

Abstract
Melanocortins are a highly conserved family of peptides and receptors that includes multiple proopiomelanocortin-derived peptides and five defined melanocortin receptors. The melanocortins have an important role in maintaining immune homeostasis and in suppressing inflammation. Within the healthy eye, the melanocortins have a central role in preventing inflammation and maintaining immune privilege. A central mediator of the anti-inflammatory activity is the non-steroidogenic melanocortin peptide alpha-melanocyte stimulating hormone. In this review we summarize the major findings of melanocortin regulation of ocular immunobiology with particular interest in the ability of melanocortin to induce immune tolerance and cytoprotection. The melanocortins have therapeutic potential because their mechanisms of action in regulating immunity are distinctly different from the actions of steroids.
Keywords: alpha-MSH, eye, glucocorticoids, melanocortins
INTRODUCTION
The melanocortin (MC) system encompasses multiple peptides including α-, β-, γ-melanocyte stimulating hormone (MSH), adrenocorticotropic hormone (ACTH), and five MC receptors (MCR1-5) that are expressed in a multitude of cells and tissues.1,2 The MC peptides and receptors are highly conserved across multiple species3 from lower vertebrates to vertebrates and appeared early in evolution2 but remain largely unchanged,4 suggesting a critical and ubiquitous role. The role of the MC system in the neuroendocrine control of inflammation is understood and known to be affected by the release of corticotropin-releasing hormone in response to infection or stress; this stimulates the production of pro-opiomelanocortin (POMC), which is processed to generate the MC peptides. The myriad functions of the MCs are carried out through their binding to the five known MC receptors, each of which is expressed in specific cell types.5,6 ACTH stimulates melanocortin receptor 2 (MC2R) on the adrenal cortex to produce glucocorticoids (GCs). Although the anti-inflammatory effects of GCs are well known and are one way in which MCs control inflammation indirectly, there is a growing understanding of the distinct and direct effects of GCs and MCs as immune modulators (Table 1).
TABLE 1.
Effects of glucocorticoids and α–MSH on the eye.
| Glucocorticoid | α–MSH | |
|---|---|---|
| Immune effects | • Suppresses proinflammatory signals • Suppresses immune cell activity |
• Suppresses proinflammatory signals • Promotes production of anti-inflammatory cytokines • Induces suppressor antigen presenting cells • Induces the activation of regulatory T cells • Natural regulator of immunity in healthy eyes |
| Optic nerve effects | • Indirect damage of optic nerve due to increased intraocular pressure • May sensitize nerve cells to disease-induced apoptosis |
• Transiently lowers intraocular pressure • Can protect neuronal cells from damage |
| Adverse effects | • Causes cataracts • Induces increased intraocular pressure and glaucoma • Many other adverse effects |
• No known adverse effects |
It has long been established that GC production is controlled selectively by ACTH, and that ACTH induces steroidogenesis via binding to MC2R. MC2R is abundantly expressed from the adrenal cortex, specifically the zona fasciculata, where the GCs are produced, and the zona glomerulosa, where the mineralocorticoid aldosterone is produced.7 The essential role of MC2R in steroidogenesis is clear as mutations in MC2R result in familial GC deficiencies8 and Mc2r null mice lack adrenal gland development and steroid production.9 The MC receptors have varying affinities for the different MCs, but all recognize a highly conserved sequence (HFRW) that is common to all MC peptides.10 Unlike the other MC receptors, MC2R exclusively binds ACTH7,11 and this specificity is conferred by peptide sequences that are unique to ACTH.12 Animal studies confirm that α-MSH is not capable of stimulating GC secretion13,14 and only ACTH was able to restore wild-type GC production in a Pomc null model.15 Thus, the anti-inflammatory effects of α-MSH are not due to the induction of GC production.
To dissect the direct, non-steroid-mediated effects of MCs in whole organisms multiple approaches have been used. Adrenalectomization,16–18 MCR knockouts,19–23 and administration of specific MCs such as α-MSH24–27 have all been applied to demonstrate the direct pleiotropic effects of the MC system in controlling inflammation. MCs control inflammation through specific downstream modulation of immune cells, processes, cytokines, and chemokines.2,4,28 These studies have helped to uncover how MCs utilize different MCRs in specific cells and tissues to regulate acute, systemic, and chronic inflammation and provide a strong foundation for their use to treat a variety of inflammatory conditions.
ADVERSE EFFECTS OF GLUCOCORTICOIDS ON THE EYE
Although GC therapy has been a mainstay in the treatment of inflammation for decades, it affects many different cells beyond the immune system. Acting via the GC receptor, GCs utilize both transactivation of target genes and transrepression mechanisms to regulate a number of transcription factors including NFkB, AP-1, and p38 MAP kinase29,30 and influence a number of different pathways including stress responses, metabolism, and development. As such, the profile of undesirable side effects from prolonged systemic GC therapy is significant and includes bone loss, diabetes, muscle wasting, impaired wound healing, gastric ulcers, weight gain, cardiovascular disease, and neuropsychiatric issues.31,32
In the eye, the adverse effects of GCs include increased intraocular pressure (IOP), which is directly associated with the development of glaucoma.33–35 IOP is regulated in part by outflow of the aqueous humor via the trabecular meshwork (TM), and increased resistance in this outflow pathway is one of the primary causes of elevated intraocular pressure that can lead to glaucoma.-36 GCs are known to increase outflow resistance of the TM in some individuals.37,38 This GC sensitivity could be due to variation in the relative levels of alternatively spliced GC receptor isoforms in the TM.39,40 GCs have numerous effects on the TM, including changes in TM morphology, gene expression, extracellular matrix, and cytoskeleton as well as inhibition of phagocytosis, proliferation, and migration of TM cells.41 Also, GC treatment causes apoptosis in bovine TM cells,38 and in human TM cells at high concentrations.42 While the exact mechanism by which GCs increase IOP is unclear, it has been reported that 40–60% of patients undergoing chronic GC therapy develop ocular hypertension.43 These adverse effects are also dependent on the type of GC therapy. A recent study on the treatment of uveitis with steroid implants44,45 found that 65% of patients treated with these implants developed increased IOP and 23% developed glaucoma. In comparison, increased IOP was noted in 49% of patients undergoing periocular GC injections,46 and 24% of those receiving systemic therapy, composed of GCs and immunosuppressive medications, developed increased IOP.45
Beyond influencing intraocular pressure, GC therapy has been linked to cataract formation, primarily posterior subcapsular cataracts.47–51 Lens epithelial cells are known to express GC receptors52 and GC treatment induces significant gene expression changes in these cells.53,54 The exact mechanism by which GC treatment induces cataract formation is probably complex. GCs have been shown to induce apoptosis in lens epithelial cells,55,56 but GC treatment also disrupts the differentiation and proliferation of lens cells and causes their aberrant migration to the posterior pole of the lens.57 These factors, as well as changes to the lens environment, via GC effects on other ocular tissues, may all contribute to cataract formation. One meta-analysis of several studies reported that approximately 22% of patients undergoing systemic treatment with GCs developed cataracts, which was correlated with the dosage and duration of treatment.49 Similarly, 20% of patients receiving periocular GC injections developed cataracts within 12 months.46 However, a recent study found that 91% of patients receiving steroid implants for uveitis developed cataracts within 24 months.44
α-MSH AND THE EYE
The microenvironment of the eye is immune privileged in that it has several mechanisms to suppress the action of inflammation. Over the past couple of decades it has become clear that melanocortin pathways have an important role in maintaining immune privilege.58–60 The melanocortin pathways are involved in suppressing proinflammatory signals and, unlike steroids, promote the immune system to regulate itself within the ocular microenvironment. Aqueous humor, the fluid filling the anterior chamber of the eye, suppresses the in vitro activation of immune cells that mediate hypersensitivity;61 moreover, these aqueous humor-treated immune cells are able to suppress other hypersensitivity-mediating T cells.62 At the molecular level this immunoregulatory activity of aqueous humor is mediated by α-MSH. Although TGF-β2 is the most abundant anti-inflammatory factor in aqueous humor,63–65 it exists in a latent form and must be activated before it can affect immune cells whereas the effects of α-MSH are more immediate. In vitro studies have demonstrated that TGF-β2 is an enhancer of the immune regulating activity induced by α-MSH.66
In the healthy eye there is constitutive expression of α-MSH. In aqueous humor the physiological concentration is around 30 pg/mL and the level of α-MSH is relatively constant among mammals. The sources of α-MSH production in the eye are not fully known; however, retinal pigment epithelial (RPE) cells have been shown to be a source of α-MSH.60 In culture, RPE cells produce about 2 ng of α-MSH in 24 hours58,60 and express the necessary endopeptidases and post-translational enzymes to process a functional α-MSH peptide from POMC.67 Other cells in the eye, such as the cells of the iris and ciliary body, also express these enzymes, suggesting that they are also potential sources of α-MSH.68 At ocular physiological concentrations under serum-free conditions in vitro, as found within the ocular microenvironment, α-MSH is highly potent and suppresses most signals of inflammation while stimulating regulatory activity.26,58,60,62,66,69 Evidence suggests that there is a loss of α-MSH expression in eyes with autoimmune disease or damaged retinas.60 Treatment of mice suffering from ocular inflammation (uveitis) with α-MSH suppresses the inflammation, and may re-establish some features of immune privilege and immune regulation.22,70–72 The finding of α-MSH activity within the eye has not only promoted examination of the role of neuropeptides in regulating immunity but, more importantly, has also provided molecular mechanisms for the anti-inflammatory activity seen within the immune privileged eye.
Activated effector T cells treated with aqueous humor or α-MSH produce their own TGF-β, are FoxP3 positive, and function as CD4+CD25+ inducible Treg (iTreg) cells.69,73 In addition, these α-MSH-induced iTreg cells require reactivation to their cognate antigen to mediate suppression, although the mechanism of suppression mediated by the α-MSH– induced iTreg cells is non-specific and will suppress all inflammatory activity in their immediate microenvironment. The induction of regulatory immunity instead of complete suppression of inflammation provides the eye with a directed mechanism of protection against autoimmune attack. Induction of regulatory immunity by α-MSH is mediated through its effects on both the antigen presenting cells (APC) and on effector T cells through their expression of melanocortin receptors 1, 3, and 5.26,72,74–77 The induction of regulatory immunity and signaling through melanocortin receptors other than MC2R provides a strong indication that α-MSH might be an alternative non-steroidogenic therapeutic approach to regulate immunity.
The RPE, through α-MSH together with another neuropeptide NPY, promotes the differentiation of macrophages into suppressor cells that suppress inflammation and immunity.60 This regulation also appears to convert the retinal microglial cells into suppressor cells. The benefit of such cells is that, although they occupy a tissue microenvironment that is hypoxic and full of oxidized byproducts, they are not activated to mediate inflammation;60,78,79 on the contrary, they suppress the activation of inflammation and effector T cells. RPE cells from wounded retinas are diminished in α-MSH production and support the activation of macrophages that mediate inflammation.60 Therefore, in ocular disease, changes in RPE production of α-MSH may contribute to the disease pathology. The RPE-induced suppressor cells are not themselves suppressed in immune activity. While they continue to phagocytize opsonized materials, α-MSH suppresses the activation of phagolysosomes.80,81 The implication is that although α-MSH-affected cells can clear materials through phagocytosis, they cannot process or degrade the materials through conventional acidic-lysosome pathways. This may be a mechanism of immune privilege by which processing of recognizable autoantigen peptides for MHC presentation is prevented. One interesting phenomenon is that, without α-MSH, healthy RPE cells induce apoptosis in macrophages.60,81 While the mechanism by which RPE induces apoptosis in the macrophages is unknown, this unique finding suggests that a macrophage responding to α-MSH is protected from apoptosis;82 however, such a rescued macrophage will now be a suppressor cell. This could be part of a mechanism of immune privilege to select for immune cells that can be manipulated to be suppressor cells within the healthy retina. This mechanism is highly dependent on the production of α-MSH by the RPE.
There is increasing evidence that specific melanocortin receptors may be associated with different actions of α-MSH (Figure 1). While the suppression of pro-inflammatory signals on innate immune cells by α-MSH is thought to occur most often through MC1R and MC3R,74,75 the activation of regulatory activity appears to be through MC5R.26,77 The role of MC5R in regulation is seen in the difference between wild-type mice and Mc5r knockout mice that have recovered from experimental autoimmune uveitis (EAU).83 Mice, unlike humans, recover from EAU without medical intervention. Post-EAU spleens of wild-type mice exhibit regulatory immunity that provides resistance to reactivation of the autoimmune uveitis.77,84 Mice without MC5R recover from EAU on their own, but they lack this regulatory immunity in the spleen.84 If the post-EAU Mc5r knockout mice are reimmunized to reactivate EAU, they display an immediate reactivation of uveitis that becomes extremely severe. If the spleen cells from post-EAU wild-type mice are adoptively transferred to Mc5r-knockout mice before the reimmunization, the second episode is suppressed. This finding demonstrates that mice possess a melanocortin-dependent induction of regulatory immunity that prevents a memory immune response to autoantigen of the retina and a systemic resistance to retinal autoantigens.
FIGURE 1.
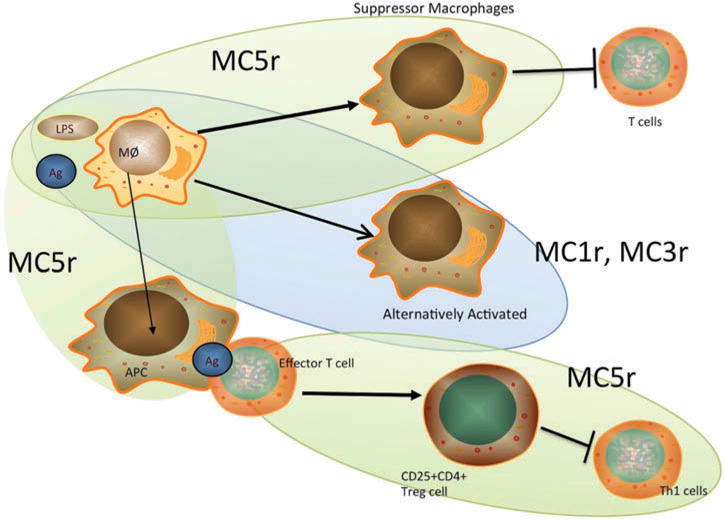
Effects of different melanocortin receptors on immune activity. Through MC1r, MC3r, and MC5r, α-MSH suppresses inflammation and promotes the activation of anti-inflammatory activity and regulatory immunity. In macrophages, through MC1r and MC3r, α-MSH mediates alternative activation by which the macrophages suppress inflammation.74,75 Through MC5r, α-MSH mediates the activation of suppressor cell activity in macrophages that suppresses effector T cell activation and viability.58,60 Through MC5r on T cells, α-MSH promotes the activation of regulatory T cell activity and the suppression of effector T cells.26,69 In addition, through MC5r, α-MSH induces antigen presenting cells to promote activation inducible Treg cells72, 77 (see Figure 2). Ag, antigen; LPS, lipopolysaccharide; MØ, macrophage.
The MC5R-dependent regulatory immunity can be induced during uveitis by injecting α-MSH peptide into the EAU mice.22,72,77 This has also been demonstrated in the mouse model of multiple sclerosis, experimental autoimmune encephalomyelitis.85,86 Both antigen presenting cells (APC) and effector T cells express MC5R, and can be the target of α-MSH induction of regulatory activity. As described above, treating activated effector T cells with α-MSH induces the expansion and activation of inducible Treg cells.-26,69 Blocking MC5R or using T cells from immunized MC5R knock-out mice prevents α-MSH induction of regulatory activity. α-MSH upregulates expression of FoxP3 in T cells, indicating that α-MSH promotes T cell differentiation into regulatory cells.69 There is very little cytokine production by these α-MSH-treated T cells, other than TGF-β. The cells function as regulatory T cells, and through adoptive transfer, suppress inflammation within the tissue microenvironment where their antigen specificity is presented.62,77,87 Therefore, by stimulating MC5r with α-MSH peptide or a melanocortin analogue that targets only MC5r, it is possible to create a unique population of Treg cells from antigen-specific T cells that can be used to target tissues of inflammation, graft rejection, and autoimmune disease.
When the spleen cells from the post-EAU mice were examined, it was not the T cells but rather the APC that were MC5R dependent for regulatory activity.77 These APC are unique in that they express markers of both myeloid and granulocytic cells. They are found in the spleens of mice, but mediate regulatory immunity only after EAU and only if MC5R is expressed. Resting APC stimulated by α-MSH express the ectoenzymes that convert ATP into adenosine.72 There is also an increase in their expression of PD-L1, a surface adhesion protein that regulates T cell activation.73 They process and present antigen and, when mixed with antigen specific effector T cells, the APC promote regulatory activity by the T cells. These Treg cells have the characteristics of inducible Treg cells, suggesting that α-MSH, through MC5R, upregulates APC generation of adenosine and expression of PD-L1 to promote the counterconversion of effector T cells into inducible Treg cells.72,73 Therefore, as part of the resolution of EAU, there is a melanocortin-dependent pathway that localizes in the spleen a unique population of APC that promote the activation of protective autoreactive T cells (Figure 2). This pathway can be induced by α-MSH therapy and, since expression of the melanocortins is conserved, has the potential to be a new therapeutic approach to suppression of autoimmune disease in humans. It will be of interest to see whether this is a pathway that, while spontaneously activated in mice, needs to be awakened in humans with a melanocortin-based therapy.
FIGURE 2.
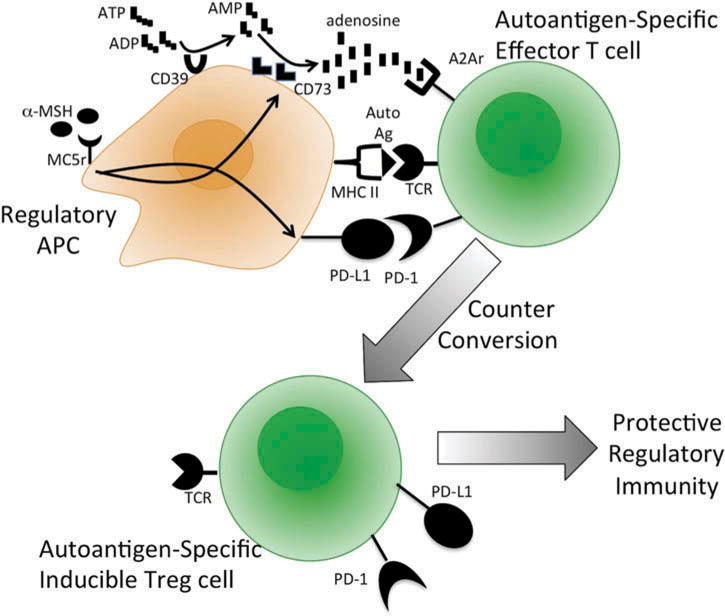
Melanocortin-driven induction of autoantigen-specific protective immunity. Self-resolution or α-MSH treatment of experimental autoimmune uveitis in mice is followed by the induction of autoantigen-specific Treg cells that provide resistance to the recurrence of uveitis.72,84 This is driven by the action of α-MSH, through MC5r on macrophages, to induce antigen presenting activity that counter-converts autoantigen-specific effector T cells into inducible Treg cells.2,77 α-MSH upregulates APC generation of adenosine and expression of PD-L1.72 This process requires autoantigen-specific effector T cells expressing the adenosine 2A receptor.72 Ag, antigen, CD39, ecto-nucleoside triphosphate diphosphohydrolase 1; CD73, ecto-5′-nucleotidase; MHC II, major histocompatibility class II antigen; TCR, T-cell receptor.
These studies have shown that the MCs have an extremely important role in preventing inflammation within the eye. In addition, there is strong potential for use of the MC system in therapy for uveitis. Also, therapeutic benefits of using the MCs may go beyond immune regulation within the eye. Through MC1R, α-MSH protects the RPE from oxidative stress-induced apoptosis.88 In addition, α-MSH may have neurotrophic activity that promotes photoreceptor survival.89 This suggests that, besides the anti-inflammatory and immune regulating activity of α-MSH therapy, there will be an additional benefit of maintaining and promoting the health of other cells of the retina. Therefore, the MCs hold an important role in maintaining an anti-inflammatory microenvironment and in the general health of the retina. Moreover, the anti-inflammatory and immune-regulating activity of the melanocortins has the potential to provide a novel therapeutic approach to many autoimmune and inflammatory diseases. This approach contrasts with the general suppression of immunity by GCs and with the biologic therapies that target one specific proinflammatory cytokine at a time. MC therapy manipulates the immune cells to function in a manner that suppresses inflammation and promotes immune tolerance and cell survival to potentially regenerate a healthy ocular microenvironment (Figure 3).
FIGURE 3.
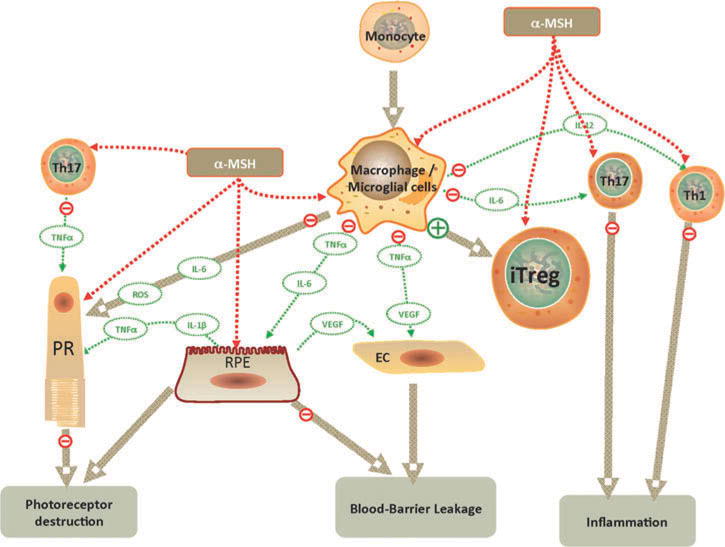
Effects of α-MSH on uveitis and retinal cell health. Therapeutic application of α-MSH has the potential to suppress inflammation, induce immune tolerance, and promote retinal cell survival (see text). α-MSH can directly influence immune cells through their expression of melanocortin receptors to suppress activation of effector T cells (Th1, Th17 cells), while promoting iTreg cell activation. In macrophages, α-MSH suppresses production of proinflammatory cytokines and promotes antigen presenting activity that converts effector T cells into functional iTreg cells. In addition, RPE and photoreceptors express melanocortin receptors through which α-MSH promotes cell survival. EC, endothelial cell; IL-1β, -6, -12; interleukin-1beta, -6, -12; iTreg: inducible Treg cells; RPE, retinal pigment epithelial cells; PR, photoreceptor cell; ROS, reactive oxygen species; Th: T helper cells; TNFα; tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor.
MELANOCORTINS AS ALTERNATIVES TO GLUCOCORTICOIDS
GCs have been the primary therapeutic option for the treatment of ocular inflammatory disorders for over 60 years.90 Despite the popularity of GCs for treatment, they have a number of side effects that cause serious complications. As mentioned above, a significant portion of the population is susceptible to increased IOP from treatment with GCs. In addition to the well-documented relationship of GCs with glaucoma and cataracts, GC therapy can also lead to a number of other adverse effects in the eye including viral retinitis-91 and central serous chorioretinopathy,92,93 which is characterized by serous fluid accumulation in the retina resulting in localized detachment of the neurosensory retina.94 GC therapy is also associated with an increased risk of infection, impaired wound healing, and thinning of the cornea.95,96
In contrast, α-MSH is present in the aqueous humor of the eye and acts to prevent inflammation that could be damaging to the eye.58,97 α-MSH has the ability to influence T cells in a positive immune modulating capacity by suppressing immune response through the release of inhibitory cytokines, metabolic inhibition, or cytolysis of effector T-cells and by suppressing the maturation of dendritic cells.58,59,66,69,98,99 α-MSH suppresses production of proinflammatory cytokines, including IFN-γ58 and TNFα,100 while increasing production of anti-inflammatory cytokines such as IL-10.101 In addition to its anti-inflammatory actions, α-MSH has been shown to suppress apoptosis in a number of cell types.82,102–104 The immune modulation exhibited by α-MSH, coupled with its distinctive cytoprotective effect, makes α-MSH a potentially advantageous alternative treatment and has generated a number of mechanistic studies to examine α-MSH as a therapeutic tool in the treatment of ocular inflammation. Use of α-MSH and its analogs to treat uveitis in a variety of model systems has shown them to be effective in reducing the severity and promoting resolution of inflammation.22,70,105 In addition to the treatment of inflammatory disorders, topical α-MSH has been shown to transiently lower intraocular pressure,106 which contrasts with the increase in pressure often seen with GC treatment. While the anti-inflammatory actions of GCs suppress the immune system, MCs appear to have multipathway immunomodulatory properties to handle the acute inflammation with pro-resolving properties that alter the phenotype of immune cells, allowing them to be more modulatory.20
MCs have potent anti-inflammatory and cytoprotective functions in reducing inflammation and inducing resolution in other tissues and conditions. α-MSH normalizes oxidative stress, diminishes apoptosis, and reduces retinal damage in diabetic retinopathy.104,107 In rodent model systems, MCs promote restoration of nerve damage from spinal cord injury,108,109 modulate inflammation, and attenuate apoptosis and damage after traumatic brain injury.110 Recently, α-MSH has been demonstrated to protect against brain damage and cognitive decline in murine models of Alzheimer’s disease.111–113
The neuroprotective functions of α-MSH can also reduce the severity of autoimmune encephalomyelitis (EAE) in rodent models.85,114 This contrasts with GCs, which have been found to increase apoptosis of retinal nerve ganglion cells in EAE models.115 EAE is an inflammatory demyelinating disease of the central nervous system which, importantly, is a model system for multiple sclerosis (MS). ACTH has been known to be effective in the treatment of MS for many years.116 Historically, the effects of ACTH therapy in MS were thought to be due to the promotion of steroidogenesis through binding of MC2R. However, several studies now indicate that ACTH functions in alleviating MS exacerbations via binding to other MC receptors rather than by the induction of steroid production.117 This, along with results from model systems such as EAE, suggests that MS may be successfully treated with MC peptides.
MCs also exhibit cytoprotective properties that oppose the damaging effects of GCs. A significant side effect of systemic GC therapy is osteoporosis.118 Although ACTH induces steroidogenesis, this results in a lower plasma level of GCs than typical steroid administration119,120 and, moreover, has a cytoprotective effect on bone and can protect against osteonecrosis induced by GCs.121 In addition, while GC therapy can increase the risk of infection, α-MSH and its analogues have been shown to have significant antimicrobial activity.122,123
The differences noted above suggest that MCs may offer a less toxic alternative to GCs for the treatment of inflammation. Toxicity studies found that massive doses of α-MSH, 5000-fold greater than the dose necessary to affect thermoregulation in rabbits, were required before toxicity became lethal in some animals.124 Furthermore, α-MSH and analogues have been shown to have no significant adverse effects in clinical studies, especially in comparison to other anti-inflammatory therapies.125–131
CONCLUSION
Melanocortins have shown promise in the treatment of a number of autoimmune disorders. α-MSH administration reduces disease activity in model systems of rheumatoid arthritis,132 systemic lupus erythematosus,133 and inflammatory bowel disease.134 Recent evidence has shown that ACTH is effective in lupus,135 dermatomyositis or polymyositis,136 infantile spasms, MS, and nephrotic syndrome.137 These are all indications in which steroids are generally considered the mainstay of treatment, and where their toxic side effects often require an alternative therapy. Importantly, in several of these indications, MCs have shown to be efficacious where steroids alone have been ineffective,137–139 further suggesting their mechanism of action is distinct from steroids (Table 1).
Acknowledgments
Writing support for this manuscript was provided by Kelly P. Smith, and financial support for AWT was provided in part from the Massachusetts Lions Eye Research Foundation.
Footnotes
DECLARATION OF INTEREST
CMC and JY are employees of Mallinckrodt Pharmaceuticals, and AWT has been a consultant for Mallinckrodt Pharmaceuticals.
References
- 1.Catania A, Gatti S, Colombo G, et al. Targeting melanocortin receptors as a novel strategy to control inflammation. Pharmacol Rev. 2004;56(1):1–29. doi: 10.1124/pr.56.1.1. [DOI] [PubMed] [Google Scholar]
- 2.Catania A, Lonati C, Sordi A, et al. The melanocortin system in control of inflammation. ScientificWorldJournal. 2010;10:1840–1853. doi: 10.1100/tsw.2010.173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Schioth HB, Haitina T, Ling MK, et al. Evolutionary conservation of the structural, pharmacological, and genomic characteristics of the melanocortin receptor subtypes. Peptides. 2005;26(10):1886–1900. doi: 10.1016/j.peptides.2004.11.034. [DOI] [PubMed] [Google Scholar]
- 4.Ottaviani E, Franchini A, Genedani S. ACTH and its role in immune-neuroendocrine functions. A comparative study. Curr Pharm Des. 1999;5(9):673–681. [PubMed] [Google Scholar]
- 5.Cooray SN, Clark AJ. Melanocortin receptors and their accessory proteins. Mol Cell Endocrinol. 2011;331(2):215–221. doi: 10.1016/j.mce.2010.07.015. [DOI] [PubMed] [Google Scholar]
- 6.Cone RD, Mountjoy KG, Robbins LS, et al. Cloning and functional characterization of a family of receptors for the melanotropic peptides. Ann N Y Acad Sci. 1993;680:342–363. doi: 10.1111/j.1749-6632.1993.tb19694.x. [DOI] [PubMed] [Google Scholar]
- 7.Mountjoy KG, Robbins LS, Mortrud MT, et al. The cloning of a family of genes that encode the melanocortin receptors. Science. 1992;257(5074):1248–1251. doi: 10.1126/science.1325670. [DOI] [PubMed] [Google Scholar]
- 8.Clark AJ, McLoughlin L, Grossman A. Familial glucocorticoid deficiency associated with point mutation in the adrenocorticotropin receptor. Lancet. 1993;341(8843):461–462. doi: 10.1016/0140-6736(93)90208-x. [DOI] [PubMed] [Google Scholar]
- 9.Chida D, Nakagawa S, Nagai S, et al. Melanocortin 2 receptor is required for adrenal gland development, steroidogenesis, and neonatal gluconeogenesis. Proc Natl Acad Sci U S A. 2007;104(46):18205–18210. doi: 10.1073/pnas.0706953104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Schioth HB, Muceniece R, Larsson M, et al. Binding of cyclic and linear MSH core peptides to the melanocortin receptor subtypes. Eur J Pharmacol. 1997;319(2–3):369–373. doi: 10.1016/s0014-2999(96)00959-4. [DOI] [PubMed] [Google Scholar]
- 11.Schioth HB, Chhajlani V, Muceniece R, et al. Major pharmacological distinction of the ACTH receptor from other melanocortin receptors. Life Sci. 1996;59(10):797–801. doi: 10.1016/0024-3205(96)00370-0. [DOI] [PubMed] [Google Scholar]
- 12.Costa JL, Bui S, Reed P, et al. Mutational analysis of evolutionarily conserved ACTH residues. Gen Comp Endocrinol. 2004;136(1):12–16. doi: 10.1016/j.ygcen.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 13.Nussdorfer GG, Mazzocchi G, Malendowicz LK. Acute effects of alpha-MSH on the rat zona glomerulosa in vivo. Biochem Biophys Res Commun. 1986;141(3):1279–1284. doi: 10.1016/s0006-291x(86)80184-x. [DOI] [PubMed] [Google Scholar]
- 14.Shenker Y, Villareal JZ, Sider RS, et al. Alpha-melanocyte-stimulating hormone stimulation of aldosterone secretion in hypophysectomized rats. Endocrinology. 1985;116(1):138–141. doi: 10.1210/endo-116-1-138. [DOI] [PubMed] [Google Scholar]
- 15.Costa JL, Forbes S, Brennan MB, et al. Genetic modifications of mouse proopiomelanocortin peptide processing. Mol Cell Endocrinol. 2011;336(1–2):14–22. doi: 10.1016/j.mce.2010.12.032. [DOI] [PubMed] [Google Scholar]
- 16.Getting SJ, Christian HC, Flower RJ, et al. Activation of melanocortin type 3 receptor as a molecular mechanism for adrenocorticotropic hormone efficacy in gouty arthritis. Arthritis Rheum. 2002;46(10):2765–2775. doi: 10.1002/art.10526. [DOI] [PubMed] [Google Scholar]
- 17.Gubner R, Cote L, Hughes J, et al. Comparative effects of aminopterin, cortisone and ACTH in experimental formaldehyde arthritis and psoriatic arthritis. J Invest Dermatol. 1952;19(4):297–305. doi: 10.1038/jid.1952.100. [DOI] [PubMed] [Google Scholar]
- 18.Yeh JK, Evans JF, Niu QT, et al. A possible role for melanocortin peptides in longitudinal growth. J Endocrinol. 2006;191(3):677–686. doi: 10.1677/joe.1.06729. [DOI] [PubMed] [Google Scholar]
- 19.Auriemma M, Brzoska T, Klenner L, et al. alpha-MSH-stimulated tolerogenic dendritic cells induce functional regulatory T cells and ameliorate ongoing skin inflammation. J Invest Dermatol. 2012;132(7):1814–1824. doi: 10.1038/jid.2012.59. [DOI] [PubMed] [Google Scholar]
- 20.Montero-Melendez T, Patel HB, Seed M, et al. The melanocortin agonist AP214 exerts anti-inflammatory and pro-resolving properties. Am J Pathol. 2011;179(1):259–269. doi: 10.1016/j.ajpath.2011.03.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Patel HB, Bombardieri M, Sampaio AL, et al. Anti-inflammatory and antiosteoclastogenesis properties of endogenous melanocortin receptor type 3 in experimental arthritis. FASEB J. 2010;24(12):4835–4843. doi: 10.1096/fj.10-167759. [DOI] [PubMed] [Google Scholar]
- 22.Lee DJ, Biros DJ, Taylor AW. Injection of an alpha-melanocyte stimulating hormone expression plasmid is effective in suppressing experimental autoimmune uveitis. Int Immunopharmacol. 2009;9(9):1079–1086. doi: 10.1016/j.intimp.2009.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lee SN, Ryu JH, Joo JH, et al. Alpha-melanocyte-stimulating hormone inhibits tumor necrosis factor alpha-stimulated MUC5AC expression in human nasal epithelial cells. Am J Respir Cell Mol Biol. 2011;44(5):716–724. doi: 10.1165/rcmb.2009-0420OC. [DOI] [PubMed] [Google Scholar]
- 24.Cooper A, Robinson SJ, Pickard C, et al. Alpha-melanocyte-stimulating hormone suppresses antigen-induced lymphocyte proliferation in humans independently of melanocortin 1 receptor gene status. J Immunol. 2005;175(7):4806–4813. doi: 10.4049/jimmunol.175.7.4806. [DOI] [PubMed] [Google Scholar]
- 25.Buggy JJ. Binding of alpha-melanocyte-stimulating hormone to its G-protein-coupled receptor on B-lymphocytes activates the Jak/STAT pathway. Biochem J. 1998;331(Pt 1):211–216. doi: 10.1042/bj3310211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Taylor A, Namba K. In vitro induction of CD25+ CD4+ regulatory T cells by the neuropeptide alpha-melanocyte stimulating hormone (alpha-MSH) Immunol Cell Biol. 2001;79(4):358–367. doi: 10.1046/j.1440-1711.2001.01022.x. [DOI] [PubMed] [Google Scholar]
- 27.Loser K, Brzoska T, Oji V, et al. The neuropeptide alpha-melanocyte-stimulating hormone is critically involved in the development of cytotoxic CD8+ T cells in mice and humans. PLoS One. 2010;5(2):e8958. doi: 10.1371/journal.pone.0008958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brzoska T, Luger TA, Maaser C, et al. Alpha-melanocyte-stimulating hormone and related tripeptides: biochemistry, antiinflammatory and protective effects in vitro and in vivo, and future perspectives for the treatment of immune-mediated inflammatory diseases. Endocr Rev. 2008;29(5):581–602. doi: 10.1210/er.2007-0027. [DOI] [PubMed] [Google Scholar]
- 29.Ratman D, Vanden Berghe W, Dejager L, et al. How glucocorticoid receptors modulate the activity of other transcription factors: a scope beyond tethering. Mol Cell Endocrinol. 2013;380(1–2):41–54. doi: 10.1016/j.mce.2012.12.014. [DOI] [PubMed] [Google Scholar]
- 30.Cruz-Topete D, Cidlowski JA. One hormone, two actions: anti- and pro-inflammatory effects of glucocorticoids. Neuroimmunomodulation. 2015;22(1–2):20–32. doi: 10.1159/000362724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Schacke H, Docke WD, Asadullah K. Mechanisms involved in the side effects of glucocorticoids. Pharmacol Ther. 2002;96(1):23–43. doi: 10.1016/s0163-7258(02)00297-8. [DOI] [PubMed] [Google Scholar]
- 32.Harris E, Tiganescu A, Tubeuf S, et al. The prediction and monitoring of toxicity associated with long-term systemic glucocorticoid therapy. Curr Rheumatol Rep. 2015;17(6):513. doi: 10.1007/s11926-015-0513-4. [DOI] [PubMed] [Google Scholar]
- 33.Sommer A, Tielsch JM, Katz J, et al. Relationship between intraocular pressure and primary open angle glaucoma among white and black Americans. The Baltimore Eye Survey. Arch Ophthalmol. 1991;109(8):1090–1095. doi: 10.1001/archopht.1991.01080080050026. [DOI] [PubMed] [Google Scholar]
- 34.AGIS-Investigators. The Advanced Glaucoma Intervention Study (AGIS): 7. The relationship between control of intraocular pressure and visual field deterioration.The AGIS Investigators. Am J Ophthalmol. 2000;130(4):429–440. doi: 10.1016/s0002-9394(00)00538-9. [DOI] [PubMed] [Google Scholar]
- 35.Gordon MO, Beiser JA, Brandt JD, et al. The Ocular Hypertension Treatment Study: baseline factors that predict the onset of primary open-angle glaucoma. Arch Ophthalmol. 2002;120(6):714–720. doi: 10.1001/archopht.120.6.714. discussion 829–730. [DOI] [PubMed] [Google Scholar]
- 36.Tan JC, Peters DM, Kaufman PL. Recent developments in understanding the pathophysiology of elevated intraocular pressure. Curr Opin Ophthalmol. 2006;17(2):168–174. doi: 10.1097/01.icu.0000193079.55240.18. [DOI] [PubMed] [Google Scholar]
- 37.Wordinger RJ, Clark AF. Effects of glucocorticoids on the trabecular meshwork: towards a better understanding of glaucoma. Prog Retin Eye Res. 1999;18(5):629–667. doi: 10.1016/s1350-9462(98)00035-4. [DOI] [PubMed] [Google Scholar]
- 38.Gu Y, Zeng S, Qiu P, et al. Apoptosis of bovine trabecular meshwork cells induced by dexamethasone. Zhonghua Yan Ke Za Zhi. 2002;38(5):302–304. [PubMed] [Google Scholar]
- 39.Zhang X, Clark AF, Yorio T. Regulation of glucocorticoid responsiveness in glaucomatous trabecular meshwork cells by glucocorticoid receptor-beta. Invest Ophthalmol Vis Sci. 2005;46(12):4607–4616. doi: 10.1167/iovs.05-0571. [DOI] [PubMed] [Google Scholar]
- 40.Jain A, Wordinger RJ, Yorio T, et al. Role of the alternatively spliced glucocorticoid receptor isoform GRbeta in steroid responsiveness and glaucoma. J Ocul Pharmacol Ther. 2014;30(2–3):121–127. doi: 10.1089/jop.2013.0239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Clark AF, Wordinger RJ. The role of steroids in outflow resistance. Exp Eye Res. 2009;88(4):752–759. doi: 10.1016/j.exer.2008.10.004. [DOI] [PubMed] [Google Scholar]
- 42.Sharma A, Patil AJ, Mansoor S, et al. Effects of dexamethasone on human trabecular meshwork cells in vitro. Graefes Arch Clin Exp Ophthalmol. 2013;251(7):1741–1746. doi: 10.1007/s00417-013-2343-2. [DOI] [PubMed] [Google Scholar]
- 43.Lewis JM, Priddy T, Judd J, et al. Intraocular pressure response to topical dexamethasone as a predictor for the development of primary open-angle glaucoma. Am J Ophthalmol. 1988;106(5):607–612. doi: 10.1016/0002-9394(88)90595-8. [DOI] [PubMed] [Google Scholar]
- 44.Kempen JH, Altaweel MM, Holbrook JT, et al. Randomized comparison of systemic anti-inflammatory therapy versus fluocinolone acetonide implant for intermediate, posterior, and panuveitis: the multicenter uveitis steroid treatment trial. Ophthalmology. 2011;118(10):1916–1926. doi: 10.1016/j.ophtha.2011.07.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Friedman DS, Holbrook JT, Ansari H, et al. Risk of elevated intraocular pressure and glaucoma in patients with uveitis: results of the multicenter uveitis steroid treatment trial. Ophthalmology. 2013;120(8):1571–1579. doi: 10.1016/j.ophtha.2013.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sen HN, Vitale S, Gangaputra SS, et al. Periocular corticosteroid injections in uveitis: effects and complications. Ophthalmology. 2014;121(11):2275–2286. doi: 10.1016/j.ophtha.2014.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Black RL, Oglesby RB, Von Sallmann L, et al. Posterior subcapsular cataracts induced by corticosteroids in patients with rheumatoid arthritis. JAMA. 1960;174:166–171. doi: 10.1001/jama.1960.63030020005014. [DOI] [PubMed] [Google Scholar]
- 48.Butcher JM, Austin M, McGalliard J, et al. Bilateral cataracts and glaucoma induced by long term use of steroid eye drops. BMJ. 1994;309(6946):43. doi: 10.1136/bmj.309.6946.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Urban RC, Jr, Cotlier E. Corticosteroid-induced cataracts. Surv Ophthalmol. 1986;31(2):102–110. doi: 10.1016/0039-6257(86)90077-9. [DOI] [PubMed] [Google Scholar]
- 50.Costagliola C, Cati-Giovannelli B, Piccirillo A, et al. Cataracts associated with long-term topical steroids. Br J Dermatol. 1989;120(3):472–473. doi: 10.1111/j.1365-2133.1989.tb04181.x. [DOI] [PubMed] [Google Scholar]
- 51.Klein BE, Klein R, Lee KE, et al. Drug use and five-year incidence of age-related cataracts: The Beaver Dam Eye Study. Ophthalmology. 2001;108(9):1670–1674. doi: 10.1016/s0161-6420(01)00656-x. [DOI] [PubMed] [Google Scholar]
- 52.James ER, Robertson L, Ehlert E, et al. Presence of a transcriptionally active glucocorticoid receptor alpha in lens epithelial cells. Invest Ophthalmol Vis Sci. 2003;44(12):5269–5276. doi: 10.1167/iovs.03-0401. [DOI] [PubMed] [Google Scholar]
- 53.James ER, Fresco VM, Robertson LL. Glucocorticoid-induced changes in the global gene expression of lens epithelial cells. J Ocul Pharmacol Ther. 2005;21(1):11–27. doi: 10.1089/jop.2005.21.11. [DOI] [PubMed] [Google Scholar]
- 54.Gupta V, Galante A, Soteropoulos P, et al. Global gene profiling reveals novel glucocorticoid induced changes in gene expression of human lens epithelial cells. Mol Vis. 2005;11:1018–1040. [PubMed] [Google Scholar]
- 55.Petersen A, Carlsson T, Karlsson JO, et al. Effects of dexamethasone on human lens epithelial cells in culture. Mol Vis. 2008;14:1344–1352. [PMC free article] [PubMed] [Google Scholar]
- 56.Sharma A, Pirouzmanesh A, Patil J, et al. Evaluation of the toxicity of triamcinolone acetonide and dexamethasone sodium phosphate on human lens epithelial cells (HLE B-3) J Ocul Pharmacol Ther. 2011;27(3):265–271. doi: 10.1089/jop.2010.0120. [DOI] [PubMed] [Google Scholar]
- 57.James ER. The etiology of steroid cataract. J Ocul Pharmacol Ther. 2007;23(5):403–420. doi: 10.1089/jop.2006.0067. [DOI] [PubMed] [Google Scholar]
- 58.Taylor AW, Streilein JW, Cousins SW. Identification of alpha-melanocyte stimulating hormone as a potential immunosuppressive factor in aqueous humor. Curr Eye Res. 1992;11(12):1199–1206. doi: 10.3109/02713689208999545. [DOI] [PubMed] [Google Scholar]
- 59.Taylor AW, Alard P, Yee DG, et al. Aqueous humor induces transforming growth factor-beta (TGF-beta)-producing regulatory T-cells. Curr Eye Res. 1997;16(9):900–908. doi: 10.1076/ceyr.16.9.900.5043. [DOI] [PubMed] [Google Scholar]
- 60.Kawanaka N, Taylor AW. Localized retinal neuropeptide regulation of macrophage and microglial cell functionality. Journal of Neuroimmunology. 2011;232(1–2):17–25. doi: 10.1016/j.jneuroim.2010.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kaiser CJ, Ksander BR, Streilein JW. Inhibition of lymphocyte proliferation by aqueous humor. Regional Immunology. 1989;2(1):42–49. [PubMed] [Google Scholar]
- 62.Nishida T, Taylor A. Specific aqueous humor factors induce activation of regulatory T cells. Investigative Ophthalmology & Visual Science. 1999;40(10):2268–2274. [PubMed] [Google Scholar]
- 63.Cousins SW, McCabe MM, Danielpour D, et al. Identification of transforming growth factor-beta as an immunosuppressive factor in aqueous humor. Invest Ophthalmol Vis Sci. 1991;32:33–43. [PubMed] [Google Scholar]
- 64.Granstein R, Staszewski R, Knisely T, et al. Aqueous humor contains transforming growth factor-b and a small (<3500 daltons) inhibitor of thymocyte proliferation. Journal of Immunology. 1990;144:3021–3027. [PubMed] [Google Scholar]
- 65.Jampel HD, Roche N, Stark WJ, et al. Transforming growth factor-beta in human aqueous humor. Curr Eye Res. 1990;9(10):963–969. doi: 10.3109/02713689009069932. [DOI] [PubMed] [Google Scholar]
- 66.Namba K, Kitaichi N, Nishida T, et al. Induction of regulatory T cells by the immunomodulating cytokines alpha-melanocyte-stimulating hormone and transforming growth factor-beta2. J Leukoc Biol. 2002;72(5):946–952. [PubMed] [Google Scholar]
- 67.Zmijewski MA, Sharma RK, Slominski AT. Expression of molecular equivalent of hypothalamic-pituitary-adrenal axis in adult retinal pigment epithelium. The Journal of endocrinology. 2007;193(1):157–169. doi: 10.1677/joe.1.06927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ortego J, Coca-Prados M. Molecular characterization and differential gene induction of the neuroendocrine-specific genes neurotensin, neurotensin receptor, PC1, PC2, and 7B2 in the human ocular ciliary epithelium. J Neurochem. 1997;69(5):1829–1839. doi: 10.1046/j.1471-4159.1997.69051829.x. [DOI] [PubMed] [Google Scholar]
- 69.Taylor AW, Lee DJ. The alpha-melanocyte stimulating hormone induces conversion of effector T cells into treg cells. J Transplant. 2011;2011:246856. doi: 10.1155/2011/246856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Nishida T, Miyata S, Itoh Y, et al. Anti-inflammatory effects of alpha-melanocyte-stimulating hormone against rat endotoxin-induced uveitis and the time course of inflammatory agents in aqueous humor. Int Immunopharmacol. 2004;4(8):1059–1066. doi: 10.1016/j.intimp.2004.04.011. [DOI] [PubMed] [Google Scholar]
- 71.Shiratori K, Ohgami K, Ilieva IB, et al. Inhibition of endotoxin-induced uveitis and potentiation of cyclooxygenase-2 protein expression by alpha-melanocyte-stimulating hormone. Invest Ophthalmol Vis Sci. 2004;45(1):159–164. doi: 10.1167/iovs.03-0492. [DOI] [PubMed] [Google Scholar]
- 72.Lee DJ, Taylor AW. Both MC5r and A2Ar are required for protective regulatory immunity in the spleen of post-experimental autoimmune uveitis in mice. J Immunol. 2013;191(8):4103–4111. doi: 10.4049/jimmunol.1300182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lee DJ, Taylor AW. Recovery from experimental autoimmune uveitis promotes induction of antiuveitic inducible Tregs. J Leukoc Biol. 2015;97(6):1101–1109. doi: 10.1189/jlb.3A1014-466RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li D, Taylor AW. Diminishment of alpha-MSH anti-inflammatory activity in MC1r siRNA-transfected RAW264.7 macrophages. J Leukoc Biol. 2008;84(1):191–198. doi: 10.1189/jlb.0707463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Getting SJ, Christian HC, Lam CW, et al. Redundancy of a functional melanocortin 1 receptor in the anti-inflammatory actions of melanocortin peptides: studies in the recessive yellow (e/e) mouse suggest an important role for melanocortin 3 receptor. J Immunol. 2003;170(6):3323–3330. doi: 10.4049/jimmunol.170.6.3323. [DOI] [PubMed] [Google Scholar]
- 76.Neumann Andersen G, Nagaeva O, Mandrika I, et al. MC (1) receptors are constitutively expressed on leucocyte subpopulations with antigen presenting and cytotoxic functions. Clin Exp Immunol. 2001;126(3):441–446. doi: 10.1046/j.1365-2249.2001.01604.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Lee DJ, Taylor AW. Following EAU recovery there is an associated MC5r-dependent APC induction of regulatory immunity in the spleen. Invest Ophthalmol Vis Sci. 2011;52(12):8862–8867. doi: 10.1167/iovs.11-8153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Robertson MJ, Erwig LP, Liversidge J, et al. Retinal microenvironment controls resident and infiltrating macrophage function during uveoretinitis. Invest Ophthalmol Vis Sci. 2002;43(7):2250–2257. [PubMed] [Google Scholar]
- 79.Dick AD, Carter D, Robertson M, et al. Control of myeloid activity during retinal inflammation. J Leukoc Biol. 2003;74(2):161–166. doi: 10.1189/jlb.1102535. [DOI] [PubMed] [Google Scholar]
- 80.Phan TA, Taylor AW. The neuropeptides alpha-MSH and NPY modulate phagocytosis and phagolysosome activation in RAW 264.7 cells. J Neuroimmunol. 2013;260(1–2):9–16. doi: 10.1016/j.jneuroim.2013.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Taylor AW, Dixit S, Yu J. Retinal Pigment Epithelial Cell Line Suppression of Phagolysosome Activation. Int J Ophthalmol Eye Sci. 2015;(Suppl 2(1)):1–6. [PMC free article] [PubMed] [Google Scholar]
- 82.Taylor AW. Alpha-melanocyte stimulating hormone (alpha-MSH) is a post-caspase suppressor of apoptosis in RAW 264.7 macrophages. PLoS One. 2013;8(8):e74488. doi: 10.1371/journal.pone.0074488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Caspi RR, Roberge FG, Chan CC, et al. A new model of autoimmune disease. Experimental autoimmune uveoretinitis induced in mice with two different retinal antigens. J Immunol. 1988;140(5):1490–1495. [PubMed] [Google Scholar]
- 84.Kitaichi N, Namba K, Taylor A. Inducible immune regulation following autoimmune disease in the immune-privileged eye. Journal of Leukocyte Biology. 2005;77(4):496–502. doi: 10.1189/jlb.0204114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Taylor AW, Kitaichi N. The diminishment of experimental autoimmune encephalomyelitis (EAE) by neuropeptide alpha-melanocyte stimulating hormone (alpha-MSH) therapy. Brain Behav Immun. 2008;22(5):639–646. doi: 10.1016/j.bbi.2007.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Yin P, Luby TM, Chen H, et al. Generation of expression constructs that secrete bioactive alphaMSH and their use in the treatment of experimental autoimmune encephalomyelitis. Gene Ther. 2003;10(4):348–355. doi: 10.1038/sj.gt.3301902. [DOI] [PubMed] [Google Scholar]
- 87.Ng TF, Kitaichi N, Taylor AW. In vitro-generated autoimmune regulatory T cells enhance intravitreous allogeneic retinal graft survival. Investigative Ophthalmology & Visual Science. 2007;48(11):5112–5117. doi: 10.1167/iovs.07-0175. [DOI] [PubMed] [Google Scholar]
- 88.Cheng LB, Cheng L, Bi HE, et al. Alpha-melanocyte stimulating hormone protects retinal pigment epithelium cells from oxidative stress through activation of melanocortin 1 receptor-Akt-mTOR signaling. Biochem Biophys Res Commun. 2014;443(2):447–452. doi: 10.1016/j.bbrc.2013.11.113. [DOI] [PubMed] [Google Scholar]
- 89.Naveh N. Melanocortins applied intravitreally delay retinal dystrophy in Royal College of Surgeons rats. Graefes Arch Clin Exp Ophthalmol. 2003;241(12):1044–1050. doi: 10.1007/s00417-003-0781-y. [DOI] [PubMed] [Google Scholar]
- 90.Steffensen EH. Corticotropin, cortisone, and hydrocortisone in treatment of ocular disease. J Am Med Assoc. 1952;150(17):1660–1664. doi: 10.1001/jama.1952.03680170014004. [DOI] [PubMed] [Google Scholar]
- 91.Takakura A, Tessler HH, Goldstein DA, et al. Viral retinitis following intraocular or periocular corticosteroid administration: a case series and comprehensive review of the literature. Ocul Immunol Inflamm. 2014;22(3):175–182. doi: 10.3109/09273948.2013.866256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Bouzas EA, Karadimas P, Pournaras CJ. Central serous chorioretinopathy and glucocorticoids. Surv Ophthalmol. 2002;47(5):431–448. doi: 10.1016/s0039-6257(02)00338-7. [DOI] [PubMed] [Google Scholar]
- 93.Karadimas P, Bouzas EA. Glucocorticoid use represents a risk factor for central serous chorioretinopathy: a prospective, case-control study. Graefes Arch Clin Exp Ophthalmol. 2004;242(9):800–802. doi: 10.1007/s00417-004-0885-z. [DOI] [PubMed] [Google Scholar]
- 94.Wang M, Munch IC, Hasler PW, et al. Central serous chorioretinopathy. Acta Ophthalmol. 2008;86(2):126–145. doi: 10.1111/j.1600-0420.2007.00889.x. [DOI] [PubMed] [Google Scholar]
- 95.Cunningham ET, Jr, Wender JD. Practical approach to the use of corticosteroids in patients with uveitis. Can J Ophthalmol. 2010;45(4):352–358. doi: 10.3129/i10-081. [DOI] [PubMed] [Google Scholar]
- 96.McGhee CN, Dean S, Danesh-Meyer H. Locally administered ocular corticosteroids: benefits and risks. Drug Saf. 2002;25(1):33–55. doi: 10.2165/00002018-200225010-00004. [DOI] [PubMed] [Google Scholar]
- 97.Taylor AW. Ocular immunosuppressive microenvironment. Chem Immunol Allergy. 2007;92:71–85. doi: 10.1159/000099255. [DOI] [PubMed] [Google Scholar]
- 98.Vignali DA, Collison LW, Workman CJ. How regulatory T cells work. Nat Rev Immunol. 2008;8(7):523–532. doi: 10.1038/nri2343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Taylor AW, Streilein JW. Inhibition of antigen-stimulated effector T cells by human cerebrospinal fluid. Neuroimmunomodulation. 1996;3(2–3):112–118. doi: 10.1159/000097235. [DOI] [PubMed] [Google Scholar]
- 100.Wong KY, Rajora N, Boccoli G, et al. A potential mechanism of local anti-inflammatory action of alpha-melanocyte-stimulating hormone within the brain: modulation of tumor necrosis factor-alpha production by human astrocytic cells. Neuroimmunomodulation. 1997;4(1):37–41. doi: 10.1159/000097313. [DOI] [PubMed] [Google Scholar]
- 101.Bhardwaj RS, Schwarz A, Becher E, et al. Pro-opiomelanocortin-derived peptides induce IL-10 production in human monocytes. J Immunol. 1996;156(7):2517–2521. [PubMed] [Google Scholar]
- 102.Bohm M, Wolff I, Scholzen TE, et al. alpha-Melanocyte-stimulating hormone protects from ultraviolet radiation-induced apoptosis and DNA damage. J Biol Chem. 2005;280(7):5795–5802. doi: 10.1074/jbc.M406334200. [DOI] [PubMed] [Google Scholar]
- 103.Chai B, Li JY, Zhang W, et al. Melanocortin-4 receptor-mediated inhibition of apoptosis in immortalized hypothalamic neurons via mitogen-activated protein kinase. Peptides. 2006;27(11):2846–2857. doi: 10.1016/j.peptides.2006.05.005. [DOI] [PubMed] [Google Scholar]
- 104.Zhang L, Dong L, Liu X, et al. alpha-Melanocyte-stimulating hormone protects retinal vascular endothelial cells from oxidative stress and apoptosis in a rat model of diabetes. PLoS One. 2014;9(4):e93433. doi: 10.1371/journal.pone.0093433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Edling AE, Gomes D, Weeden T, et al. Immunosuppressive activity of a novel peptide analog of alpha-melanocyte stimulating hormone (alpha-MSH) in experimental autoimmune uveitis. J Neuroimmunol. 2011;236(1–2):1–9. doi: 10.1016/j.jneuroim.2011.04.015. [DOI] [PubMed] [Google Scholar]
- 106.Naveh N, Kaplan-Messas A, Marshall J. Mechanism related to reduction of intraocular pressure by melanocortins in rabbits. Br J Ophthalmol. 2000;84(12):1411–1414. doi: 10.1136/bjo.84.12.1411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Catania A. Neuroprotective actions of melanocortins: a therapeutic opportunity. Trends Neurosci. 2008;31(7):353–360. doi: 10.1016/j.tins.2008.04.002. [DOI] [PubMed] [Google Scholar]
- 108.Joosten EA, Majewska B, Houweling DA, et al. Alpha-melanocyte stimulating hormone promotes regrowth of injured axons in the adult rat spinal cord. J Neurotrauma. 1999;16(6):543–553. doi: 10.1089/neu.1999.16.543. [DOI] [PubMed] [Google Scholar]
- 109.Bharne AP, Upadhya MA, Kokare DM, et al. Effect of alpha-melanocyte stimulating hormone on locomotor recovery following spinal cord injury in mice: Role of serotonergic system. Neuropeptides. 2011;45(1):25–31. doi: 10.1016/j.npep.2010.10.001. [DOI] [PubMed] [Google Scholar]
- 110.Schaible EV, Steinstrasser A, Jahn-Eimermacher A, et al. Single administration of tripeptide alpha-MSH(11–13) attenuates brain damage by reduced inflammation and apoptosis after experimental traumatic brain injury in mice. PLoS One. 2013;8(8):e71056. doi: 10.1371/journal.pone.0071056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Giuliani D, Galantucci M, Neri L, et al. Melanocortins protect against brain damage and counteract cognitive decline in a transgenic mouse model of moderate Alzheimers disease. Eur J Pharmacol. 2014;740:144–150. doi: 10.1016/j.ejphar.2014.06.063. [DOI] [PubMed] [Google Scholar]
- 112.Giuliani D, Bitto A, Galantucci M, et al. Melanocortins protect against progression of Alzheimer’s disease in triple-transgenic mice by targeting multiple pathophysiological pathways. Neurobiol Aging. 2014;35(3):537–547. doi: 10.1016/j.neurobiolaging.2013.08.030. [DOI] [PubMed] [Google Scholar]
- 113.Ma K, McLaurin J. alpha-Melanocyte stimulating hormone prevents GABAergic neuronal loss and improves cognitive function in Alzheimer’s disease. J Neurosci. 2014;34(20):6736–6745. doi: 10.1523/JNEUROSCI.5075-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Fang J, Han D, Hong J, et al. SValpha-MSH, a novel alpha-melanocyte stimulating hormone analog, ameliorates autoimmune encephalomyelitis through inhibiting auto-reactive CD4(+) T cells activation. J Neuroimmunol. 2014;269(1–2):9–19. doi: 10.1016/j.jneuroim.2014.01.010. [DOI] [PubMed] [Google Scholar]
- 115.Diem R, Hobom M, Maier K, et al. Methylprednisolone increases neuronal apoptosis during autoimmune CNS inflammation by inhibition of an endogenous neuroprotective pathway. J Neurosci. 2003;23(18):6993–7000. doi: 10.1523/JNEUROSCI.23-18-06993.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Miller H, Newell DJ, Ridley A. Multiple sclerosis. Treatment of acute exacerbations with corticotrophin (A. C.T.H.) Lancet. 1961;2(7212):1120–1122. doi: 10.1016/s0140-6736(61)91030-3. [DOI] [PubMed] [Google Scholar]
- 117.Arnason BG, Berkovich R, Catania A, et al. Mechanisms of action of adrenocorticotropic hormone and other melanocortins relevant to the clinical management of patients with multiple sclerosis. Mult Scler. 2013;19(2):130–136. doi: 10.1177/1352458512458844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Weinstein RS. Clinical practice. Glucocorticoid-induced bone disease. N Engl J Med. 2011;365(1):62–70. doi: 10.1056/NEJMcp1012926. [DOI] [PubMed] [Google Scholar]
- 119.Besser GM, Butler PW, Plumpton FS. Adrenocorticotrophic action of long-acting tetracosactrin compared with corticotrophin-gel. Br Med J. 1967;4(5576):391–394. doi: 10.1136/bmj.4.5576.391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Coburg AJ, Gray SH, Katz FH, et al. Disappearance rates and immunosuppression of intermittent intravenously administered prednisolone in rabbits and human beings. Surg Gynecol Obstet. 1970;131(5):933–942. [PMC free article] [PubMed] [Google Scholar]
- 121.Zaidi M, Sun L, Robinson LJ, et al. ACTH protects against glucocorticoid-induced osteonecrosis of bone. Proc Natl Acad Sci U S A. 2010;107(19):8782–8787. doi: 10.1073/pnas.0912176107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Grieco P, Carotenuto A, Auriemma L, et al. Novel alpha-MSH peptide analogues with broad spectrum antimicrobial activity. PLoS One. 2013;8(4):e61614. doi: 10.1371/journal.pone.0061614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Singh M, Mukhopadhyay K. Alpha-melanocyte stimulating hormone: an emerging anti-inflammatory antimicrobial peptide. Biomed Res Int. 2014;2014:874610. doi: 10.1155/2014/874610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Holdeman M, Lipton JM. Effects of massive doses of alpha-MSH on thermoregulation in the rabbit. Brain Res Bull. 1985;14(4):327–330. doi: 10.1016/0361-9230(85)90193-5. [DOI] [PubMed] [Google Scholar]
- 125.Ugwu SO, Blanchard J, Dorr RT, et al. Skin pigmentation and pharmacokinetics of melanotan-I in humans. Biopharm Drug Dispos. 1997;18(3):259–269. doi: 10.1002/(sici)1099-081x(199704)18:3<259::aid-bdd20>3.0.co;2-x. [DOI] [PubMed] [Google Scholar]
- 126.Dorr RT, Lines R, Levine N, et al. Evaluation of melanotan-II, a superpotent cyclic melanotropic peptide in a pilot phase-I clinical study. Life Sci. 1996;58(20):1777–1784. doi: 10.1016/0024-3205(96)00160-9. [DOI] [PubMed] [Google Scholar]
- 127.Dorr RT, Dvorakova K, Brooks C, et al. Increased eumelanin expression and tanning is induced by a superpotent melanotropin [Nle4-D-Phe7]-alpha-MSH in humans. Photochem Photobiol. 2000;72(4):526–532. doi: 10.1562/0031-8655(2000)072<0526:ieeati>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 128.Dorr RT, Ertl G, Levine N, et al. Effects of a superpotent melanotropic peptide in combination with solar UV radiation on tanning of the skin in human volunteers. Arch Dermatol. 2004;140(7):827–835. doi: 10.1001/archderm.140.7.827. [DOI] [PubMed] [Google Scholar]
- 129.Royalty JE, Konradsen G, Eskerod O, et al. Investigation of safety, tolerability, pharmacokinetics, and pharmacodynamics of single and multiple doses of a long-acting alpha-MSH analog in healthy overweight and obese subjects. J Clin Pharmacol. 2014;54(4):394–404. doi: 10.1002/jcph.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Safarinejad MR. Evaluation of the safety and efficacy of bremelanotide, a melanocortin receptor agonist, in female subjects with arousal disorder: a double-blind placebo-controlled, fixed dose, randomized study. J Sex Med. 2008;5(4):887–897. doi: 10.1111/j.1743-6109.2007.00698.x. [DOI] [PubMed] [Google Scholar]
- 131.Rosen RC, Diamond LE, Earle DC, et al. Evaluation of the safety, pharmacokinetics and pharmacodynamic effects of subcutaneously administered PT-141, a melanocortin receptor agonist, in healthy male subjects and in patients with an inadequate response to Viagra. Int J Impot Res. 2004;16(2):135–142. doi: 10.1038/sj.ijir.3901200. [DOI] [PubMed] [Google Scholar]
- 132.Ceriani G, Diaz J, Murphree S, et al. The neuropeptide alpha-melanocyte-stimulating hormone inhibits experimental arthritis in rats. Neuroimmunomodulation. 1994;1(1):28–32. doi: 10.1159/000097087. [DOI] [PubMed] [Google Scholar]
- 133.Botte DA, Noronha IL, Malheiros DM, et al. Alpha-melanocyte stimulating hormone ameliorates disease activity in an induced murine lupus-like model. Clin Exp Immunol. 2014;177(2):381–390. doi: 10.1111/cei.12336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Rajora N, Boccoli G, Catania A, et al. alpha-MSH modulates experimental inflammatory bowel disease. Peptides. 1997;18(3):381–385. doi: 10.1016/s0196-9781(96)00345-2. [DOI] [PubMed] [Google Scholar]
- 135.Fiechtner J, Montroy T. Treatment of moderately to severely active systemic lupus erythematosus with adrenocorticotropic hormone: a single-site, open-label trial. Lupus. 2014;23(9):905–912. doi: 10.1177/0961203314532562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Levine T. Treating refractory dermatomyositis or polymyositis with adrenocorticotropic hormone gel: a retrospective case series. Drug Des Devel Ther. 2012;6:133–139. doi: 10.2147/DDDT.S33110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Bomback AS, Tumlin JA, Baranski J, et al. Treatment of nephrotic syndrome with adrenocorticotropic hormone (ACTH) gel. Drug Des Devel Ther. 2011;5:147–153. doi: 10.2147/DDDT.S17521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Bomback AS, Canetta PA, Beck LH, Jr, et al. Treatment of resistant glomerular diseases with adrenocorticotropic hormone gel: a prospective trial. Am J Nephrol. 2012;36(1):58–67. doi: 10.1159/000339287. [DOI] [PubMed] [Google Scholar]
- 139.Baram TZ, Mitchell WG, Tournay A, et al. High-dose corticotropin (ACTH) versus prednisone for infantile spasms: a prospective, randomized, blinded study. Pediatrics. 1996;97(3):375–379. [PMC free article] [PubMed] [Google Scholar]