

Abstract
Efficient islet release from the pancreas requires the combination of collagenase, neutral protease (cNP), or thermolysin (TL). Recently, it has been shown that clostripain (CP) may also contribute to efficient islet release from the human pancreas. The aim of this study was to evaluate the impact of these proteases on human islet integrity in a prospective approach. Islets were isolated from the pancreas of 10 brain-dead human organ donors. Purified islets were precultured for 3 to 4 d at 37 °C to ensure that preparations were cleared of predamaged islets, and only integral islets were subjected to 90 min of incubation at 37 °C in Hank’s balanced salt solution supplemented with cNP, TL, or CP. The protease concentrations were calculated for a pancreas of 100 g trimmed weight utilizing 120 dimethyl-casein units of cNP, 70,000 caseinase units of TL, or 200 benzoyl-l-arginine-ethyl-ester units of CP (1×). These activities were then increased both 5× and 10×. After subsequent 24-h culture in enzyme-free culture medium, treated islets were assessed and normalized to sham-treated controls. Compared with controls and CP, islet yield was significantly reduced by using the 5× activity of cNP and TL, inducing also fragmentation and DNA release. Viability significantly decreased not until adding the 1× activity of cNP, 5× activity of TL, or 10× activity of CP. Although mitochondrial function was significantly lowered by 1× cNP and 5× TL, CP did not affect mitochondria at any concentration. cNP- and TL-incubated islets significantly lost intracellular insulin already at 1× activity, while the 10× activity of CP had to be added to observe a similar effect. cNP and TL have a similar toxic potency regarding islet integrity. CP also induces adverse effects on islets, but the toxic threshold is generally higher. We hypothesize that CP can serve as supplementary protease to minimize cNP or TL activity for efficient pancreas digestion.
Keywords: human islet isolation, human islet integrity, neutral protease, thermolysin, clostripain
Introduction
Over the last decade, human islet transplantation has become established as a successful procedure to treat life-threatening hypoglycemia unawareness in patients with type 1 diabetes unresponsive to conventional medical treatment.1,2 However, the variability and inconsistency of human islet isolation remain a challenge, as it is rarely possible to use a single donor pancreas per recipient to achieve islet graft longevity preventing islet transplantation from being made more widely available.3 Whereas this partly relates to the intrinsic variability of the human donor pancreas,4–7 the variability between different enzyme batches and also between different enzyme products significantly contributes to the inconsistency of human islet isolation outcome.8,9
The aim of enzymatic pancreas digestion is to ensure maximal liberation of morphologically intact islets from the surrounding acinar tissue. This requires the synergistic interaction between collagenases class I and class II and a supplementary neutral protease (NP) extracted from Clostridium histolyticum (cNP) or Bacillus thermoproteolyticus rokko (thermolysin [TL]) to degrade different collagen structures during pancreas digestion.10 Recently, it has also been shown that the tryptic-like activities from Clostridium histolyticum (clostripain [CP]) may also contribute to efficient islet release from the human pancreas.11,12 Therefore, the important debate about the ideal composition of enzyme blends for human pancreas digestion is still ongoing, with specific focus currently on the selection of the most effective supplementary type of neutral protease.13 Previous studies clearly demonstrated that neutral protease have an important function in the interaction between collagenases class I and class II14,15 and are particularly crucial for the integrity of isolated islets. Observations in rat and human pancreases indicated a dose-dependent detrimental effect of these enzymes on the morphological and functional viability of isolated islets.14,16–18
A large-scale study analyzing nearly 200 human islet isolations found that islets isolated with a TL-containing enzyme blend have a lower functional potency than islets isolated by means of clostridial cNP.19 The less harmful effect of cNP on human islet isolation outcome was confirmed by another comprehensive report when compared to supplementation with TL.13 Nevertheless, the multiplicity of factors that vary between the different enzyme blends assessed makes it difficult to draw firm conclusions from both studies.
To the best of our knowledge, no prospective study has been undertaken so far to directly compare the effect of different supplementary NPs on human islet integrity. The aim of the present study therefore was to evaluate the impact of different activities of cNP, TL, and CP on the viability and survival of isolated and precultured human islets.
Materials and Methods
Human Islet Isolation
Human pancreases with appropriate research consent and ethical approval were retrieved from 10 brain-dead human multiorgan donors. In situ cold perfusion was performed with University of Wisconsin solution (ViaSpan?, DuPont Pharmaceuticals Ltd., Hertfordshire, UK; n = 6) or with histidine–tryptophan–ketoglutarate (Custodiol, Köhler Chemie GmbH, Alsbach, Germany; n = 4). The donors (3 female/7 male) were characterized by an age of 58 ± 4 years, a body mass index of 24.5 ± 1.4, and a cold ischemia time of 13.1 ± 2.2 h.
After arrival in the central islet isolation facility in Uppsala, Sweden, pancreases were processed by the same isolation team using standard techniques as previously described in detail.20 This involved intraductal infusion of collagenase NB1 and cNP (Serva/Nordmark Arzneimittel GmbH & Co. KG, Uetersen, Germany) and density-gradient purification. Purified islets were then cultured for 3 to 4 d in normal air at 37 °C in bicarbonate-free Connaught Medical Research Laboratories (CMRL) 1066 supplemented with 25 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 1 mM pyruvate, 10 mM nicotinamide (PAA, Pasching, Austria), 10% fetal bovine serum, 2.5 mM l-glutamine, 100 U/µg/mL penicillin–streptomycin (Thermo Fisher, Stockholm, Sweden), and 20 µg/mL ciprofloxacin (Bayer, Leverkusen, Germany) to ensure that preparations were cleared of predamaged islets and only integral islets subjected to 90 min of protease incubation. Following purification and after islet culture, islet yield was determined in a standardized procedure converting islet yield to islet equivalents (IEQ) with an average diameter of 150 µm. Isolated islets were distinguished from exocrine tissue using insulin-specific dithizone staining (Sigma-Aldrich AB, Stockholm, Sweden).21
Islet Treatment
After culture, recovered islets were seeded as 500 IEQ-aliquots into 6-well plates equipped with 8 µm pore-sized inserts (Costar, Sigma-Aldrich) and incubated in 3 mL of bicarbonate-free Hank’s balanced salt solution (HBSS, PAA) supplemented with different activities of CP, cNP (Serva), or TL (Roche Diagnostics, Mannheim, Germany). All enzymes were provided in premium grade. The enzymatic activity added per islet aliquot (Table 1) was calculated on the basis of a human pancreas of 100 g trimmed weight which is perfused with a volume of 300 mL HBSS supplemented with 2,000 Wuensch units 4-Phenylazobenzyloxycarbonyl (PZ-U) of collagenase activity. Depending on the enzyme manufacturer’s recommendations or experience-based preferences, collagenase could be supplemented with 120 dimethyl-casein units of cNP,11 70,000 caseinase units of TL,22 and/or 200 benzoyl-l-arginine-ethyl-ester (BAEE) units of CP.11,12 These activities were additionally increased 5- and 10-fold to evaluate the toxic potency of supplementary proteases within a broad range (Table 1). Islets incubated in plain HBSS served as sham-treated controls. After 90 min of incubation at 37 °C in an incubator feeded with normal air as atmosphere, enzyme-treated islets were transferred to 6-well plates filled with 3 mL of fresh culture medium supplemented as described above and cultured for 24 h at 37 °C in normal air prior to islet characterization.
Table 1.
Protease Activities.
| Activity | n | Control (HBSS) | Clostripain (Benzoyl-l-Arginine- Ethyl-Ester-U/mL) | Neutral Protease (Dimethyl-Casein-U/mL) | Thermolysin (Caseinase-U/mL) |
|---|---|---|---|---|---|
| 1-Fold | 10 | — | 0.67 | 0.4 | 233 |
| 5-Fold | 10 | — | 3.35 | 2.0 | 1,167 |
| 10-Fold | 10 | — | 6.70 | 4.0 | 2,333 |
Islet Characterization
After enzyme treatment and subsequent 24-h culture as described above, remaining islets were harvested and quantified as IEQ. Islet morphological integrity was individually categorized by 3 examiners according to a semiquantitative fragmentation score from 0 (no fragmentation) to 3 (extensive fragmentation) using a blind design. After assessment, the average of the individual scores was calculated. Representative images for the different fragmentation scores are shown in Fig. 1A–D. Islet viability was measured using Syto-13 (Invitrogen-Molecular Probes, Stockholm, Sweden) and ethidium bromide (Sigma-Aldrich) for staining of viable and dead cells, respectively.23 Briefly, 30 to 40 islets were collected into a 2 mL microcentrifuge tube. After sedimentation in phosphate-buffered saline (PBS) by normal gravity, supernatant was removed and replaced by 10 µL of 25 µM Syto-13 and 10 µL of 50 µM ethidium bromide prior to incubation in the dark for 3 min. After duplicate washing with PBS, stained islets were resuspended in 200 µL of PBS and transferred as duplicate samples of 100 µL each in a black 96-well plate. The fluorescence of Syto-13 and ethidium bromide was quantified against PBS as background at 485/535 nm and 510/595 nm, respectively, using a fluorometric plate reader.
Fig. 1.
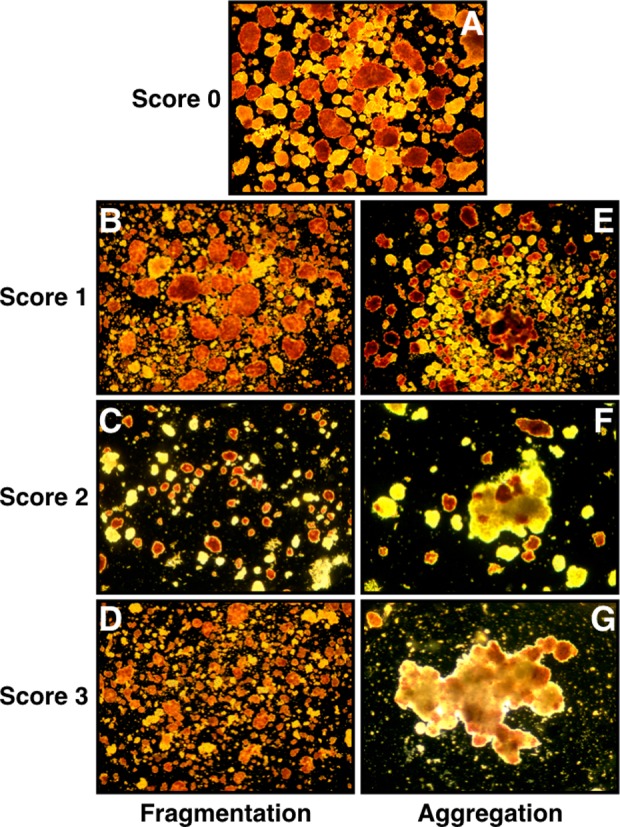
Representative images for scoring islet morphology with respect to fragmentation (left panel, B, C, and D) and aggregation (right panel, E, F, and G). Nonaffected islets are shown in Fig. 1A.
After sonification in acid ethanol, the intracellular insulin content of recovered islets was measured utilizing an ELISA kit specific for human insulin (Mercodia, Uppsala, Sweden) and normalized to islet DNA content determined by means of a fluorometric assay (Quant-iT, Invitrogen-Molecular Probes).24 Mitochondrial activity was evaluated by measuring the conversion of the tetrazolium compound 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS, Promega, Mannheim, Germany) into formazan as previously described.25 Briefly, aliquots of 100 islets were collected in triplicate for each experimental group in 200 µL of culture medium supplemented as described above. After incubation at 37 °C for 3 h, formazan formation was determined in 150 µL of supernatant at 490 nm and expressed as optical density per microgram DNA. DNA was measured in MTS-treated islets as described above except that the DNA standard samples were stained with the same proportion of MTS as islet samples.
Release of DNA strands was blindly quantified by 3 examiners using a semiquantitative islet aggregation score from 0 (no aggregation) to 3 (extensive aggregation). After assessment, the average of the individual scores was calculated. Representative images for the different aggregation scores are shown in Fig. 1A and E–G. Islet overall survival was calculated by simultaneously considering the recovery of initially incubated IEQ and the proportion of viable islet cells.
Data Analysis
All statistical analysis was performed utilizing Prism 6.0h for MacIntosh (GraphPad, La Jolla, CA, USA). Analysis of data was carried out by the nonparametric Friedman test followed by Dunn’s test for multiple comparison. Differences were considered significant at P less than 0.05. P values more than 0.05 are termed nonsignificant (NS). Data are normalized to control islets incubated in plain HBSS. For clarity, results are expressed as mean ± standard error rather than the correct nonparametric measures of median and quartiles.
Results
Islet Yield and Fragmentation
Treatment with 1-fold enzyme activity revealed the highest recovery after incubation with CP, reaching nearly the same islet recovery than sham-treated controls (Table 2). All 3 proteases induced islet loss when using the 5-fold (P < 0.05) and 10-fold activity (P < 0.001). Differences between CP, cNP, and TL became significant at the 5-fold level (P < 0.05) but were diminished when enzymes were used at 10-fold activity (NS).
Table 2.
Recovery of Initially Incubated Islet Equivalents (%).
| Activity | n | Control | Clostripain | Neutral Protease | Thermolysin |
|---|---|---|---|---|---|
| 1-Fold | 10 | 89.0 ± 4.8 | 88.1 ± 6.4 | 79.4 ± 5.9 | 77.1 ± 7.2 |
| 5-Fold | 10 | — | 72.3 ± 5.5* | 65.8 ± 5.8a,*, † | 64.2 ± 7.2b,*, † |
| 10-Fold | 10 | — | 61.4 ± 5.1b,*** | 46.8 ± 5.8b,*** | 49.9 ± 5.8b,*** |
a P < 0.01, b P < 0.001 versus control.
*P < 0.05, ***P < 0.001 versus 1-fold.
† P < 0.05 versus clostripain.
Islet fragmentation was assessed by means of a semiquantitative scoring system. Representative images for the different fragmentation scores are shown in Fig. 1A–D. This demonstrated that primarily islets incubated with cNP were affected with regard to morphological integrity. Islet fragmentation was noted using the 5-fold activity of cNP and reached statistical significance at the 10-fold activity of cNP (P < 0.001) and TL (P < 0.05) in comparison with sham-treated islets (Table 3). Compared with the 1-fold activity of the corresponding enzyme, the detrimental effect became significant when the 10-fold activity of cNP (P < 0.01) or TL (P < 0.05) was applied. Nevertheless, in comparison to CP, cNP induced significantly higher fragmentation at the 5-fold (P < 0.05) and 10-fold level (P < 0.01) while TL did not (NS). In contrast, CP was not harmful for islet morphological integrity even when the 10-fold activity was used.
Table 3.
Islet Fragmentation Score.
| Activity | n | Control (0–3) | Clostripain (0–3) | Neutral Protease (0–3) | Thermolysin (0–3) |
|---|---|---|---|---|---|
| 1-Fold | 10 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.10 ± 0.10 | 0.00 ± 0.00 |
| 5-Fold | 10 | — | 0.00 ± 0.00 | 0.70 ± 0.15† | 0.40 ± 0.16 |
| 10-Fold | 10 | — | 0.20 ± 0.13 | 1.20 ± 0.13b,**, ‡ | 0.80 ± 0.13a,* |
a P < 0.05, b P < 0.001 versus control.
*P < 0.05, **P < 0.01 versus 1-fold.
† P < 0.05, ‡ P < 0.01 versus clostripain.
Islet DNA Release
Islet DNA release was quantified using a semiquantitative aggregation score. Representative images for the different aggregation scores are shown in Fig. 1A, D, and E. Islet aggregation became most relevant in islets treated with cNP and TL. Although a 10-fold activity of CP (P < 0.05) was needed to cause a significant DNA release compared with HBSS-incubated islets, cNP and TL (P < 0.05) induced significant aggregation at the 5-fold level (Table 4). However, in all enzymes assessed, the 10-fold activity was significantly more harmful for islets when compared with the 1-fold activity of the corresponding enzyme. When the detrimental effect of different enzyme activity was compared with the corresponding CP level only cNP caused a statistically stronger aggregation at 5-fold activity (P < 0.05), while a 10-fold activity of TL was needed to detect a statistically significant difference to CP (P < 0.05; Table 4).
Table 4.
Islet Aggregation Score.
| Activity | n | Control (0–3) | Clostripain (0–3) | Neutral Protease (0–3) | Thermolysin (0–3) |
|---|---|---|---|---|---|
| 1-Fold | 10 | 0.00 ± 0.00 | 0.00 ± 0.00** | 0.80 ± 0.25** | 0.40 ± 0.22*** |
| 5-Fold | 10 | 0.20 ± 0.13* | 1.40 ± 0.27a, † | 1.40 ± 0.40a | |
| 10-Fold | 10 | 1.10 ± 0.18a | 2.40 ± 0.22b, † | 2.20 ± 0.29b, † |
a P < 0.05, b P < 0.001 versus control.
*P < 0.05, **P < 0.01, ***P < 0.001 versus 10-fold.
† P < 0.05 versus clostripain.
Islet Viability
The differences in islet viability were relatively small but were nevertheless significant when using Syto-13 and ethidium bromide in an unbiased plate reader assay. Compared with sham-treated islets, a significantly adverse effect on viability was observed in islets incubated with cNP (P < 0.01) already at the 1-fold level, while islets treated with TL were characterized by a decreased viability using the 5-fold activity (P < 0.001; Table 5). CP-treated islets were not affected until a 10-fold enzyme activity was added (P < 0.05). Assessment of viability within the experimental groups showed that even the 10-fold activity of cNP did not significantly alter the viability in comparison to the corresponding 1-fold level. The discrete reduction in viability within the other experimental groups became significant using the 10-fold activity of cNP (P < 0.05) or TL (P < 0.001; Table 5). Comparison of CP with the other proteases at the same activity level revealed that cNP (P < 0.05) was significantly more noxious already at 1-fold activity (P < 0.05) while TL (P < 0.05) was characterized by a significantly adverse effect on viability not until adding the 5-fold activity (Table 5).
Table 5.
Islet Viability (%).
| Activity | n | Control | Clostripain | Neutral Protease | Thermolysin |
|---|---|---|---|---|---|
| 1-Fold | 10 | 72.4 ± 3.1 (100%) | 69.6 ± 3.1 (96.2 ± 0.7%) | 66.1 ± 3.4b (91.1 ± 1.3%)† | 67.4 ± 3.6 (92.8 ± 1.7%) |
| 5-Fold | 10 | — | 68.4 ± 3.5 (94.2 ± 1.1%) | 64.8 ± 3.8c (88.9 ± 1.8%)† | 64.3 ± 3.7c (88.5 ± 2.0%)† |
| 10-Fold | 10 | — | 67.8 ± 3.6a (93.3 ± 1.4%) | 63.4 ± 4.2c,* (86.7 ± 2.4%)† | 62.2 ± 3.7c,*** (85.5 ± 2.0%)‡ |
Note. Normalized data are shown in brackets.
a P < 0.05, b P < 0.01, c P < 0.001 versus control.
*P < 0.05, ***P < 0.001 versus 1-fold.
† P < 0.05, ‡ P < 0.001 versus clostripain.
Mitochondrial Function
The mitochondrial function correlated with morphological viability. In comparison to sham-treated controls, the mitochondrial function of CP-treated islets was not reduced by any of the activities used (Table 6). In contrast, cNP significantly altered mitochondrial function already at the 1-fold level when compared with sham-treated islets (P < 0.01). Moreover, cNP was significantly more reductive for mitochondrial activity at all 3 levels when compared with corresponding CP-treated islets (P < 0.05). The mitochondrial function of TL-treated islets was significantly decreased in comparison with sham-treated islets when the 5-fold activity was applied (P < 0.01). Adding the 10-fold activity of TL, the inhibitory effect on mitochondria was also significantly stronger when compared with CP (P < 0.05). Except CP, all proteases had a significantly larger adverse effect on mitochondrial function using the 10-fold activity when opposed to the 1-fold activity of the corresponding enzyme (P < 0.05; Table 6).
Table 6.
Mitochondrial Function (Optical Density/µg DNA).
| Activity | n | Control | Clostripain | Neutral Protease | Thermolysin |
|---|---|---|---|---|---|
| 1-Fold | 10 | 51.2 ± 20.1 (100%) | 43.4 ± 16.4 (92.0 ± 2.9%) | 44.1 ± 19.7a (83.8 ± 3.9%)† | 43.4 ± 19.9 (88.4 ± 6.4%) |
| 5-Fold | 10 | — | 41.9 ± 15.6 (88.5 ± 3.3%) | 40.7 ± 17.4b (76.7 ± 4.0%)† | 37.9 ± 15.8a (80.8 ± 4.9%) |
| 10-Fold | 10 | — | 44.3 ± 17.6 (88.1 ± 3.8%) | 37.7 ± 16.3b,* (71.5 ± 5.2%)† | 33.3 ± 12.2b,* (71.9 ± 4.2%)† |
Note. Normalized data are shown in brackets.
a P < 0.01, b P < 0.001 versus control.
*P < 0.05 versus 1-fold.
† P < 0.05 versus clostripain.
Intracellular Insulin Content
Compared with sham-treated islets incubated in plain HBSS, a reduction in the intracellular insulin content (µU/ng DNA) was noted in all experimental groups (Table 7). However, the protease-induced decrease in intracellular insulin reached statistical significance at different activity levels. Although the insulin content of CP-treated islets was significantly reduced only when a 10-fold activity was applied (P < 0.05 vs. control), islets treated with cNP (P < 0.05) or TL (P < 0.01) lost a significant proportion of insulin already using the 1-fold activity. At this level, cNP- (P < 0.05) and TL-incubated islets (P < 0.01) stored a significantly lower amount of insulin in comparison with CP-treated islets. No significant differences were observed using the 5- or 10-fold activity of enzymes.
Table 7.
Intracellular Insulin Content (µU/ng DNA).
| Activity | n | Control | Clostripain | Neutral Protease | Thermolysin |
|---|---|---|---|---|---|
| 1-Fold | 10 | 150.2 ± 22.3 (100%) | 128.6 ± 21.7 (90.2 ± 11.3%) | 91.9 ± 17.5a (66.2 ± 11.7%)† | 86.7 ± 21.8b (54.3 ± 8.3%)‡ |
| 5-Fold | 10 | — | 99.6 ± 19.8* (70.3 ± 12.2%) | 78.4 ± 14.2b (59.7 ± 9.8%) | 98.2 ± 20.3 (64.5 ± 8.9%) |
| 10-Fold | 10 | — | 91.2 ± 16.6a,* (64.0 ± 8.7%) | 93.1 ± 15.0a (66.8 ± 8.8%) | 75.1 ± 13.1b (51.7 ± 6.0%) |
Note. Normalized data are shown in brackets.
a P < 0.05, b P < 0.01 versus control.
*P < 0.05, **P < 0.01 versus 1-fold.
† P < 0.05, ‡ P < 0.01 versus clostripain.
Islet Overall Survival
Compared with sham-treated controls and with the 1-fold level, a significantly negative effect of CP on islet overall survival was not observed until the 10-fold activity was applied (P < 0.001; Table 8). In contrast, the difference to controls (P < 0.001) and to the corresponding 1-fold level (P < 0.05) reached statistical significance using the 5-fold activity of cNP and TL. Compared with the corresponding concentration of CP, the harmful effect of TL and cNP on islet overall survival was significantly larger at the respectively 1-fold (P < 0.05) and 5-fold level (P < 0.05), but not when a 10-fold activity was used.
Table 8.
Islet Overall Survival (%).
| Activity | n | Control | Clostripain | Neutral Protease | Thermolysin |
|---|---|---|---|---|---|
| 1-Fold | 10 | 65.2 ± 4.9 | 62.2 ± 5.6 | 53.7 ± 5.7 | 52.0 ± 5.7† |
| 5-Fold | 10 | — | 49.9 ± 4.4 | 43.0 ± 4.4a,*, † | 40.7 ± 4.7a,*, † |
| 10-Fold | 10 | — | 42.2 ± 4.4a,*** | 29.2 ± 3.7a,*** | 30.9 ± 4.3a,*** |
a P < 0.001 versus control.
*P < 0.05, ***P < 0.001 versus 1-fold.
† P < 0.05, ‡ P < 0.01 versus clostripain.
Discussion
The multiplicity of factors that affect human islet isolation outcome makes it difficult to conclusively quantify the influence of individual enzymes on pancreas digestion. Although it is feasible to control for variables when pancreases from standardized rodents are studied,17,26 a prospective approach with human donors would require numerous pancreases to obtain statistically relevant data, something that is just not achievable with the shortage of suitable human donor pancreases required for clinical whole pancreas and islet transplant programs.27,28
To minimize the influence of donor-related and enzyme batch-dependent variables on the outcome of our experiments, we investigated the effect of different supplementary proteases on already isolated islets which enables the direct comparison between standardized samples. To diminish the effect of the enzyme blends used during the initial pancreas digestion, purified islets were precultured for 3 to 4 d at 37 °C to ensure that preparations were cleared of predamaged islets and only integral islets used.29–32 In order to identify a potentially toxic threshold, different levels of enzyme activities were applied. The calculation of these levels was based on previous experiences18,19 or on specifications defined during participation in the international trial of the Edmonton protocol.22
The few retrospective comparisons that had previously been performed on human pancreases had indicated that TL-containing blends may have a negative impact on islet integrity when compared with blends supplemented with cNP.13,19 A similar observation was made during rodent islet isolation demonstrating that islets isolated by means of liberase and TL fail to restore normoglycemia in isologous recipients when compared with islets released by means of collagenase P.33 However, the complexity of different enzyme isoforms influencing the efficiency of enzyme blends prevents a clear conclusion from these studies.34,35 Nevertheless, as a consequence of its potential harmfulness, TL was replaced by dispase in the manufacture of Liberase PI in order to improve islet isolation outcome from the pig pancreas.36 In contrast, our study surprisingly indicates that the adverse effect of cNP from Clostridium histolyticum on islet survival, morphological integrity, and viability appears to be equivalent or even stronger in comparison to TL. It is worth mentioning that those detrimental effects were observed at concentrations that are within the range of commonly used protease activities.37–39
Previous findings in rodent, porcine, and human islet isolation suggested that CP may serve as an alternative to cNP and TL. These studies demonstrated that the use of CP as additional supplement improves islet isolation outcome with respect to digestion time, yield, viability, and transplantability of islets.11,12,40 However, it cannot be excluded that these observations are related to a synergism between CP and other proteases. The already large extent of our study did not allow to realize an even more comprehensive experimental setting by additionally exposing human islets to protease combinations of CP plus cNP or TL which would add significant information. An increase in islet fragmentation but not of islet yield was noted when TL was combined with CP to digest rat pancreases. Opposite findings were made when CP was combined with cNP resulting in a significantly increased yield of nonfragmented rat islets.41 It has to be discussed whether the observations of Dendo et al. are related to an incompatibility between CP and TL, which are produced by different species, or whether the deteriorated islet morphology is secondary to the increased activation of endogenous proteases by TL plus CP.41 So far, no study has been completed to evaluate whether CP can serve as sole supplement for collagenase to replace cNP or TL as supplementary proteases. Pilot experiments performed in our lab nevertheless indicate that it is possible to successfully isolate islets from human pancreases of different sizes using 200 to 300 BAEE-U of CP as only supplementary activity for collagenase. More experiments are needed to verify this observation in a separate study.
The reasons for the relatively low toxicity of CP in comparison to cNP and TL are unknown so far. It can be speculated that these proteases have different specificities to digest the extracellular matrix surrounding the islets. Collagen IV and laminin 511 are the most abundant extracellular matrix proteins in the basement membrane of human islets.42 Particularly laminin 511 seems to play an essential role for β cell survival and function in vitro.43–45 As recently shown, the islet basement membrane is degraded during human islet isolation affecting the expression of several extracellular matrix proteins, such as collagen IV and laminin, which completely disappeared during subsequent islet culture. The harmfulness of this process was demonstrated by the leakage of adenylate kinase revealing a damage of the plasma membranes and mitochondria as well.46 This is in agreement with our findings demonstrating a reduction in membrane integrity, as expressed by the Syto-13/ethidium bromide viability assay, and the increased aggregation score indicating the release of DNA, a phenomenon that is frequently observed when human pancreases are overdigested or predamaged by prolonged warm ischemia time. The loss of morphological integrity was additionally verified by the increase in islet fragmentation. In this context, it is quite likely that the disruption of basement membranes and plasma membranes is accompanied by leakage of intracellularly stored insulin. It cannot be ruled out that also mitochondrial structures are compromised by enzymes as suggested by the reduced activity of these organelles determined by the MTS assay. In previous basic experiments in mice, it had been demonstrated that microdissected islets express a higher metabolic activity associated with a higher insulin content compared to collagenase-isolated islets.47
Recent studies revealed that CP has a significantly lower affinity to digest laminin 511 in comparison with cNP and TL.41,48 As a consequence, the integrity of the basement membrane of human islets would be better preserved using CP instead of cNP or TL for human pancreas digestion providing a significant advantage for islet survival and function during culture and after transplantation.49–51 Nevertheless, the present data indicate that CP can induce adverse effects on various parameters of islet integrity and viability, but the toxic threshold appears to be generally higher compared with cNP and TL. In this perspective, we hypothesize that the effects of different NPs on islet metabolism are secondary to the degradation of several extracellular matrix proteins within the basement membrane essential to maintain islet morphological and functional integrity.
In summary, our study compares for the first time the potential toxicity and harmfulness of different supplementary proteases for human islets in a prospective approach. It was demonstrated that cNP and TL have a similar toxic potency with regard to islet survival, viability, and morphological integrity. The adverse effects were observed at concentrations that are within the range of commonly used protease activities. In contrast, much higher concentrations were needed to induce detrimental effects with CP. Future studies should aim to investigate whether CP can act as sole supplement for collagenase or whether it has to be combined with another supplementary protease.
Acknowledgment
The authors wish to thank Sana Asif and Karin Andersson for technical assistance.
Footnotes
Ethical Approval: This study was approved by our institutional review board.
Statement of Human and Animal Rights: In situ cold perfusion was performed on 10 brain-dead human multiorgan donors.
Statement of Informed Consent: Human pancreases were retrieved with appropriate research consent and ethical approval.
Declaration of Conflicting Interests: The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding: The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by grants from the Diabetes Wellness Network Sweden, the Swedish Medical Research Council (K2015-54X-12219-19-4), the Nordic Insulin Fund, the EFSD/Novo Nordisk grant, the Ernfors Family Fund, Barn Diabetes Fonden, the Swedish Diabetes Association, and the Juvenile Diabetes Foundation International.
References
- 1. Meyer C, Hering BJ, Grossmann R, Brandhorst H, Brandhorst D, Gerich J, Federlin K, Bretzel RG. Improved glucose counterregulation and autonomic symptoms after intraportal islet transplants alone in patients with long-standing type I diabetes mellitus. Transplantation. 1998;66(2):233–240. [DOI] [PubMed] [Google Scholar]
- 2. Ryan EA, Shandro T, Green K, Paty BW, Senior PA, Bigam D, Shapiro AM, Vantyghem MC. Assessment of the severity of hypoglycemia and glycemic lability in type 1 diabetic subjects undergoing islet transplantation. Diabetes. 2004;53(4):955–962. [DOI] [PubMed] [Google Scholar]
- 3. Shapiro AM, Ricordi C, Hering B. Edmonton’s islet success has indeed been replicated elsewhere. Lancet. 2003;362(9391):1242. [DOI] [PubMed] [Google Scholar]
- 4. Benhamou PY, Watt PC, Mullen Y, Ingles S, Watanabe Y, Nomura Y, Hober C, Miyamoto M, Kenmochi T, Passaro EP, et al. Human islet isolation in 104 consecutive cases. Factors affecting isolation success. Transplantation. 1994;57(12):1804–1810. [PubMed] [Google Scholar]
- 5. Lakey JR, Warnock GL, Rajotte RV, Suarez-Alamazor ME, Ao Z, Shapiro AM, Kneteman NM. Variables in organ donors that affect the recovery of human islets of Langerhans. Transplantation. 1996;61(7):1047–1053. [DOI] [PubMed] [Google Scholar]
- 6. O’Gorman D, Kin T, Murdoch T, Richer B, McGhee-Wilson D, Ryan EA, Shapiro JA, Lakey JR. The standardization of pancreatic donors for islet isolations. Transplantation. 2005;80(6):801–806. [DOI] [PubMed] [Google Scholar]
- 7. Niclauss N, Bosco D, Morel P, Demuylder-Mischler S, Brault C, Milliat-Guittard L, Colin C, Parnaud G, Muller YD, Giovannoni L, et al. Influence of donor age on islet isolation and transplantation outcome. Transplantation. 2011;91(3):360–366. [DOI] [PubMed] [Google Scholar]
- 8. Nano R, Clissi B, Melzi R, Calori G, Maffi P, Antonioli B, Marzorati S, Aldrighetti L, Freschi M, Grochowiecki T, et al. Islet isolation for allotransplantation: variables associated with successful islet yield and graft function. Diabetologia. 2005;48(5):906–912. [DOI] [PubMed] [Google Scholar]
- 9. Hanley SC, Paraskevas S, Rosenberg L. Donor and isolation variables predicting human islet isolation success. Transplantation. 2008;85(7):950–955. [DOI] [PubMed] [Google Scholar]
- 10. McCarthy RC, Breite AG, Green ML, Dwulet FE. Tissue dissociation enzymes for isolating human islets for transplantation: factors to consider in setting enzyme acceptance criteria. Transplantation. 2011;91(2):137–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Brandhorst H, Friberg A, Andersson HH, Felldin M, Foss A, Salmela K, Lundgren T, Tibell A, Tufveson G, Korsgren O, et al. The importance of tryptic-like activity in purified enzyme blends for efficient islet isolation. Transplantation. 2009;87(3):370–375. [DOI] [PubMed] [Google Scholar]
- 12. Ståhle M, Foss A, Gustafsson B, Lempinen M, Lundgren T, Rafael E, Tufveson G, Korsgren O, Friberg A. Clostripain, the missing link in the enzyme blend for efficient human islet isolation. Transplant Direct. 2015;1(5):e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Balamurugan AN, Loganathan G, Bellin MD, Wilhelm JJ, Harmon J, Anazawa T, Soltani SM, Radosevich DM, Yuasa T, Tiwari M, et al. A new enzyme mixture to increase the yield and transplant rate of autologous and allogeneic human islet products. Transplantation. 2012;93(7):693–702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Brandhorst H, Alt A, Huettler S, Raemsch-Guenther N, Kurfuerst M, Bretzel RG, Brandhorst D. The ratio between class II and class I collagenase determines the amount of neutral protease activity required for efficient islet release from the rat pancreas. Transplant Proc. 2005;37(1):215–216. [DOI] [PubMed] [Google Scholar]
- 15. Kin T, O’Gorman D, Zhai X, Pawlick R, Imes S, Senior P, Shapiro AM. Nonsimultaneous administration of pancreas dissociation enzymes during islet isolation. Transplantation. 2009;87(11):1700–1705. [DOI] [PubMed] [Google Scholar]
- 16. Wolters GH, Vos-Scheperkeuter GH, van Deijnen JH, van Schilfgaarde R. An analysis of the role of collagenase and protease in the enzymatic dissociation of the rat pancreas for islet isolation. Diabetologia. 1992;35(8):735–742. [DOI] [PubMed] [Google Scholar]
- 17. Bucher P, Bosco D, Mathe Z, Matthey-Doret D, Andres A, Kurfuerst M, Ramsch-Gunther N, Buhler L, Morel P, Berney T. Optimization of neutral protease to collagenase activity ratio for islet of Langerhans isolation. Transplant Proc. 2004;36(4):1145–1146. [DOI] [PubMed] [Google Scholar]
- 18. Brandhorst H, Brendel MD, Eckhard M, Bretzel RG, Brandhorst D. Influence of neutral protease activity on human islet isolation outcome. Transplant Proc. 2005;37(1):241–242. [DOI] [PubMed] [Google Scholar]
- 19. Brandhorst H, Friberg A, Nilsson B, Andersson HH, Felldin M, Foss A, Salmela K, Tibell A, Tufveson G, Korsgren O, et al. Large-scale comparison of Liberase HI and collagenase NB1 utilized for human islet isolation. Cell Transplant. 2010;19(1):3–8. [DOI] [PubMed] [Google Scholar]
- 20. Brandhorst H, Brandhorst D, Brendel MD, Hering BJ, Bretzel RG. Assessment of intracellular insulin content during all steps of human islet isolation procedure. Cell Transplant. 1998;7(5):489–495. [DOI] [PubMed] [Google Scholar]
- 21. Ricordi C, Gray DW, Hering BJ, Kaufman DB, Warnock GL, Kneteman NM, Lake SP, London NJ, Socci C, Alejandro R, et al. Islet isolation assessment in man and large animals. Acta Diabetol Lat. 1990;27(3):185–195. [DOI] [PubMed] [Google Scholar]
- 22. Shapiro AM, Ricordi C, Hering BJ, Auchincloss H, Lindblad R, Robertson RP, Secchi A, Brendel MD, Berney T, Brennan DC, et al. International trial of the Edmonton protocol for islet transplantation. N Engl J Med. 2006;355(13):1318–1330. [DOI] [PubMed] [Google Scholar]
- 23. Barnett MJ, McGhee-Wilson D, Shapiro AM, Lakey JR. Variation in human islet viability based on different membrane integrity stains. Cell Transplant. 2004;13(5):481–488. [DOI] [PubMed] [Google Scholar]
- 24. Brandhorst H, Asif S, Andersson K, Theisinger B, Andersson HH, Felldin M, Foss A, Salmela K, Tibell A, Tufveson G, et al. A new oxygen carrier for improved long-term storage of human pancreata before islet isolation. Transplantation. 2010;89(2):155–160. [DOI] [PubMed] [Google Scholar]
- 25. Brandhorst D, Iken M, Tanioka Y, Brendel MD, Bretzel RG, Brandhorst H. Influence of collagenase loading on long-term preservation of pig pancreas by the two-layer method for subsequent islet isolation. Transplantation. 2005;79(1):38–43. [DOI] [PubMed] [Google Scholar]
- 26. Brandhorst H, Raemsch-Guenther N, Raemsch C, Friedrich O, Huettler S, Kurfuerst M, Korsgren O, Brandhorst D. The ratio between collagenase class I and class II influences the efficient islet release from the rat pancreas. Transplantation. 2008;85(3):456–461. [DOI] [PubMed] [Google Scholar]
- 27. Hudson A, Bradbury L, Johnson R, Fuggle SV, Shaw JA, Casey JJ, Friend PJ, Watson CJ. The UK pancreas allocation scheme for whole organ and islet transplantation. Am J Transplant. 2015;15(9):2443–2455. [DOI] [PubMed] [Google Scholar]
- 28. Ekser B, Cooper DK, Tector AJ. The need for xenotransplantation as a source of organs and cells for clinical transplantation. Int J Surg. 2015;23(Pt B):199–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Rijkelijkhuizen JK, van der Burg MP, Tons A, Terpstra OT, Bouwman E. Pretransplant culture selects for high-quality porcine islets. Pancreas. 2006;32(4):403–407. [DOI] [PubMed] [Google Scholar]
- 30. Barton FB, Rickels MR, Alejandro R, Hering BJ, Wease S, Naziruddin B, Oberholzer J, Odorico JS, Garfinkel MR, Levy M, et al. Improvement in outcomes of clinical islet transplantation: 1999-2010. Diabetes Care. 2012;35(7):1436–1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Brandhorst D, Brandhorst H, Hering BJ, Bretzel RG. Long-term survival, morphology and in vitro function of isolated pig islets under different culture conditions. Transplantation. 1999;67(12):1533–1541. [DOI] [PubMed] [Google Scholar]
- 32. Ilieva A, Yuan S, Wang R, Duguid WP, Rosenberg L. The structural integrity of the islet in vitro: the effect of incubation temperature. Pancreas. 1999;19(3):297–303. [DOI] [PubMed] [Google Scholar]
- 33. Vargas F, Julian JF, Llamazares JF, Garcia-Cuyas F, Jimenez M, Pujol-Borrell R, Vives-Pi M. Engraftment of islets obtained by collagenase and Liberase in diabetic rats: a comparative study. Pancreas. 2001;23(4):406–413. [DOI] [PubMed] [Google Scholar]
- 34. Barnett MJ, Zhai X, LeGatt DF, Cheng SB, Shapiro AM, Lakey JR. Quantitative assessment of collagenase blends for human islet isolation. Transplantation. 2005;80(6):723–728. [DOI] [PubMed] [Google Scholar]
- 35. Yamamoto T, Ricordi C, Messinger S, Sakuma Y, Miki A, Rodriguez R, Haertter R, Khan A, Alejandro R, Ichii H. Deterioration and variability of highly purified collagenase blends used in clinical islet isolation. Transplantation. 2007;84(8):997–1002. [DOI] [PubMed] [Google Scholar]
- 36. Cavanagh TJ, Lakey JR, Dwulet F, Wright MJ, Wile K, Albertson T, Fetterhoff T. Improved pig islet yield and post-culture recovery using Liberase PI purified enzyme blend. Transplant Proc. 1998;30(2):367. [DOI] [PubMed] [Google Scholar]
- 37. Szot GL, Lee MR, Tavakol MM, Lang J, Dekovic F, Kerlan RK, Stock PG, Posselt AM. Successful clinical islet isolation using a GMP-manufactured collagenase and neutral protease. Transplantation. 2009;88(6):753–756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Balamurugan AN, Breite AG, Anazawa T, Loganathan G, Wilhelm JJ, Papas KK, Dwulet FE, McCarthy RC, Hering BJ. Successful human islet isolation and transplantation indicating the importance of class 1 collagenase and collagen degradation activity assay. Transplantation. 2010;89(8):954–961. [DOI] [PubMed] [Google Scholar]
- 39. Caballero-Corbalan J, Brandhorst H, Asif S, Korsgren O, Engelse M, de Koning E, Pattou F, Kerr-Conte J, Brandhorst D. Mammalian tissue-free Liberase: a new GMP-graded enzyme blend for human islet isolation. Transplantation. 2010;90(3):332–333. [DOI] [PubMed] [Google Scholar]
- 40. Klock G, Kowalski MB, Hering BJ, Eiden ME, Weidemann A, Langer S, Zimmermann U, Federlin K, Bretzel RG. Fractions from commercial collagenase preparations: use in enzymic isolation of the islets of Langerhans from porcine pancreas. Cell Transplant. 1996;5(5):543–551. [DOI] [PubMed] [Google Scholar]
- 41. Dendo M, Maeda H, Yamagata Y, Murayama K, Watanabe K, Imura T, Inagaki A, Igarashi Y, Katoh Y, Ebina M, et al. Synergistic effect of neutral protease and clostripain on rat pancreatic islet isolation. Transplantation. 2015;99(7):1349–1355. [DOI] [PubMed] [Google Scholar]
- 42. Virtanen I, Banerjee M, Palgi J, Korsgren O, Lukinius A, Thornell LE, Kikkawa Y, Sekiguchi K, Hukkanen M, Konttinen YT, et al. Blood vessels of human islets of Langerhans are surrounded by a double basement membrane. Diabetologia. 2008;51(7):1181–1191. [DOI] [PubMed] [Google Scholar]
- 43. Otonkoski T, Banerjee M, Korsgren O, Thornell LE, Virtanen I. Unique basement membrane structure of human pancreatic islets: implications for beta-cell growth and differentiation. Diabetes Obes Metab. 2008;10(Suppl 4):119–127. [DOI] [PubMed] [Google Scholar]
- 44. Banerjee M, Virtanen I, Palgi J, Korsgren O, Otonkoski T. Proliferation and plasticity of human beta cells on physiologically occurring laminin isoforms. Mol Cell Endocrinol. 2012;355(1):78–86. [DOI] [PubMed] [Google Scholar]
- 45. Daoud J, Petropavlovskaia M, Rosenberg L, Tabrizian M. The effect of extracellular matrix components on the preservation of human islet function in vitro. Biomaterials. 2010;31(7):1676–1682. [DOI] [PubMed] [Google Scholar]
- 46. Cross SE, Vaughan RH, Willcox AJ, McBride AJ, Abraham AA, Han B, Johnson JD, Maillard E, Bateman PA, Ramracheya RD, et al. Key matrix proteins within the pancreatic islet basement membrane are differentially digested during human islet isolation. Am J Transplant. 2017;17(2):451–461. [DOI] [PubMed] [Google Scholar]
- 47. Atkins T, Matty AJ. Metabolic viability of freehand microdissected and collagenase-isolated islets of Langerhans. J Endocrinol. 1970;46(2):17–18. [PubMed] [Google Scholar]
- 48. Brandhorst D, Brandhorst H, Johnson PRV. Enzyme development for human islet isolation: five decades of progress or stagnation? Rev Diabet Stud. 2017;14(1):22–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Wang RN, Rosenberg L. Maintenance of beta-cell function and survival following islet isolation requires re-establishment of the islet-matrix relationship. J Endocrinol. 1999;163(2):181–190. [DOI] [PubMed] [Google Scholar]
- 50. Kaido T, Yebra M, Cirulli V, Rhodes C, Diaferia G, Montgomery AM. Impact of defined matrix interactions on insulin production by cultured human beta-cells: effect on insulin content, secretion, and gene transcription. Diabetes. 2006;55(10):2723–2729. [DOI] [PubMed] [Google Scholar]
- 51. Irving-Rodgers HF, Choong FJ, Hummitzsch K, Parish CR, Rodgers RJ, Simeonovic CJ. Pancreatic islet basement membrane loss and remodeling after mouse islet isolation and transplantation: impact for allograft rejection. Cell Transplant. 2014;23(1):59–72. [DOI] [PubMed] [Google Scholar]