like behaviors, and depression-like behaviors [44]. Pharmacological blockade of 5-HT2C receptors (SB242084) in the BLA conveyed efficacy to a systemically applied SSRI (fluvoxamine) for inhibition of emotional responses (vocalizations) and anxiety-like pain behaviors but not mechanical hypersensitivity [154]. The beneficial behavioral effects of 5-HT2C receptor knockdown in the BLA involved inhibition of irregular and burst firing and evoked activity of CeLC neurons in neuropathic rats [44]. At the synaptic level, 5-HT2C receptor knockdown in the BLA blocked the increase in excitatory transmission at the BLA-CeLC synapse in brain slices from neuropathic rats but had similar inhibitory effects on feedforward inhibition under control conditions and in the neuropathic pain model. 5-HT2C receptor is predominantly expressed in GABAergic neurons, but increased expression in non-GABAergic BLA cells was detected in the neuropathic pain state. The underlying mechanisms of this switch remain to be determined.
Another recent strategy to mitigate pain-related amygdala hyperactivity is activation of small-conductance calcium-activated potassium (SK) channels in the CeA. SK channels are calcium-sensitive, voltage-insensitive potassium channels that are expressed in somatic and dendritic regions of the neuron in a brain region-specific manner [155–158]. Somatically expressed SK channels regulate neuronal excitability by mediating the medium afterhyperpolarization (mAHP) to decrease action potential firing rate [155]. In the amygdala, SK channels regulate action potential firing of neurons in the lateral CeA [159] but not LA [160]. SK channels also regulate dendritic excitability to modulate synaptic transmission and plasticity. In the amygdala, activation of synaptic SK channels in the LA acts as a postsynaptic shunt to reduce excitatory synaptic transmission [161], whereas removal of SK channels from the postsynaptic membrane of LA neurons by a PKA-dependent mechanism facilitates excitatory transmission and synaptic plasticity [162].
A clinically available compound that can inhibit SK channels is riluzole, an FDA approved drug for the treatment of amyotrophic lateral sclerosis (ALS) that easily crosses the blood-brain barrier [163, 164]. It should be noted that other actions of riluzole include inhibition of voltage-gated calcium channels, rapidly inactivating voltage-gated and persistent sodium channels, and glutamate receptor currents [165–167]. Systemically applied riluzole had antinociceptive effects in the formalin test [168–170], in the carrageenan model of hindpaw inflammation [171], and in neuropathic pain models [172–175]. Riluzole also produced pain relief in patients with irritable bowel syndrome [176]. The site and mechanism of pain-related riluzole effects were not identified in these studies. Systemic application of riluzole inhibited emotional responses (audible and ultrasonic vocalizations), but not mechanosensitivity (spinal withdrawal reflexes), in a rodent model of arthritic pain, and these inhibitory effects were reversed by stereotaxic (intra-CeA) administration of a blocker of SK channels (apamin) but not of large-conductance calcium-activated potassium BK channels (charybdotoxin) [177].
An interesting observation is that not every intervention targeting the amygdala to inhibit emotional-affective responses to pain affects mechanosensitivity. This is true for riluzole [177] as well as for NPS [142], an mGluR5 antagonist [178], and an SSRI [154], and may suggest differential roles of neurochemically distinct intra-amygdala circuits.
8. Conclusions
The amygdala is a key node in the interaction of emotional-affective factors with sensory and cognitive aspects of pain. The synaptic and cellular analysis of amygdala function and plasticity as the neurobiological basis of certain pain behaviors has provided a model system for the study of brain mechanisms of pain. The better understanding of relevant intra- and extra-amygdalar circuits and their neurochemical and molecular signatures should yield novel targets for therapeutic interventions because amygdala activity is causally linked to pain behaviors, and therefore, controlling abnormally enhanced amygdala activity is a desirable goal for pain management.
Acknowledgments
Work in the authors' laboratory is supported by National Institutes of Health (NIH) Grant nos. NS081121 and NS038261.
Abbreviations
- 2-AG:
2-arachidonoylglycerol
- 5-HT:
Serotonin
- ACC:
Anterior cingulate cortex
- CeA:
Central nucleus of the amygdala
- CeLC:
Laterocapsular division of the CeA
- CeM:
Medial division of the CeA
- CGRP:
Calcitonin gene-related peptide
- CRF:
Corticotropin-releasing factor
- EPSC:
Excitatory postsynaptic current
- ERK:
Extracellular signal-regulated kinase
- HT:
Hypothalamus
- IBS:
Irritable bowel syndrome
- IPSP/C:
Inhibitory postsynaptic potential/current
- ITC:
Intercalated cell mass of the amygdala
- LA/BLA:
Lateral/basolateral amygdala nuclei
- mGluR:
Metabotropic glutamate receptor
- mPFC:
Medial prefrontal cortex
- NMDA:
N-methyl-D-aspartate
- NPS:
Neuropeptide S
- NPSR:
NPS receptor
- PAG:
Periaqueductal gray
- PAM:
Positive allosteric modulator
- PB:
Parabrachial nucleus
- PKA:
Protein kinase A
- PKCδ:
Protein kinase C delta
- SK:
Small-conductance calcium-activated potassium channel
- SSRI:
Selective serotonin reuptake inhibitor.
Conflicts of Interest
There are no conflicts of interest.
References
- 1.Janak P. H., Tye K. M. From circuits to behaviour in the amygdala. Nature. 2015;517(7534):284–292. doi: 10.1038/nature14188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Marek R., Strobel C., Bredy T. W., Sah P. The amygdala and medial prefrontal cortex: partners in the fear circuit. Journal of Physiology. 2013;591(10):2381–2391. doi: 10.1113/jphysiol.2012.248575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pape H. C., Pare D. Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear. Physiological Reviews. 2010;90(2):419–463. doi: 10.1152/physrev.00037.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Phelps E. A., LeDoux J. E. Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron. 2005;48(2):175–187. doi: 10.1016/j.neuron.2005.09.025. [DOI] [PubMed] [Google Scholar]
- 5.Apkarian A. V., Neugebauer V., Koob G., et al. Neural mechanisms of pain and alcohol dependence. Pharmacology Biochemistry and Behavior. 2013;112:34–41. doi: 10.1016/j.pbb.2013.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bushnell M. C., Ceko M., Low L. A. Cognitive and emotional control of pain and its disruption in chronic pain. Nature Reviews Neuroscience. 2013;14(7):502–511. doi: 10.1038/nrn3516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Neugebauer V. Amygdala pain mechanisms. Handbook of Experimental Pharmacology. 2015;227:261–284. doi: 10.1007/978-3-662-46450-2_13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Neugebauer V., Galhardo V., Maione S., Mackey S. C. Forebrain pain mechanisms. Brain Research Reviews. 2009;60(1):226–242. doi: 10.1016/j.brainresrev.2008.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Neugebauer V., Li W., Bird G. C., Han J. S. The amygdala and persistent pain. Neuroscientist. 2004;10(3):221–234. doi: 10.1177/1073858403261077. [DOI] [PubMed] [Google Scholar]
- 10.Simons L. E., Moulton E. A., Linnman C., Carpino E., Becerra L., Borsook D. The human amygdala and pain: evidence from neuroimaging. Human Brain Mapping. 2014;35(2):527–538. doi: 10.1002/hbm.22199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Vachon-Presseau E., Centeno M. V., Ren W., et al. The emotional brain as a predictor and amplifier of chronic pain. Journal of Dental Research. 2016;95(6):605–612. doi: 10.1177/0022034516638027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Veinante P., Yalcin I., Barrot M. The amygdala between sensation and affect: a role in pain. Journal of Molecular Psychiatry. 2013;1(1):p. 9. doi: 10.1186/2049-9256-1-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bernard J. F., Alden M., Besson J.-M. The organization of the efferent projections from the pontine parabrachial area to the amygdaloid complex: a Phaseolus vulgaris leucoagglutinin (PHA-L) study in the rat. Journal of Comparative Neurology. 1993;329(2):201–229. doi: 10.1002/cne.903290205. [DOI] [PubMed] [Google Scholar]
- 14.Gauriau C., Bernard J.-F. Pain pathways and parabrachial circuits in the rat. Experimental Physiology. 2002;87(2):251–258. doi: 10.1113/eph8702357. [DOI] [PubMed] [Google Scholar]
- 15.Baliki M. N., Apkarian A. V. Nociception, pain, negative moods, and behavior selection. Neuron. 2015;87(3):474–491. doi: 10.1016/j.neuron.2015.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hebben N., Corkin S., Eichenbaum H., Shedlack K. Diminished ability to interpret and report internal states after bilateral medial temporal resection: case H.M. Behavioral Neuroscience. 1985;99(6):1031–1039. doi: 10.1037//0735-7044.99.6.1031. [DOI] [PubMed] [Google Scholar]
- 17.Dossani R. H., Missios S., Nanda A. The legacy of Henry Molaison (1926–2008) and the impact of his bilateral mesial temporal lobe surgery on the study of human memory. World Neurosurgery. 2015;84(4):1127–1135. doi: 10.1016/j.wneu.2015.04.031. [DOI] [PubMed] [Google Scholar]
- 18.Kulkarni B., Bentley D. E., Elliott R., et al. Arthritic pain is processed in brain areas concerned with emotions and fear. Arthritis and Rheumatism. 2007;56(4):1345–1354. doi: 10.1002/art.22460. [DOI] [PubMed] [Google Scholar]
- 19.Tillisch K., Mayer E. A., Labus J. S. Quantitative meta-analysis identifies brain regions activated during rectal distension in irritable bowel syndrome. Gastroenterology. 2011;140(1):91–100. doi: 10.1053/j.gastro.2010.07.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bernard J. F., Huang G. F., Besson J. M. Nucleus centralis of the amygdala and the globus pallidus ventralis: electrophysiological evidence for an involvement in pain processes. Journal of Neurophysiology. 1992;68(2):551–569. doi: 10.1152/jn.1992.68.2.551. [DOI] [PubMed] [Google Scholar]
- 21.Neugebauer V., Li W. Processing of nociceptive mechanical and thermal information in central amygdala neurons with knee-joint input. Journal of Neurophysiology. 2002;87(1):103–112. doi: 10.1152/jn.00264.2001. [DOI] [PubMed] [Google Scholar]
- 22.Bon K., Lanteri-Minet M., Michiels J. F., Menetrey D. Cyclophosphamide cystitis as a model of visceral pain in rats: a c-fos and Krox-24 study at telencephalic levels, with a note on pituitary adenylate cyclase activating polypeptide (PACAP) Experimental Brain Research. 1998;122(2):165–174. doi: 10.1007/s002210050504. [DOI] [PubMed] [Google Scholar]
- 23.Crock L. W., Kolber B. J., Morgan C. D., et al. Central amygdala metabotropic glutamate receptor 5 in the modulation of visceral pain. Journal of Neuroscience. 2012;32(41):14217–14226. doi: 10.1523/jneurosci.1473-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Greenwood-Van Meerveld B., Johnson A. C., Schulkin J., Myers D. A. Long-term expression of corticotropin-releasing factor (CRF) in the paraventricular nucleus of the hypothalamus in response to an acute colonic inflammation. Brain Research. 2006;1071(1):91–96. doi: 10.1016/j.brainres.2005.11.071. [DOI] [PubMed] [Google Scholar]
- 25.Han J. S., Neugebauer V. Synaptic plasticity in the amygdala in a visceral pain model in rats. Neuroscience Letters. 2004;361(1–3):254–257. doi: 10.1016/j.neulet.2003.12.027. [DOI] [PubMed] [Google Scholar]
- 26.Nishii H., Nomura M., Aono H., Fujimoto N., Matsumoto T. Up-regulation of galanin and corticotropin-releasing hormone mRNAs in the key hypothalamic and amygdaloid nuclei in a mouse model of visceral pain. Regulatory Peptides. 2007;141(1–3):105–112. doi: 10.1016/j.regpep.2006.12.022. [DOI] [PubMed] [Google Scholar]
- 27.Sadler K. E., McQuaid N. A., Cox A. C., Behun M. N., Trouten A. M., Kolber B. J. Divergent functions of the left and right central amygdala in visceral nociception. Pain. 2017;158(4):747–759. doi: 10.1097/j.pain.0000000000000830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Suwanprathes P., Ngu M., Ing A., Hunt G., Seow F. c-Fos immunoreactivity in the brain after esophageal acid stimulation. American Journal of Medicine. 2003;115(3):31S–38S. doi: 10.1016/s0002-9343(03)00190-6. [DOI] [PubMed] [Google Scholar]
- 29.Adedoyin M. O., Vicini S., Neale J. H. Endogenous N-acetylaspartylglutamate (NAAG) inhibits synaptic plasticity/transmission in the amygdala in a mouse inflammatory pain model. Molecular Pain. 2010;6:p. 60. doi: 10.1186/1744-8069-6-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Carrasquillo Y., Gereau R. W. Activation of the extracellular signal-regulated kinase in the amygdala modulates pain perception. Journal of Neuroscience. 2007;27(7):1543–1551. doi: 10.1523/jneurosci.3536-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Carrasquillo Y., Gereau R. W. Hemispheric lateralization of a molecular signal for pain modulation in the amygdala. Molecular Pain. 2008;4:p. 24. doi: 10.1186/1744-8069-4-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cheng S. J., Chen C.-C., Yang H.-W., et al. Role of extracellular signal-regulated kinase in synaptic transmission and plasticity of a nociceptive input on capsular central amygdaloid neurons in normal and acid-induced muscle pain mice. Journal of Neuroscience. 2011;31(6):2258–2270. doi: 10.1523/jneurosci.5564-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bird G. C., Lash L. L., Han J. S., Zou X., Willis W. D., Neugebauer V. Protein kinase A-dependent enhanced NMDA receptor function in pain-related synaptic plasticity in rat amygdala neurones. Journal of Physiology. 2005;564(3):907–921. doi: 10.1113/jphysiol.2005.084780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fu Y., Neugebauer V. Differential mechanisms of CRF1 and CRF2 receptor functions in the amygdala in pain-related synaptic facilitation and behavior. Journal of Neuroscience. 2008;28(15):3861–3876. doi: 10.1523/jneurosci.0227-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Han J. S., Li W., Neugebauer V. Critical role of calcitonin gene-related peptide 1 receptors in the amygdala in synaptic plasticity and pain behavior. Journal of Neuroscience. 2005;25(46):10717–10728. doi: 10.1523/jneurosci.4112-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ji G., Neugebauer V. Hemispheric lateralization of pain processing by amygdala neurons. Journal of Neurophysiology. 2009;102(4):2253–2264. doi: 10.1152/jn.00166.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Neugebauer V., Li W. Differential sensitization of amygdala neurons to afferent inputs in a model of arthritic pain. Journal of Neurophysiology. 2003;89(2):716–727. doi: 10.1152/jn.00799.2002. [DOI] [PubMed] [Google Scholar]
- 38.Neugebauer V., Li W., Bird G. C., Bhave G., Gereau R. W. Synaptic plasticity in the amygdala in a model of arthritic pain: differential roles of metabotropic glutamate receptors 1 and 5. Journal of neuroscience. 2003;23(1):52–63. doi: 10.1523/JNEUROSCI.23-01-00052.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ren W., Kiritoshi T., Gregoire S., et al. Neuropeptide S: a novel regulator of pain-related amygdala plasticity and behaviors. Journal of Neurophysiology. 2013;110(8):1765–1781. doi: 10.1152/jn.00874.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ren W., Neugebauer V. Pain-related increase of excitatory transmission and decrease of inhibitory transmission in the central nucleus of the amygdala are mediated by mGluR1. Molecular Pain. 2010;6:p. 93. doi: 10.1186/1744-8069-6-93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tappe-Theodor A., Fu Y., Kuner R., Neugebauer V. Homer1a signaling in the amygdala counteracts pain-related synaptic plasticity, mGluR1 function and pain behaviors. Molecular Pain. 2011;7:p. 38. doi: 10.1186/1744-8069-7-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Goncalves L., Dickenson A. H. Asymmetric time-dependent activation of right central amygdala neurones in rats with peripheral neuropathy and pregabalin modulation. European Journal of Neuroscience. 2012;36(9):3204–3213. doi: 10.1111/j.1460-9568.2012.08235.x. [DOI] [PubMed] [Google Scholar]
- 43.Ikeda R., Takahashi Y., Inoue K., Kato F. NMDA receptor-independent synaptic plasticity in the central amygdala in the rat model of neuropathic pain. Pain. 2007;127(1):161–172. doi: 10.1016/j.pain.2006.09.003. [DOI] [PubMed] [Google Scholar]
- 44.Ji G., Zhang W., Mahimainathan L., et al. 5-HT2C receptor knockdown in the amygdala inhibits neuropathic-pain-related plasticity and behaviors. Journal of Neuroscience. 2017;37(6):1378–1393. doi: 10.1523/jneurosci.2468-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wilcox S. L., Veggeberg R., Lemme J., et al. Increased functional activation of limbic brain regions during negative emotional processing in migraine. Frontiers in Human Neuroscience. 2016;10:p. 366. doi: 10.3389/fnhum.2016.00366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Simons L. E., Elman I., Borsook D. Psychological processing in chronic pain: a neural systems approach. Neuroscience and Biobehavioral Reviews. 2014;39:61–78. doi: 10.1016/j.neubiorev.2013.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Qi R., Liu C., Ke J., et al. Abnormal amygdala resting-state functional connectivity in irritable bowel syndrome. American Journal of Neuroradiology. 2016;37(6):1139–1145. doi: 10.3174/ajnr.a4655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Icenhour A., Witt S. T., Elsenbruch S., et al. Brain functional connectivity is associated with visceral sensitivity in women with irritable bowel syndrome. NeuroImage: Clinical. 2017;15:449–457. doi: 10.1016/j.nicl.2017.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kleinhans N. M., Yang C. C., Strachan E. D., Buchwald D. S., Maravilla K. R. Alterations in connectivity on functional magnetic resonance imaging with provocation of lower urinary tract symptoms: a MAPP research network feasibility study of urological chronic pelvic pain syndromes. Journal of Urology. 2016;195(3):639–645. doi: 10.1016/j.juro.2015.09.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Duvarci S., Pare D. Amygdala microcircuits controlling learned fear. Neuron. 2014;82(5):966–980. doi: 10.1016/j.neuron.2014.04.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sripanidkulchai K., Sripanidkulchai B., Wyss J. M. The cortical projection of the basolateral amygdaloid nucleus in the rat: a retrograde fluorescent dye study. Journal of Comparative Neurology. 1984;229(3):419–431. doi: 10.1002/cne.902290310. [DOI] [PubMed] [Google Scholar]
- 52.Ossipov M. H., Morimura K., Porreca F. Descending pain modulation and chronification of pain. Current Opinion in Supportive and Palliative Care. 2014;8(2):143–151. doi: 10.1097/SPC.0000000000000055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Almeida T. F., Roizenblatt S., Tufik S. Afferent pain pathways: a neuroanatomical review. Brain Research. 2004;1000(1-2):40–56. doi: 10.1016/j.brainres.2003.10.073. [DOI] [PubMed] [Google Scholar]
- 54.Price D. D. Psychological and neural mechanisms of the affective dimension of pain. Science. 2000;288(5472):1769–1772. doi: 10.1126/science.288.5472.1769. [DOI] [PubMed] [Google Scholar]
- 55.Russchen F. T., Price J. L. Amygdalostriatal projections in the rat. Topographical organization and fiber morphology shown using the lectin PHA-L as an anterograde tracer. Neuroscience Letters. 1984;47(1):15–22. doi: 10.1016/0304-3940(84)90379-3. [DOI] [PubMed] [Google Scholar]
- 56.Wakefield C., Hall E. Hypothalamic projections to the amygdala in the cat. A light and electron microscopic study. Cell and Tissue Research. 1974;151(4):499–508. doi: 10.1007/bf00222995. [DOI] [PubMed] [Google Scholar]
- 57.Williams D. J., Crossman A. R., Slater P. The efferent projections of the nucleus accumbens in the rat. Brain Research. 1977;130(2):217–227. doi: 10.1016/0006-8993(77)90271-2. [DOI] [PubMed] [Google Scholar]
- 58.Zahm D. S., Heimer L. Specificity in the efferent projections of the nucleus accumbens in the rat: comparison of the rostral pole projection patterns with those of the core and shell. Journal of Comparative Neurology. 1993;327(2):220–232. doi: 10.1002/cne.903270205. [DOI] [PubMed] [Google Scholar]
- 59.Pare D., Duvarci S. Amygdala microcircuits mediating fear expression and extinction. Current Opinion in Neurobiology. 2012;22(4):717–723. doi: 10.1016/j.conb.2012.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Sah P., Faber E. S. L., Lopez De Armentia M., Power J. The amygdaloid complex: anatomy and physiology. Physiological Reviews. 2003;83(3):803–834. doi: 10.1152/physrev.00002.2003. [DOI] [PubMed] [Google Scholar]
- 61.Cheriyan J., Kaushik M. K., Ferreira A. N., Sheets P. L. Specific targeting of the basolateral amygdala to projectionally defined pyramidal neurons in prelimbic and infralimbic cortex. eNeuro. 2016;3(2) doi: 10.1523/eneuro.0002-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Gabbott P. L. A., Warner T. A., Busby S. J. Amygdala input monosynaptically innervates parvalbumin immunoreactive local circuit neurons in rat medial prefrontal cortex. Neuroscience. 2006;139(3):1039–1048. doi: 10.1016/j.neuroscience.2006.01.026. [DOI] [PubMed] [Google Scholar]
- 63.Krettek J. E., Price J. L. Projections from the amygdaloid complex and adjacent olfactory structures to the entorhinal cortex and to the subiculum in the rat and cat. Journal of Comparative Neurology. 1977;172(4):723–752. doi: 10.1002/cne.901720409. [DOI] [PubMed] [Google Scholar]
- 64.Krettek J. E., Price J. L. Projections from the amygdaloid complex to the cerebral cortex and thalamus in the rat and cat. Journal of Comparative Neurology. 1977;172(4):687–722. doi: 10.1002/cne.901720408. [DOI] [PubMed] [Google Scholar]
- 65.Likhtik E., Pelletier J. G., Popescu A. T., Pare D. Identification of basolateral amygdala projection cells and interneurons using extracellular recordings. Journal of Neurophysiology. 2006;96(6):3257–3265. doi: 10.1152/jn.00577.2006. [DOI] [PubMed] [Google Scholar]
- 66.Little J. P., Carter A. G. Synaptic mechanisms underlying strong reciprocal connectivity between the medial prefrontal cortex and basolateral amygdala. Journal of Neuroscience. 2013;33(39):15333–15342. doi: 10.1523/jneurosci.2385-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Senn V., Wolff S. B. E., Herry C., et al. Long-range connectivity defines behavioral specificity of amygdala neurons. Neuron. 2014;81(2):428–437. doi: 10.1016/j.neuron.2013.11.006. [DOI] [PubMed] [Google Scholar]
- 68.Holland P. C., Gallagher M. Amygdala-frontal interactions and reward expectancy. Current Opinion in Neurobiology. 2004;14(2):148–155. doi: 10.1016/j.conb.2004.03.007. [DOI] [PubMed] [Google Scholar]
- 69.McGaugh J. L. The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annual Review of Neuroscience. 2004;27(1):1–28. doi: 10.1146/annurev.neuro.27.070203.144157. [DOI] [PubMed] [Google Scholar]
- 70.Tan H., Lauzon N. M., Bishop S. F., Bechard M. A., Laviolette S. R. Integrated cannabinoid CB1 receptor transmission within the amygdala–prefrontal cortical pathway modulates neuronal plasticity and emotional memory encoding. Cerebral Cortex. 2010;20(6):1486–1496. doi: 10.1093/cercor/bhp210. [DOI] [PubMed] [Google Scholar]
- 71.Ji G., Sun H., Fu Y., et al. Cognitive impairment in pain through amygdala-driven prefrontal cortical deactivation. Journal of Neuroscience. 2010;30(15):5451–5464. doi: 10.1523/JNEUROSCI.0225-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bernard J. F., Besson J. M. The spino(trigemino)pontoamygdaloid pathway: electrophysiological evidence for an involvement in pain processes. Journal of Neurophysiology. 1990;63(3):473–490. doi: 10.1152/jn.1990.63.3.473. [DOI] [PubMed] [Google Scholar]
- 73.Burstein R., Potrebic S. Retrograde labeling of neurons in the spinal cord that project directly to the amygdala or the orbital cortex in the rat. Journal of Comparative Neurology. 1993;335(4):469–485. doi: 10.1002/cne.903350402. [DOI] [PubMed] [Google Scholar]
- 74.Wang C.-C., Willis W. D., Westlund K. N. Ascending projections from the area around the spinal cord central canal: a Phaseolus vulgaris leucoagglutinin study in rats. Journal of Comparative Neurology. 1999;415(3):341–367. doi: 10.1002/(sici)1096-9861(19991220)415:3<341::aid-cne3>3.0.co;2-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Dobolyi A., Irwin S., Makara G., Usdin T. B., Palkovits M. Calcitonin gene-related peptide-containing pathways in the rat forebrain. Journal of Comparative Neurology. 2005;489(1):92–119. doi: 10.1002/cne.20618. [DOI] [PubMed] [Google Scholar]
- 76.Schiess M. C., Callahan P. M., Zheng H. Characterization of the electrophysiological and morphological properties of rat central amygdala neurons in vitro. Journal of Neuroscience Research. 1999;58(5):663–673. doi: 10.1002/(sici)1097-4547(19991201)58:5<663::aid-jnr7>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- 77.Harrigan E. A., Magnuson D. J., Thunstedt G. M., Gray T. S. Corticotropin releasing factor neurons are innervated by calcitonin gene-related peptide terminals in the rat central amygdaloid nucleus. Brain Research Bulletin. 1994;33(5):529–534. doi: 10.1016/0361-9230(94)90079-5. [DOI] [PubMed] [Google Scholar]
- 78.Amano T., Unal C. T., Pare D. Synaptic correlates of fear extinction in the amygdala. Nature Neuroscience. 2010;13(4):489–494. doi: 10.1038/nn.2499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Amir A., Amano T., Pare D. Physiological identification and infralimbic responsiveness of rat intercalated amygdala neurons. Journal of Neurophysiology. 2011;105(6):3054–3066. doi: 10.1152/jn.00136.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Pinard C. R., Mascagni F., McDonald A. J. Medial prefrontal cortical innervation of the intercalated nuclear region of the amygdala. Neuroscience. 2012;205:112–124. doi: 10.1016/j.neuroscience.2011.12.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Royer S., Martina M., Pare D. An inhibitory interface gates impulse traffic between the input and output stations of the amygdala. Journal of Neuroscience. 1999;19(23):10575–10583. doi: 10.1523/JNEUROSCI.19-23-10575.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Strobel C., Marek R., Gooch H. M., Sullivan R. K., Sah P. Prefrontal and auditory input to intercalated neurons of the amygdala. Cell Reports. 2015;10(9):1435–1442. doi: 10.1016/j.celrep.2015.02.008. [DOI] [PubMed] [Google Scholar]
- 83.Pape H. C., Jungling K., Seidenbecher T., Lesting J., Reinscheid R. K. Neuropeptide S: a transmitter system in the brain regulating fear and anxiety. Neuropharmacology. 2010;58(1):29–34. doi: 10.1016/j.neuropharm.2009.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Reinscheid R. K. Neuropeptide S: anatomy, pharmacology, genetics and physiological functions. Results and Problems in Cell Differentiation. 2008;46:145–158. doi: 10.1007/400_2007_051. [DOI] [PubMed] [Google Scholar]
- 85.Bourgeais L., Gauriau C., Bernard J. F. Projections from the nociceptive area of the central nucleus of the amygdala to the forebrain: a PHA-L study in the rat. European Journal of Neuroscience. 2001;14(2):229–255. doi: 10.1046/j.0953-816x.2001.01640.x. [DOI] [PubMed] [Google Scholar]
- 86.Jongen-Relo A. L., Amaral D. G. Evidence for a GABAergic projection from the central nucleus of the amygdala to the brainstem of the macaque monkey: a combined retrograde tracing and in situ hybridization study. European Journal of Neuroscience. 1998;10(9):2924–2933. doi: 10.1046/j.1460-9568.1998.00299.x. [DOI] [PubMed] [Google Scholar]
- 87.Pomrenze M. B., Millan E. Z., Hopf F. W., et al. A transgenic rat for investigating the anatomy and function of corticotrophin releasing factor circuits. Frontiers in Neuroscience. 2015;9:p. 487. doi: 10.3389/fnins.2015.00487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ehrlich I., Humeau Y., Grenier F., Ciocchi S., Herry C., Luthi A. Amygdala inhibitory circuits and the control of fear memory. Neuron. 2009;62(6):757–771. doi: 10.1016/j.neuron.2009.05.026. [DOI] [PubMed] [Google Scholar]
- 89.McCall J. G., Al-Hasani R., Siuda E. R., et al. CRH engagement of the locus coeruleus noradrenergic system mediates stress-induced anxiety. Neuron. 2015;87(3):605–620. doi: 10.1016/j.neuron.2015.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ciocchi S., Herry C., Grenier F., et al. Encoding of conditioned fear in central amygdala inhibitory circuits. Nature. 2010;468(7321):277–282. doi: 10.1038/nature09559. [DOI] [PubMed] [Google Scholar]
- 91.Fadok J. P., Krabbe S., Markovic M., et al. A competitive inhibitory circuit for selection of active and passive fear responses. Nature. 2017;542(7639):96–100. doi: 10.1038/nature21047. [DOI] [PubMed] [Google Scholar]
- 92.Haubensak W., Kunwar P. S., Cai H., et al. Genetic dissection of an amygdala microcircuit that gates conditioned fear. Nature. 2010;468(7321):270–276. doi: 10.1038/nature09553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ji G., Horvath C., Neugebauer V. NR2B receptor blockade inhibits pain-related sensitization of amygdala neurons. Molecular Pain. 2009;5:p. 21. doi: 10.1186/1744-8069-5-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ji G., Neugebauer V. Differential effects of CRF1 and CRF2 receptor antagonists on pain-related sensitization of neurons in the central nucleus of the amygdala. Journal of Neurophysiology. 2007;97(6):3893–3904. doi: 10.1152/jn.00135.2007. [DOI] [PubMed] [Google Scholar]
- 95.Li W., Neugebauer V. Differential roles of mGluR1 and mGluR5 in brief and prolonged nociceptive processing in central amygdala neurons. Journal of Neurophysiology. 2004;91(1):13–24. doi: 10.1152/jn.00485.2003. [DOI] [PubMed] [Google Scholar]
- 96.Li W., Neugebauer V. Differential changes of group II and group III mGluR function in central amygdala neurons in a model of arthritic pain. Journal of Neurophysiology. 2006;96(4):1803–1815. doi: 10.1152/jn.00495.2006. [DOI] [PubMed] [Google Scholar]
- 97.Li W., Neugebauer V. Block of NMDA and non-NMDA receptor activation results in reduced background and evoked activity of central amygdala neurons in a model of arthritic pain. Pain. 2004;110(1):112–122. doi: 10.1016/j.pain.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 98.Nakagawa T., Katsuya A., Tanimoto S., et al. Differential patterns of c-fos mRNA expression in the amygdaloid nuclei induced by chemical somatic and visceral noxious stimuli in rats. Neuroscience Letters. 2003;344(3):197–200. doi: 10.1016/s0304-3940(03)00465-8. [DOI] [PubMed] [Google Scholar]
- 99.Nakao A., Takahashi Y., Nagase M., Ikeda R., Kato F. Role of capsaicin-sensitive C-fiber afferents in neuropathic pain-induced synaptic potentiation in the nociceptive amygdala. Molecular Pain. 2012;8:p. 51. doi: 10.1186/1744-8069-8-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Rouwette T., Vanelderen P., de Reus M., et al. Experimental neuropathy increases limbic forebrain CRF. European Journal of Pain. 2012;16(1):61–71. doi: 10.1016/j.ejpain.2011.05.016. [DOI] [PubMed] [Google Scholar]
- 101.Ulrich-Lai Y. M., Xie W., Meij J. T., Dolgas C. M., Yu L., Herman J. P. Limbic and HPA axis function in an animal model of chronic neuropathic pain. Physiology and Behavior. 2006;88(1-2):67–76. doi: 10.1016/j.physbeh.2006.03.012. [DOI] [PubMed] [Google Scholar]
- 102.Kolber B. J., Montana M. C., Carrasquillo Y., et al. Activation of metabotropic glutamate receptor 5 in the amygdala modulates pain-like behavior. Journal of Neuroscience. 2010;30(24):8203–8213. doi: 10.1523/jneurosci.1216-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ji G., Neugebauer V. Pro- and anti-nociceptive effects of corticotropin-releasing factor (CRF) in central amygdala neurons are mediated through different receptors. Journal of Neurophysiology. 2008;99(3):1201–1212. doi: 10.1152/jn.01148.2007. [DOI] [PubMed] [Google Scholar]
- 104.Fu Y., Han J., Ishola T., et al. PKA and ERK, but not PKC, in the amygdala contribute to pain-related synaptic plasticity and behavior. Molecular Pain. 2008;4:p. 26. doi: 10.1186/1744-8069-4-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Li Z., Ji G., Neugebauer V. Mitochondrial reactive oxygen species are activated by mGluR5 through IP3 and activate ERK and PKA to increase excitability of amygdala neurons and pain behavior. Journal of Neuroscience. 2011;31(3):1114–1127. doi: 10.1523/jneurosci.5387-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Bonaz B., Baciu M., Papillon E., et al. Central processing of rectal pain in patients with irritable bowel syndrome: an fMRI study. American Journal of Gastroenterology. 2002;97(3):654–661. doi: 10.1111/j.1572-0241.2002.05545.x. [DOI] [PubMed] [Google Scholar]
- 107.Whalen P. J., Rauch S. L., Etcoff N. L., McInerney S. C., Lee M. B., Jenike M. A. Masked presentations of emotional facial expressions modulate amygdala activity without explicit knowledge. Journal of Neuroscience. 1998;18(1):411–418. doi: 10.1523/JNEUROSCI.18-01-00411.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Rauch S. L., Whalen P. J., Shin L. M., et al. Exaggerated amygdala response to masked facial stimuli in posttraumatic stress disorder: a functional MRI study. Biological Psychiatry. 2000;47(9):769–776. doi: 10.1016/s0006-3223(00)00828-3. [DOI] [PubMed] [Google Scholar]
- 109.Cahill L., Haier R. J., White N. S., et al. Sex-related difference in amygdala activity during emotionally influenced memory storage. Neurobiology of Learning and Memory. 2001;75(1):1–9. doi: 10.1006/nlme.2000.3999. [DOI] [PubMed] [Google Scholar]
- 110.Cahill L., Uncapher M., Kilpatrick L., Alkire M. T., Turner J. Sex-related hemispheric lateralization of amygdala function in emotionally influenced memory: an fMRI investigation. Learning and Memory. 2004;11(3):261–266. doi: 10.1101/lm.70504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Likhtik E., Popa D., Apergis-Schoute J., Fidacaro G. A., Pare D. Amygdala intercalated neurons are required for expression of fear extinction. Nature. 2008;454(7204):642–645. doi: 10.1038/nature07167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Woodhams S. G., Chapman V., Finn D. P., Hohmann A. G., Neugebauer V. The cannabinoid system and pain. Neuropharmacology. 2017;124:105–120. doi: 10.1016/j.neuropharm.2017.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Chang C. H., Maren S. Strain difference in the effect of infralimbic cortex lesions on fear extinction in rats. Behavioral Neuroscience. 2010;124(3):391–397. doi: 10.1037/a0019479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hefner K., Whittle N., Juhasz J., et al. Impaired fear extinction learning and cortico-amygdala circuit abnormalities in a common genetic mouse strain. Journal of Neuroscience. 2008;28(32):8074–8085. doi: 10.1523/jneurosci.4904-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Kim S. C., Jo Y. S., Kim I. H., Kim H., Choi J.-S. Lack of medial prefrontal cortex activation underlies the immediate extinction deficit. Journal of Neuroscience. 2010;30(3):832–837. doi: 10.1523/jneurosci.4145-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Sierra-Mercado D., Padilla-Coreano N., Quirk G. J. Dissociable roles of prelimbic and infralimbic cortices, ventral hippocampus, and basolateral amygdala in the expression and extinction of conditioned fear. Neuropsychopharmacology. 2011;36(2):529–538. doi: 10.1038/npp.2010.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Apkarian A. V., Sosa Y., Sonty S., et al. Chronic back pain is associated with decreased prefrontal and thalamic gray matter density. Journal of Neuroscience. 2004;24(46):10410–10415. doi: 10.1523/jneurosci.2541-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Mayer E. A., Berman S., Suyenobu B., et al. Differences in brain responses to visceral pain between patients with irritable bowel syndrome and ulcerative colitis. Pain. 2005;115(3):398–409. doi: 10.1016/j.pain.2005.03.023. [DOI] [PubMed] [Google Scholar]
- 119.Cardoso-Cruz H., Sousa M., Vieira J. B., Lima D., Galhardo V. Prefrontal cortex and mediodorsal thalamus reduced connectivity is associated with spatial working memory impairment in rats with inflammatory pain. Pain. 2013;154(11):2397–2406. doi: 10.1016/j.pain.2013.07.020. [DOI] [PubMed] [Google Scholar]
- 120.Kelly C. J., Huang M., Meltzer H., Martina M. Reduced glutamatergic currents and dendritic branching of layer 5 pyramidal cells contribute to medial prefrontal cortex deactivation in a rat model of neuropathic pain. Frontiers in Cellular Neuroscience. 2016;10:p. 133. doi: 10.3389/fncel.2016.00133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Metz A. E., Yau H.-J., Centeno M. V., Apkarian A. V., Martina M. Morphological and functional reorganization of rat medial prefrontal cortex in neuropathic pain. Proceedings of the National Academy of Sciences. 2009;106(7):2423–2428. doi: 10.1073/pnas.0809897106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Ji G., Neugebauer V. Pain-related deactivation of medial prefrontal cortical neurons involves mGluR1 and GABAA receptors. Journal of Neurophysiology. 2011;106(5):2642–2652. doi: 10.1152/jn.00461.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Ji G., Neugebauer V. CB1 augments mGluR5 function in medial prefrontal cortical neurons to inhibit amygdala hyperactivity in an arthritis pain model. European Journal of Neuroscience. 2014;39(3):455–466. doi: 10.1111/ejn.12432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Wang G. Q., Cen C., Li C., et al. Deactivation of excitatory neurons in the prelimbic cortex via Cdk5 promotes pain sensation and anxiety. Nature Communications. 2015;6:p. 7660. doi: 10.1038/ncomms8660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Zhang Z., Gadotti V. M., Chen L., Souza I. A., Stemkowski P. L., Zamponi G. W. Role of prelimbic GABAergic circuits in sensory and emotional aspects of neuropathic pain. Cell Reports. 2015;12(5):752–759. doi: 10.1016/j.celrep.2015.07.001. [DOI] [PubMed] [Google Scholar]
- 126.Kiritoshi T., Ji G., Neugebauer V. Rescue of impaired mGluR5-driven endocannabinoid signaling restores prefrontal cortical output to inhibit pain in arthritic rats. Journal of Neuroscience. 2016;36(3):837–850. doi: 10.1523/jneurosci.4047-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.McGarry L. M., Carter A. G. Inhibitory gating of basolateral amygdala inputs to the prefrontal cortex. Journal of Neuroscience. 2016;36(36):9391–9406. doi: 10.1523/jneurosci.0874-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Sun H., Neugebauer V. mGluR1, but not mGluR5, activates feed-forward inhibition in the medial prefrontal cortex to impair decision making. Journal of Neurophysiology. 2011;106(2):960–973. doi: 10.1152/jn.00762.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Muly E. C., Maddox M., Smith Y. Distribution of mGluR1α and mGluR5 immunolabeling in primate prefrontal cortex. Journal of Comparative Neurology. 2003;467(4):521–535. doi: 10.1002/cne.10937. [DOI] [PubMed] [Google Scholar]
- 130.Fontanez-Nuin D. E., Santini E., Quirk G. J., Porter J. T. Memory for fear extinction requires mGluR5-mediated activation of infralimbic neurons. Cerebral Cortex. 2011;21(3):727–735. doi: 10.1093/cercor/bhq147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Kiritoshi T., Sun H., Ren W., et al. Modulation of pyramidal cell output in the medial prefrontal cortex by mGluR5 interacting with CB1. Neuropharmacology. 2013;66:170–178. doi: 10.1016/j.neuropharm.2012.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Marek G. J., Zhang C. Activation of metabotropic glutamate 5 (mGlu5) receptors induces spontaneous excitatory synaptic currents in layer V pyramidal cells of the rat prefrontal cortex. Neuroscience Letters. 2008;442(3):239–243. doi: 10.1016/j.neulet.2008.06.083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Lafourcade M., Elezgarai I., Mato S., Bakiri Y., Grandes P., Manzoni O. J. Molecular components and functions of the endocannabinoid system in mouse prefrontal cortex. PLoS One. 2007;2(8):p. e709. doi: 10.1371/journal.pone.0000709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Han J. S., Adwanikar H., Li Z., Ji G., Neugebauer V. Facilitation of synaptic transmission and pain responses by CGRP in the amygdala of normal rats. Molecular Pain. 2010;6:p. 10. doi: 10.1186/1744-8069-6-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Ji G., Fu Y., Adwanikar H., Neugebauer V. Non-pain-related CRF1 activation in the amygdala facilitates synaptic transmission and pain responses. Molecular Pain. 2013;9:p. 2. doi: 10.1186/1744-8069-9-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Kiritoshi T., Neugebauer V. Group II mGluRs modulate baseline and arthritis pain-related synaptic transmission in the rat medial prefrontal cortex. Neuropharmacology. 2015;95:388–394. doi: 10.1016/j.neuropharm.2015.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Jungling K., Seidenbecher T., Sosulina L., et al. Neuropeptide S-mediated control of fear expression and extinction: role of intercalated GABAergic neurons in the amygdala. Neuron. 2008;59(2):298–310. doi: 10.1016/j.neuron.2008.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Leonard S. K., Ring R. H. Immunohistochemical localization of the neuropeptide S receptor in the rat central nervous system. Neuroscience. 2011;172:153–163. doi: 10.1016/j.neuroscience.2010.10.020. [DOI] [PubMed] [Google Scholar]
- 139.Pulga A., Ruzza C., Rizzi A., Guerrini R., Calo G. Anxiolytic- and panicolytic-like effects of neuropeptide S in the mouse elevated T-maze. European Journal of Neuroscience. 2012;36(11):3531–3537. doi: 10.1111/j.1460-9568.2012.08265.x. [DOI] [PubMed] [Google Scholar]
- 140.Ruzza C., Rizzi A., Camarda V., et al. [tBu-D-Gly5]NPS, a pure and potent antagonist of the neuropeptide S receptor: in vitro and in vivo studies. Peptides. 2012;34(2):404–411. doi: 10.1016/j.peptides.2012.01.024. [DOI] [PubMed] [Google Scholar]
- 141.Xu Y. L., Gall C. M., Jackson V. R., Civelli O., Reinscheid R. K. Distribution of neuropeptide S receptor mRNA and neurochemical characteristics of neuropeptide S-expressing neurons in the rat brain. Journal of Comparative Neurology. 2007;500(1):84–102. doi: 10.1002/cne.21159. [DOI] [PubMed] [Google Scholar]
- 142.Medina G., Ji G., Gregoire S., Neugebauer V. Nasal application of neuropeptide S inhibits arthritis pain-related behaviors through an action in the amygdala. Molecular Pain. 2014;10:p. 32. doi: 10.1186/1744-8069-10-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Vachon-Presseau E., Tetreault P., Petre B., et al. Corticolimbic anatomical characteristics predetermine risk for chronic pain. Brain. 2016;139(7):1958–1970. doi: 10.1093/brain/aww100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Mason P. Contributions of the medullary raphe and ventromedial reticular region to pain modulation and other homeostatic functions. Annual Review of Neuroscience. 2001;24(1):737–777. doi: 10.1146/annurev.neuro.24.1.737. [DOI] [PubMed] [Google Scholar]
- 145.Staud R. The important role of CNS facilitation and inhibition for chronic pain. International Journal of Clinical Rheumatology. 2013;8(6):639–646. doi: 10.2217/ijr.13.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Dworkin R. H., O’Connor A. B., Audette J., et al. Recommendations for the pharmacological management of neuropathic pain: an overview and literature update. Mayo Clinic Proceedings. 2010;85(3):S3–S14. doi: 10.4065/mcp.2009.0649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Finnerup N. B., Attal N., Haroutounian S., et al. Pharmacotherapy for neuropathic pain in adults: a systematic review and meta-analysis. Lancet Neurology. 2015;14(2):162–173. doi: 10.1016/S1474-4422(14)70251-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Lee Y. C., Chen P. P. A review of SSRIs and SNRIs in neuropathic pain. Expert Opinion on Pharmacotherapy. 2010;11(17):2813–2825. doi: 10.1517/14656566.2010.507192. [DOI] [PubMed] [Google Scholar]
- 149.Brasch-Andersen C., Moller M. U., Christiansen L., et al. A candidate gene study of serotonergic pathway genes and pain relief during treatment with escitalopram in patients with neuropathic pain shows significant association to serotonin receptor2C (HTR2C) European Journal of Clinical Pharmacology. 2011;67(11):1131–1137. doi: 10.1007/s00228-011-1056-x. [DOI] [PubMed] [Google Scholar]
- 150.Jensen N. H., Cremers T. I., Sotty F. Therapeutic potential of 5-HT2C receptor ligands. Scientific World Journal. 2010;10:1870–1885. doi: 10.1100/tsw.2010.180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Campbell B. M., Merchant K. M. Serotonin 2C receptors within the basolateral amygdala induce acute fear-like responses in an open-field environment. Brain Research. 2003;993(1-2):1–9. doi: 10.1016/s0006-8993(03)03384-5. [DOI] [PubMed] [Google Scholar]
- 152.Christianson J. P., Ragole T., Amat J., et al. 5-hydroxytryptamine 2C receptors in the basolateral amygdala are involved in the expression of anxiety after uncontrollable traumatic stress. Biological Psychiatry. 2010;67(4):339–345. doi: 10.1016/j.biopsych.2009.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Heisler L. K., Zhou L., Bajwa P., Hsu J., Tecott L. H. Serotonin 5-HT2C receptors regulate anxiety-like behavior. Genes, Brain and Behavior. 2007;6(5):491–496. doi: 10.1111/j.1601-183x.2007.00316.x. [DOI] [PubMed] [Google Scholar]
- 154.Gregoire S., Neugebauer V. 5-HT2CR blockade in the amygdala conveys analgesic efficacy to SSRIs in a rat model of arthritis pain. Molecular Pain. 2013;9:p. 41. doi: 10.1186/1744-8069-9-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Adelman J. P., Maylie J., Sah P. Small-conductance Ca2+-activated K+ channels: form and function. Annual Review of Physiology. 2012;74(1):245–269. doi: 10.1146/annurev-physiol-020911-153336. [DOI] [PubMed] [Google Scholar]
- 156.Gymnopoulos M., Cingolani L. A., Pedarzani P., Stocker M. Developmental mapping of small-conductance calcium-activated potassium channel expression in the rat nervous system. Journal of Comparative Neurology. 2014;522(5):1072–1101. doi: 10.1002/cne.23466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Kohler M., Hirschberg B., Bond C. T., et al. Small-conductance, calcium-activated potassium channels from mammalian brain. Science. 1996;273(5282):1709–1714. doi: 10.1126/science.273.5282.1709. [DOI] [PubMed] [Google Scholar]
- 158.Stocker M. Ca2+-activated K+ channels: molecular determinants and function of the SK family. Nature Reviews Neuroscience. 2004;5(10):758–770. doi: 10.1038/nrn1516. [DOI] [PubMed] [Google Scholar]
- 159.Lopez de Armentia M., Sah P. Firing properties and connectivity of neurons in the rat lateral central nucleus of the amygdala. Journal of Neurophysiology. 2004;92(3):1285–1294. doi: 10.1152/jn.00211.2004. [DOI] [PubMed] [Google Scholar]
- 160.Faber E. S. L., Sah P. Functions of SK channels in central neurons. Clinical and Experimental Pharmacology and Physiology. 2007;34(10):1077–1083. doi: 10.1111/j.1440-1681.2007.04725.x. [DOI] [PubMed] [Google Scholar]
- 161.Faber E. S., Delaney A. J., Sah P. SK channels regulate excitatory synaptic transmission and plasticity in the lateral amygdala. Nature Neuroscience. 2005;8(5):635–641. doi: 10.1038/nn1450. [DOI] [PubMed] [Google Scholar]
- 162.Faber E. S., Delaney A. J., Power J. M., Sedlak P. L., Crane J. W., Sah P. Modulation of SK channel trafficking by beta adrenoceptors enhances excitatory synaptic transmission and plasticity in the amygdala. Journal of Neuroscience. 2008;28(43):10803–10813. doi: 10.1523/jneurosci.1796-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Cao Y. J., Dreixler J. C., Couey J. J., Houamed K. M. Modulation of recombinant and native neuronal SK channels by the neuroprotective drug riluzole. European Journal of Pharmacology. 2002;449(1-2):47–54. doi: 10.1016/s0014-2999(02)01987-8. [DOI] [PubMed] [Google Scholar]
- 164.Wokke J. Riluzole. Lancet. 1996;348(9030):795–799. doi: 10.1016/s0140-6736(96)03181-9. [DOI] [PubMed] [Google Scholar]
- 165.Bellingham M. C. A review of the neural mechanisms of action and clinical efficiency of riluzole in treating amyotrophic lateral sclerosis: what have we learned in the last decade? CNS Neuroscience and Therapeutics. 2011;17(1):4–31. doi: 10.1111/j.1755-5949.2009.00116.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Doble A. The pharmacology and mechanism of action of riluzole. Neurology. 1996;47(6):S233–S241. doi: 10.1212/wnl.47.6_suppl_4.233s. [DOI] [PubMed] [Google Scholar]
- 167.Wang S.-J., Wang K.-Y., Wang W.-C. Mechanisms underlying the riluzole inhibition of glutamate release from rat cerebral cortex nerve terminals (synaptosomes) Neuroscience. 2004;125(1):191–201. doi: 10.1016/j.neuroscience.2004.01.019. [DOI] [PubMed] [Google Scholar]
- 168.Blackburn-Munro G., Ibsen N., Erichsen H. K. A comparison of the anti-nociceptive effects of voltage-activated Na+ channel blockers in the formalin test. European Journal of Pharmacology. 2002;445(3):231–238. doi: 10.1016/s0014-2999(02)01765-x. [DOI] [PubMed] [Google Scholar]
- 169.Coderre T. J., Kumar N., Lefebvre C. D., Yu J. S. C. A comparison of the glutamate release inhibition and anti-allodynic effects of gabapentin, lamotrigine, and riluzole in a model of neuropathic pain. Journal of Neurochemistry. 2007;100(5):1289–1299. doi: 10.1111/j.1471-4159.2006.04304.x. [DOI] [PubMed] [Google Scholar]
- 170.Munro G., Erichsen H. K., Mirza N. R. Pharmacological comparison of anticonvulsant drugs in animal models of persistent pain and anxiety. Neuropharmacology. 2007;53(5):609–618. doi: 10.1016/j.neuropharm.2007.07.002. [DOI] [PubMed] [Google Scholar]
- 171.Abarca C., Silva E., Sepulveda M. J., Oliva P., Contreras E. Neurochemical changes after morphine, dizocilpine or riluzole in the ventral posterolateral thalamic nuclei of rats with hyperalgesia. European Journal of Pharmacology. 2000;403(1-2):67–74. doi: 10.1016/s0014-2999(00)00502-1. [DOI] [PubMed] [Google Scholar]
- 172.Chew D. J., Carlstedt T., Shortland P. J. The effects of minocycline or riluzole treatment on spinal root avulsion–induced pain in adult rats. Journal of Pain. 2014;15(6):664–675. doi: 10.1016/j.jpain.2014.03.001. [DOI] [PubMed] [Google Scholar]
- 173.Hama A., Sagen J. Antinociceptive effect of riluzole in rats with neuropathic spinal cord injury pain. Journal of Neurotrauma. 2011;28(1):127–134. doi: 10.1089/neu.2010.1539. [DOI] [PubMed] [Google Scholar]
- 174.Moon E. S., Karadimas S. K., Yu W.-R., Austin J. W., Fehlings M. G. Riluzole attenuates neuropathic pain and enhances functional recovery in a rodent model of cervical spondylotic myelopathy. Neurobiology of Disease. 2014;62:394–406. doi: 10.1016/j.nbd.2013.10.020. [DOI] [PubMed] [Google Scholar]
- 175.Sung B., Lim G., Mao J. Altered expression and uptake activity of spinal glutamate transporters after nerve injury contribute to the pathogenesis of neuropathic pain in rats. Journal of Neuroscience. 2003;23(7):2899–2910. doi: 10.1523/JNEUROSCI.23-07-02899.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Mishra S. P., Shukla S. K., Pandey B. L. A preliminary evaluation of comparative effectiveness of riluzole in therapeutic regimen for irritable bowel syndrome. Asian Pacific Journal of Tropical Biomedicine. 2014;4:S335–S340. doi: 10.12980/apjtb.4.2014c205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Thompson J. M., Ji G., Neugebauer V. Small-conductance calcium-activated potassium (SK) channels in the amygdala mediate pain-inhibiting effects of clinically available riluzole in a rat model of arthritis pain. Molecular Pain. 2015;11:p. 51. doi: 10.1186/s12990-015-0055-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Han J. S., Neugebauer V. mGluR1 and mGluR5 antagonists in the amygdala inhibit different components of audible and ultrasonic vocalizations in a model of arthritic pain. Pain. 2005;113(1):211–222. doi: 10.1016/j.pain.2004.10.022. [DOI] [PubMed] [Google Scholar]