

Abstract
Background
Understanding the molecular mechanisms of nanomaterial interacting with cellular systems is important for appropriate risk assessment. The identification of early biomarkers for potential (sub-)chronic effects of nanoparticles provides a promising approach towards cost-intensive and animal consuming long-term studies. As part of a 90-day inhalation toxicity study with CeO2 NM-212 and BaSO4 NM-220 the present investigations on gene expression and immunohistochemistry should reveal details on underlying mechanisms of pulmonary effects. The role of alveolar epithelial cells type II (AEII cells) is focused since its contribution to defense against inhaled particles and potentially resulting adverse effects is assumed. Low dose levels should help to specify particle-related events, including inflammation and oxidative stress.
Results
Rats were exposed to clean air, 0.1, 0.3, 1.0, and 3.0 mg/m3 CeO2 NM-212 or 50.0 mg/m3 BaSO4 NM-220 and the expression of 391 genes was analyzed in AEII cells after one, 28 and 90 days exposure. A total number of 34 genes was regulated, most of them related to inflammatory mediators. Marked changes in gene expression were measured for Ccl2, Ccl7, Ccl17, Ccl22, Ccl3, Ccl4, Il-1α, Il-1ß, and Il-1rn (inflammation), Lpo and Noxo1 (oxidative stress), and Mmp12 (inflammation/lung cancer). Genes related to genotoxicity and apoptosis did not display marked regulation. Although gene expression was less affected by BaSO4 compared to CeO2 the gene pattern showed great overlap. Gene expression was further analyzed in liver and kidney tissue showing inflammatory responses in both organs and marked downregulation of oxidative stress related genes in the kidney. Increases in the amount of Ce were measured in liver but not in kidney tissue. Investigation of selected genes on protein level revealed increased Ccl2 in bronchoalveolar lavage of exposed animals and increased Lpo and Mmp12 in the alveolar epithelia.
Conclusion
AEII cells contribute to CeO2 nanoparticle caused inflammatory and oxidative stress reactions in the respiratory tract by the release of related mediators. Effects of BaSO4 exposure are low. However, overlap between both substances were detected and support identification of potential early biomarkers for nanoparticle effects on the respiratory system. Signs for long-term effects need to be further evaluated by comparison to a respective exposure setting.
Electronic supplementary material
The online version of this article (10.1186/s12951-018-0343-4) contains supplementary material, which is available to authorized users.
Keywords: Nanoparticles, Gene expression, CeO2, BaSO4, Inhalation, Biomarkers, Alveolar epithelial cells type II, Inflammation, Oxidative stress
Background
Nanotechnologies and -materials are of great interest for product development and since several years its application is getting more popular. For safe use by manufacturers and consumers assessment of toxicity and adverse health effects is required. The present investigations are part of a comprehensive project on the toxicity and carcinogenicity of nanoparticles with nano-CeO2 as representative material in different concentrations covering the low to moderate dose range (0.1, 0.3, 1.0, 3.0 mg/m3), and nano-BaSO4 at one high concentration (50.0 mg/m3) for comparison to a non-toxic and safe material. The project is funded by the German Federal Ministry of Education and Research (03X0149) and should provide supporting data on a combined chronic inhalation toxicity and carcinogenicity study with cerium oxide and barium sulfate nanoparticles [NANoREG (81|0661/10|170)]. The automotive industry uses CeO2 nanoparticles in catalyzers and as fuel additive because of the material’s catalytic properties. Due to its chemical inertness, barium sulfate is used in multiple applications e.g. as filler in the biomedical sector. The nanoparticles were examined in a 90-day inhalation toxicity study with obligatory endpoints according to OECD TG 413 [1]. To identify early biomarkers for long-term effects of nanoparticles and investigate potential mechanisms of action, highly sensitive methods (gene expression analysis, immunohistochemistry) were further included in the project and are described in the present article.
Depending on their size and the striving to form agglomerates, nanoparticles penetrate different parts of the respiratory tract and likely end up in the alveolar space. They potentially react with cellular systems and induce inflammatory reactions or other molecular events. They further might pass the air–blood-barrier and translocate to extra-pulmonary organs. One major mechanism for the elimination of particles from the respiratory tract is the uptake by alveolar macrophages and clearance via the mucociliary escalator or the lymphatic system. However, the alveolar space consists of further cell types involved in host defense and lung function maintenance. Alveolar epithelial type II (AEII) cells are responsible for production and recycling of lung surfactant, play a role in turnover of the alveolar epithelia, and bear the ability to transform into alveolar epithelial type I (AEI) cells (e.g. for replacement of damaged cells). It has early been reviewed that AEII cells are an important component of the respiratory defense system against foreign material, including nanoparticles [2]. They produce and secrete a variety of factors, like surfactant proteins and chemokines to recruit macrophages and induce inflammatory processes for substance elimination [2–6]. It has been shown that carbon black nanoparticles, but not fine or nano-TiO2, or fine carbon black stimulate AEII cells to release factors for macrophage migration in vitro [7]. Furthermore, AEII cells and not macrophages are assumed to be the main producers of neutrophil attracting chemokines in the early inflammatory response to carbon nanoparticles administered via intratracheal instillation [8]. AEII cells might also internalize nanomaterial via different mechanisms, including the route of surfactant recycling [9–13]. In vivo studies on gold nanoparticles demonstrate the presence of the respective nanomaterial in lamellar bodies of AEII cells or the lung lining fluid [14, 15].
The presence of nanomaterial in the lung can cause chronic inflammatory reactions of lung tissue (e.g. after repeated administration), which can eventually result into long-term adverse effects like fibrosis or neoplastic lesions. It has recently been demonstrated that AEII cells play a role in CeO2 nanoparticle induced lung fibrosis [16]. Furthermore, it is known that AEII cells are a potential progenitor for lung tumors. As reviewed by Oberdörster [17] during particle-related inflammation, the recruited immune cells release growth factors and induce oxidative stress by the production of reactive oxygen species (ROS) for host defense. ROS can cause DNA damage, growth factors stimulate cell proliferation and by this the risk of tumor formation is increased. This hypothesis includes the well investigated process of secondary genotoxicity which is often accompanied by a present overload situation [18]. In contrast, little is known about primary mechanisms of genotoxicity, which are attributed to direct particle effects, like ROS generation due to surface reactivity [18]. Investigations of respective effects need to be done at absent inflammation. Less knowledge is published in how far CeO2 nanoparticles react via the described molecular mechanisms and if they bear a carcinogenic potential after inhalation. Several in vivo inhalation studies demonstrate the induction of inflammatory reactions [19–26]. However, the majority of these studies most likely describe effects occurring during a present overload situation. Pro-oxidative but also anti-oxidative effects have been demonstrated for CeO2 nanoparticles [19, 27–32]. Furthermore, some research indicates a genotoxic potential in vivo (intratracheal or oral application) [30, 33, 34] or in vitro [29, 35]. In contrast, first results from the chronic study mentioned earlier as well as the related dose-range finding study indicate no genotoxic effects for CeO2 [22, 36]. Investigations of Ma et al. [16, 37, 38] indicate fibrosis induction after CeO2 intratracheal instillation. More mechanistic information is needed for better understanding of partially contrary hypotheses. This includes the contribution of inflammation and overload on the one hand, and the particle’s reactivity on the other hand, to the respective molecular events. Also, the use of very high nanoparticle concentrations in most available studies requires more research on lower dose levels.
Since the major share of the inhaled nanoparticle dose usually deposits in the respiratory organs and is eliminated via mechanisms of pulmonary clearance, extra-pulmonary translocation of a smaller particle fraction is a frequently detected concomitant effect. Aalapati et al. [19] measured Ce contents in different extra-pulmonary organs after 28-day exposure in mice, with highest levels in liver and kidney and particle-related histopathological changes of the respective organs. Other in vivo studies display comparable effects for CeO2 nanoparticles [30, 39–43].
Potential mechanisms of action, especially regarding carcinogenicity of CeO2 and BaSO4 nanoparticles, were examined in this study. The highly sensitive method of gene expression analysis aims on the identification of early biomarkers for nanoparticle-related effects, especially those occurring after long-term exposure to realistic, low substance levels. The inclusion of low CeO2 nanoparticle concentrations should create a situation of absent overload and inflammation to identify substance-related pro-oxidative, pro-proliferative or apoptotic as well as genotoxic effects. We focused on AEII cells as they are potential key players in (particle induced) pulmonary toxicity. The contribution of extra-pulmonary translocation to nanoparticle toxicity mechanisms motivated additional examinations of liver and kidney tissue as organs responsible for substance elimination. The identification of early biomarkers after acute to subchronic nanoparticle exposure creates a step towards the reduction of cost-intensive, animal-consuming long-term in vivo studies. Moreover, the in vivo markers should serve as basis for further investigations in an in vitro nanoparticle-exposure setup to intensify the focus on alternatives to animal testing.
Results
Conventional endpoints according to OECD TG 413
The obligatory investigations of the 90-day inhalation toxicity study according to OECD TG 413 were comprehensively reported elsewhere [1] and are briefly summarized in Table 1. We detected increasing lung burden levels for both nanomaterials with reduced clearance halftimes of cerium at concentrations of ≥ 1.0 mg/m3. Contrastingly, barium was rapidly cleared from the lung. The response of lung tissue and associated cell populations was dominated by inflammatory reactions. Particle-laden macrophages and inflammatory cell infiltrations, directed by neutrophils were detected in histopathology and bronchoalveolar lavage (BAL) analysis. Immunohistochemistry markers for genotoxicity (CeO2 only) and cell proliferation were increased with ongoing exposure. Effects remained persistent up to 90 days subsequent to CeO2, and although to a much lower extent also after BaSO4 exposure.
Table 1.
Summary of conventional endpoints
| Investigation | CeO2 | BaSO4 |
|---|---|---|
| Retention analytics | ↑ lung burden (time- and concentration-dependent); ↓ clearance (≥ 1.0 mg/m3) | ↑ lung burden (time-dependent); rapid clearance |
| Hematology/clinical chemistry (d90 + 1rec) | ↑ blood neutrophils | NAD |
| BAL analysis | ↑ neutrophils, lymphocytes, total protein, LDH, β-glucuronidase (time-/concentration-dependent); post-exposure persistency | ↑ neutrophils; no post-exposure persistency |
| Histopathology | ↑ particle-laden macrophages and inflammatory cell infiltrations (alveolar/interstitial/lymphoid tissue), bronchiolo-alveolar hyperplasia and fibrosis; post-exposure persistency | ↑ particle-laden macrophages and inflammatory cell infiltrations; intra-epithelial eosinophilic globules and mucus cell hyperplasia in the nasal cavity; partly post-exposure persistency |
| Immunohistochemistry | ↑ γ-H2AX, 8-OHdG (genotoxicity); ↑ Ki-67 (proliferation); post-exposure persistency | ↑ Ki-67 (proliferation); partly post-exposure persistency |
NAD no abnormalities detected
Gene expression analysis in AEII cells
For AEII cells five different profiler PCR arrays (inflammatory cytokines and receptors, oxidative stress, DNA repair, apoptosis, lung cancer) were analyzed. In total, 34 genes were regulated and are listed in Table 2 and Additional file 1: Table S1. An overall upregulation of gene expression has been observed rather than downregulation. Switches between up- and downregulation of one specific gene has been detected rarely, therefore the direction of gene regulation could be determined quite explicit for most of the genes. Furthermore, the overlap between the regulated genes in response to both substances was quite high (> 60% similarity). The number of regulated genes increased with ongoing exposure time and increasing CeO2 concentration (Fig. 1). Analysis of gene distribution over the different pathways revealed major contribution of inflammatory mediators at all time points (Fig. 2). All other pathways yielded up to four regulated genes. Inflammatory- and oxidative stress-related gene numbers increased in response to CeO2. Also in response to BaSO4 a time-dependent increase of the number of regulated genes was detected. However, the distribution over the five specified groups of mediators did not increase significantly. In total, 30 different genes were affected by CeO2 and 25 genes by BaSO4 exposure. The difference was mainly due to a lower number of inflammatory mediators regulated by BaSO4. Data for the most promising genes regarding their function as potential marker for nanoparticle-related effects and linked mechanisms of action are displayed individually in the following sections. Figure 15 shows an overview of assumed mechanistic relationships between the different cell types and effects involved in CeO2 nanoparticle responses.
Table 2.
Gene regulation in AEII cells in response to CeO2 and BaSO4 after different exposure periods
| Array | Gene | CeO2 | BaSO4 | |||||
|---|---|---|---|---|---|---|---|---|
| 1 day | 28 days | 90 days | 1 day | 28 days | 90 days | |||
| Inflammatory cytokines and receptors | Ccl3 | C–C motif chemokine ligand 3 |
|
|
||||
| Ccl17 | C–C motif chemokine ligand 17 |
|
|
|||||
| Cx3cr1 | C–X3–C motif chemokine receptor 1 |
|
|
|||||
| Il1α | Interleukin 1 alpha |
|
|
|||||
| Il1β | Interleukin 1 beta |
|
|
|||||
| Il1rn | Interleukin 1 receptor antagonist |
|
|
|||||
| Tnfsf4 | Tumor necrosis factor superfamily member 4 |
|
||||||
| Ccl2 | C–C motif chemokine ligand 2 |
|
|
|
||||
| Ccl4 | C–C motif chemokine ligand 4 |
|
|
|
||||
| Ccl7 | C–C motif chemokine ligand 7 |
|
|
|||||
| Ccl11 | C–C motif chemokine ligand 11 |
|
|
|||||
| Ccl20 | C–C motif chemokine ligand 20 |
|
|
|
|
|
||
| Ccl22 | C–C motif chemokine ligand 22 |
|
|
|
|
|||
| Ccl24 | C–C motif chemokine ligand 24 |
|
|
|
|
|||
| Pf4 | platelet factor 4 |
|
|
|||||
| Cxcl9 | C–X–C motif chemokine ligand 9 |
|
|
|
||||
| Cd40lg | CD40 ligand |
|
|
|||||
| Il2rb | Interleukin 2 receptor, beta |
|
|
|||||
| Oxidative stress | Lpo | Lactoperoxidase |
|
|
|
|
||
| Noxo1 | NADPH oxidase organizer 1 |
|
|
|
||||
| Hba1 | Hemoglobin alpha, adult chain 2 |
|
|
|
|
|
||
| Scd1 | Stearoyl-Coenzyme A desaturase 1 |
|
|
|
||||
| Krt1 | Keratin 1 |
|
|
|||||
| DNA repair | Msh5 | mutS homolog 5 |
|
|||||
| Exo1 | Exonuclease 1 |
|
|
|
|
|
||
| Mutyh | MutY homolog (E. coli) |
|
|
|||||
| Apoptosis | Birc5 | Baculoviral IAP repeat-containing 5 |
|
|
|
|||
| Lung cancer | Cyp1b1 | Cytochrome P450, family 1, subfamily b, polypeptide 1 |
|
|||||
| Fabp4 | Fatty acid binding protein 4 |
|
|
|
||||
| Mmp12 | Matrix metallopeptidase 12 |
|
|
|
||||
| Opcml | Opioid binding protein/cell adhesion molecule-like |
|
|
|
|
|||
| Tcf21 | Transcription factor 21 |
|
||||||
| Thbs2 | Thrombospondin 2 |
|
||||||
| Top2a | Topoisomerase (DNA) II alpha |
|
||||||
| Total | 34 | 11 | 18 | 24 | 9 | 10 | 15 | |
 upregulation;
upregulation;
 downregulation;
downregulation;
 up- or downregulation (differences between CeO2 dose groups); cut-off: FR ≤ − 2.0 or ≥ 2.0
up- or downregulation (differences between CeO2 dose groups); cut-off: FR ≤ − 2.0 or ≥ 2.0
Fig. 1.
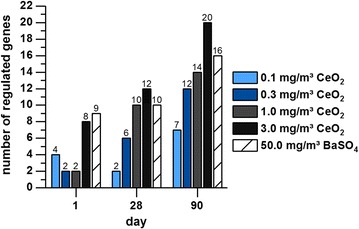
Number of regulated genes per concentration and time point. The number of regulated genes in AEII cells isolated from rats exposed to 0.1, 0.3, 1.0 or 3.0 mg/m3 CeO2 or 50.0 mg/m3 BaSO4 nanoparticles for one, 28 or 90 days is illustrated
Fig. 2.
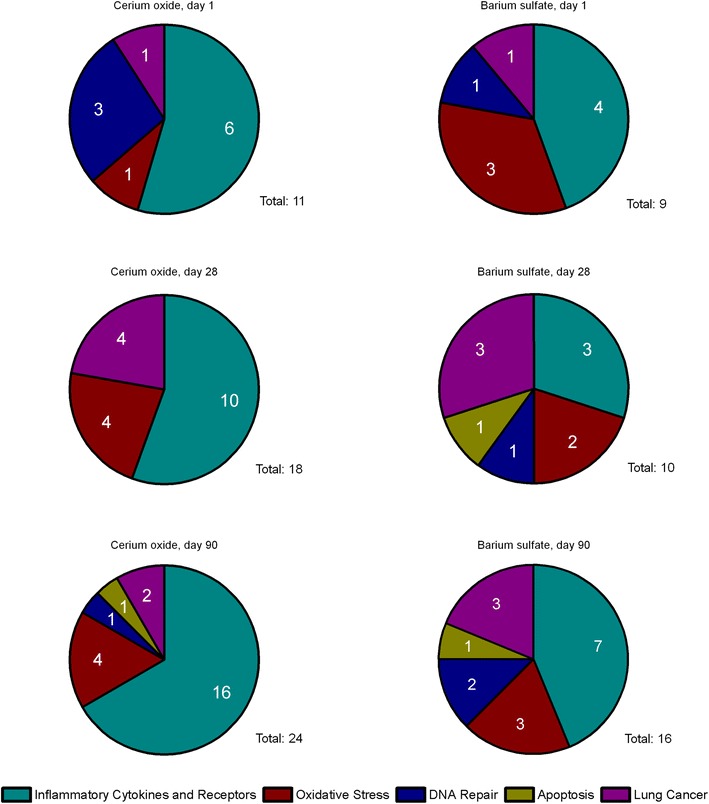
Distribution of regulated genes over the analyzed pathways. The number of regulated genes in isolated AEII cells after CeO2 nanoparticle (pool of all dose groups) and BaSO4 nanoparticle exposure is illustrated per time point and pathway
Fig. 15.
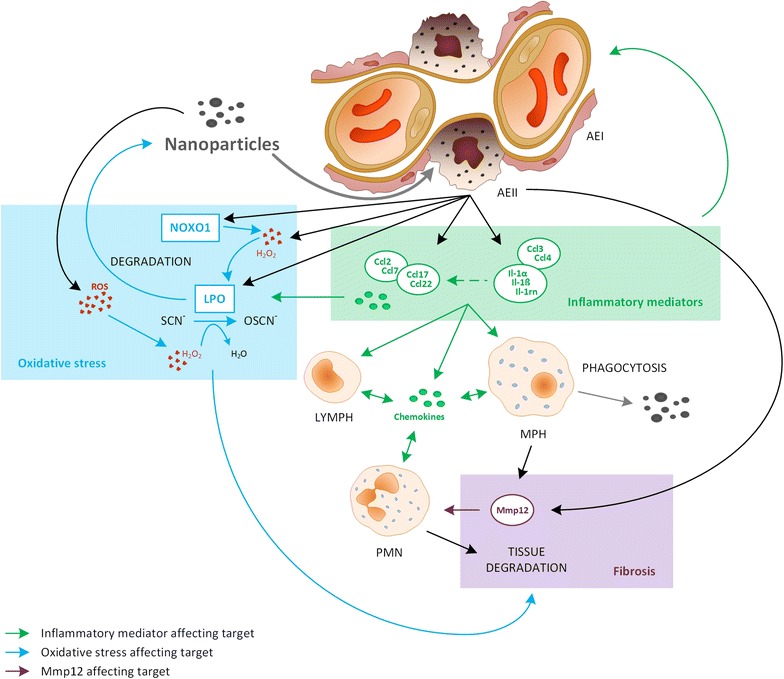
Potential mechanistic relationship between the mediators regulated in AEII cells after CeO2 nanoparticle exposure
Inflammatory cytokines and receptors
The chemokines Ccl2, Ccl7, Ccl17 and Ccl22 (Fig. 3a–d) showed the most distinct upregulation of all inflammatory mediators tested. Regulation was highest after 90 days exposure to 3.0 mg/m3 CeO2. Especially Ccl22 (Fig. 3d) showed a very high response to CeO2 with a gene expression up to 19-fold higher than the control group. The regulation pattern for these four chemokines is quite similar. The inflammatory mediators Ccl3, Ccl4, Ccl24, Il-1α, Il-1β, Il-1rn were upregulated at low CeO2 concentrations (0.1, 0.3 mg/m3) with a similar expression pattern (Fig. 3e–i). Although slight upregulation was detected for several mediators after 28 days, effects were highest after 90 days exposure. In contrast, BaSO4 only affected Ccl2, Ccl7, and Ccl22.
Fig. 3.
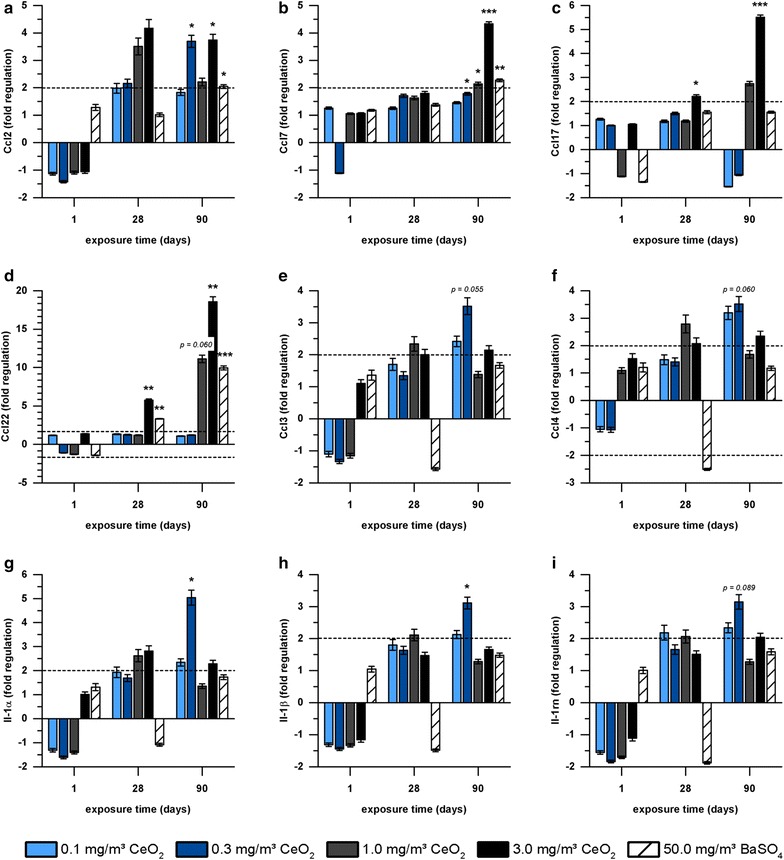
Gene expression of inflammatory mediators in AEII cells. Gene expression of a Ccl2, b Ccl7, c Ccl17, d Ccl22, e Ccl3, f Ccl4, g Il-1α, h Il-1ß, and i Il-1rn in AEII cells of rats exposed to 0.1, 0.3, 1.0 or 3.0 mg/m3 CeO2 or 50.0 mg/m3 BaSO4 nanoparticles for one, 28 or 90 days is illustrated. Values are expressed as mean fold regulation of clean air control ± SD; cut-off: fold regulation ≤ − 2.0 or ≥ 2.0 (dotted line), *p < 0.05, **p < 0.01, ***p < 0.001; n ≤ 5; Student’s T-test analysis of the replicate 2−∆Ct values for each gene in the control and treatment groups
Oxidative stress
Four genes related to oxidative stress were regulated after CeO2 nanoparticle exposure (Table 2). The highest responses were detected for Lpo and Noxo1 with a concentration- and time-dependent increase (Fig. 4). Expression levels exceeded the fold regulation cut off already at 0.3 mg/m3 CeO2 nanoparticle exposure and reached values up to 56-fold higher compared to clean air inhalation after 90 days exposure to 3.0 mg/m3 CeO2. Noxo1 was significantly upregulated after 28 and 90 days in response to the highest concentration of CeO2. BaSO4 exposure revealed a total number of five regulated genes related to oxidative stress (Table 2). Lpo was affected in a similar way as after CeO2 exposure, with a peak fold regulation of 25. Noxo1 was upregulated only after 28 days exposure to BaSO4.
Fig. 4.
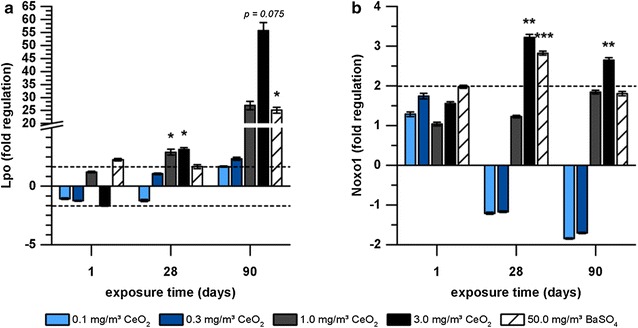
Expression of oxidative stress-related genes in AEII cells in response to nanoparticle exposure. Gene expression of a Lpo and b Noxo1 in AEII cells of rats exposed to 0.1, 0.3, 1.0 or 3.0 mg/m3 CeO2 or 50.0 mg/m3 BaSO4 nanoparticles for one, 28 or 90 days is illustrated. Values are expressed as mean fold regulation of clean air control ± SD; cut-off: fold regulation ≤ − 2.0 or ≥ 2.0 (dotted line), *p < 0.05, **p < 0.01, ***p < 0.001; n ≤ 5; Student’s T-test analysis of the replicate 2−∆Ct values for each gene in the control and treatment groups
DNA repair and apoptosis
Three genes related to DNA repair were regulated (Table 2), whereas Exo1 showed the most distinct regulation (data not shown). The impact of BaSO4 on Exo1 expression was higher than effects of CeO2. Birc5, an apoptosis-related gene was slightly upregulated after 90-day nanoparticle exposure (data not shown).
Lung cancer
The lung cancer pathway revealed seven regulated genes (Table 2). The strongest response was detected for Mmp12 (Fig. 5). Fold regulation values markedly increased with ongoing CeO2 nanoparticle exposure and increasing concentration. BaSO4 effects were lower, but still caused significant upregulation of Mmp12 after 90 days.
Fig. 5.
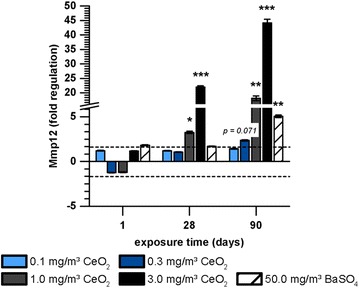
Gene expression of Mmp12 in AEII cells in response to nanoparticle exposure.Gene expression of Mmp12 in AEII cells of rats exposed to 0.1, 0.3, 1.0 or 3.0 mg/m3 CeO2 or 50.0 mg/m3 BaSO4 nanoparticles for one, 28 or 90 days is illustrated. Values are expressed as mean fold regulation of clean air control ± SD; cut-off: fold regulation ≤ − 2.0 or ≥ 2.0 (dotted line), *p < 0.05, **p < 0.01, ***p < 0.001; n ≤ 5; Student’s T-test analysis of the replicate 2−∆Ct values for each gene in the control and treatment groups
Analysis of protein expression in the alveolar compartment
For the pathways inflammation, oxidative stress and lung cancer, the following regulated genes were investigated regarding their protein expression in either BAL or lung tissue of CeO2 and BaSO4 exposed animals: Ccl2, Ccl20, Il-1α and Il-1ß (BAL analysis), Lpo and Mmp12 (immunohistochemistry of lung tissue).
Ccl2 levels in bronchoalveolar lavage
While Ccl20, IL-1α, and IL-1β protein levels in BAL did not change in response to nanoparticle exposure, Ccl2 was markedly increased with ongoing CeO2 exposure and increasing concentration (Fig. 6). In addition to gene expression analysis, mediator levels in BAL were also measured during post-exposure of up to 90 days. Although a slight decrease of Ccl2 levels was observed in this phase, elevated protein expression was overall persistent. BaSO4 also significantly affected Ccl2 protein expression in BAL.
Fig. 6.
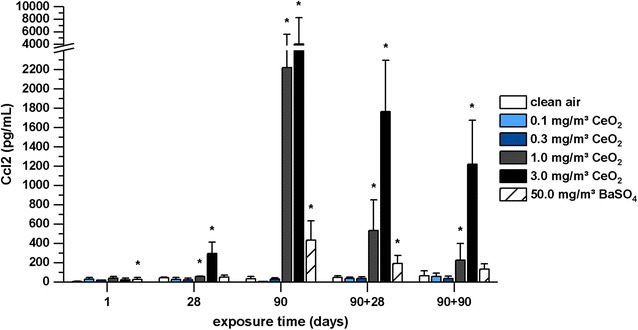
Ccl2 levels measured in bronchoalveolar lavage. Rats were exposed to clean air, 0.1, 0.3, 1.0, and 3.0 mg/m3 CeO2 nanoparticles or 50.0 mg/m3 BaSO4 nanoparticles. Ccl2 levels were determined after one, 28 and 90 days exposure and 28 and 90 days after the end of 90 days exposure (90 + 28, 90 + 90). Values are expressed as mean + SD, *p ≤ 0.05 vs. clean air control, n ≤ 5; Kruskal–Wallis–ANOVA with Mann–Whitney U-test as post hoc analysis
Immunohistochemical analysis of Lpo and Mmp12
Since gene expression of Lpo and Mmp12 was highly upregulated in AEII cells, immunohistochemistry was used to evaluate the development of their protein level. Figures 7 and 8 exemplarily show tissue sections of the alveolar region of rats exposed to clean air, 3.0 mg/m3 CeO2, or 50.0 mg/m3 BaSO4 for 90 days. The red colored signal indicates increased protein expression of Lpo (Fig. 7) or Mmp12 (Fig. 8) in response to both materials, while clean air did not cause any response. Lpo was detected in the alveolar epithelia in close proximity to the opening of the terminal bronchioli in regions of accumulated particle-laden macrophages and induced inflammation. The Lpo positive cells were morphologically consistent with AEII cells. Furthermore, no significant levels were measured in alveolar macrophages. This indicates that the increased Lpo gene expression in AEII cells caused also increased Lpo protein levels in the same cells. Lpo positive areas were additionally quantified per total tissue area. This analysis revealed an increasing signal with ongoing nanoparticle exposure, significant after 90 days for both substances (Fig. 9). The effect of CeO2 was higher compared to BaSO4 according to gene expression results. This is also visible in Fig. 7. The lung tissue overview sections contain more areas of CeO2 particle-laden macrophages with positive Lpo signal in epithelial cells (Fig. 7C) than BaSO4 related signals (Fig. 7E). As seen in Fig. 7D, the brown colored CeO2 particles accumulated in macrophages are histologically visible. The analyzed tissue was therefore further used for quantification of cerium (Fig. 10). The signal was significantly exceeding control levels and was increasing with ongoing particle exposure.
Fig. 7.
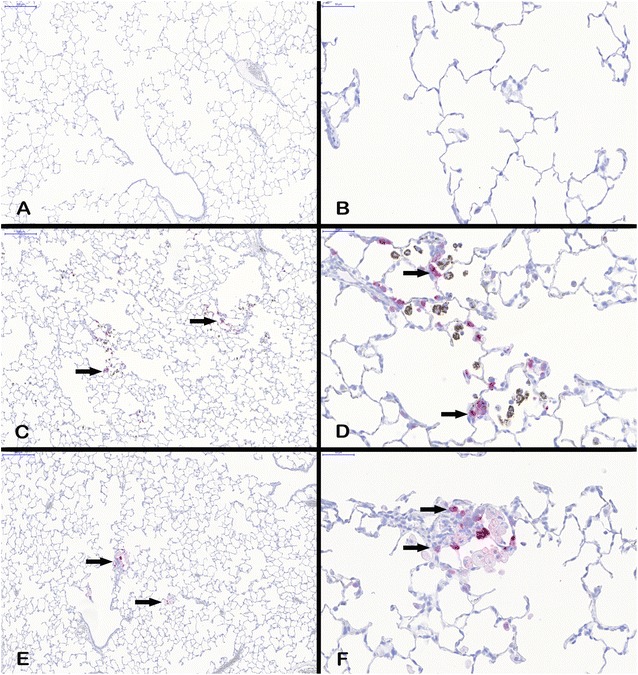
Lpo protein expression in lung tissue. All examples illustrate findings after 90-day nanoparticle exposure. A lung tissue overview, ×10, and B detailed view, ×40, after clean air exposure, C Lpo positive cells (arrows) in lung tissue overview, ×10, and D detailed view, ×40, after 3.0 mg/m3 CeO2 exposure, and E Lpo positive cells (arrows) in lung tissue overview, ×10, and F detailed view, ×40, after 50.0 mg/m3 BaSO4 exposure. The images show Lpo-positive cells counterstained with hematoxylin
Fig. 8.
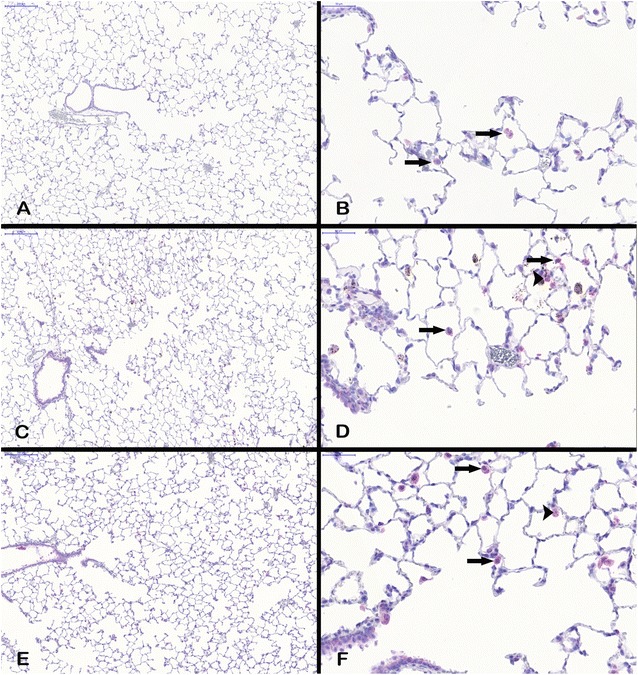
Mmp12 protein expression in lung tissue. All examples illustrate findings after 90-day nanoparticle exposure. A Mmp12 positive macrophages (arrows) in lung tissue overview (×10) and B detailed view (×40) after clean air exposure, C Mmp12 positive macrophages (arrows) and AEII cells (arrowhead) in lung tissue overview (×10) and D detailed view (×40) after 3.0 mg/m3 CeO2 exposure, and E Mmp12 positive macrophages (arrows) and AEII cells (arrowhead) in lung tissue overview (×10) and F detailed view (×40) after 50.0 mg/m3 BaSO4 exposure. The images show Mmp12-positive cells counterstained with hematoxylin
Fig. 9.
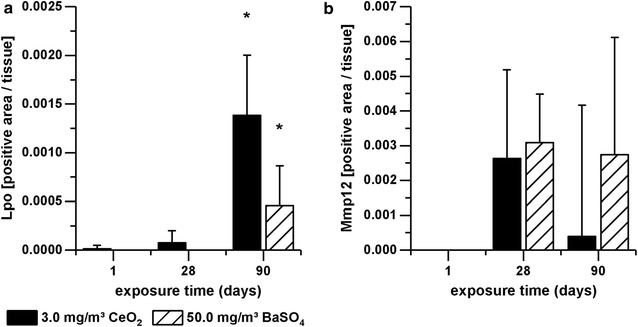
Lpo and Mmp12 levels in alveolar tissue. Rats were exposed to clean air, 3.0 mg/m3 CeO2 nanoparticles or 50.0 mg/m3 BaSO4 nanoparticles. a Lpo and b Mmp12 levels were determined immunohistochemically after one, 28 and 90 days exposure. Values are expressed as positive area per total tissue, normalized to the control group, mean + SD, *p < 0.05 vs. clean air control, n ≤ 5; Kruskal–Wallis–ANOVA with Mann–Whitney U-test as post hoc analysis
Fig. 10.
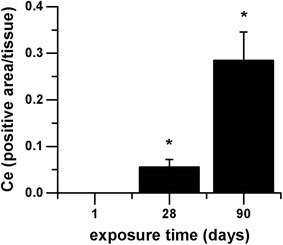
Cerium levels in alveolar tissue. Rats were exposed to clean air or 3.0 mg/m3 CeO2 nanoparticles. Cerium levels were determined in tissue slices after one, 28 and 90 days exposure. Values are expressed as positive area per total tissue, normalized to the control group, mean + SD, *p < 0.05 vs. clean air control, n ≤ 5; Kruskal–Wallis–ANOVA with Mann–Whitney U-test as post hoc analysis
Mmp12 protein levels were increased after 28 and 90 days nanoparticle exposure, but did not yield any significance, due to high variation between individual tissue slides (Fig. 9b). The picture of the lung sections (Fig. 8) indicates that the positive signal originates predominantly from alveolar macrophages rather than epithelial cells. However, single Mmp12 positive AEII cells were also detectable.
Gene expression analysis of liver and kidney tissue
In liver and kidney tissue 14 and 18 genes respectively were regulated in total (Table 3, Additional file 1: Table S1). Similar to the AEII cells a high response concerning the amount of regulated genes was seen for the endpoint inflammation (50% of total for liver, ~ 30% of total for kidney; Fig. 11). However, in kidney tissue even more genes were found to be related to oxidative stress, most of them downregulated. In both tissues a minor portion of the regulated genes was DNA repair-related. Comparison of both substances indicated a higher response of gene regulation to CeO2 than to BaSO4.
Table 3.
Gene regulation in liver and kidney in response to 90 days CeO2 and BaSO4 exposure
| Array | Gene | CeO2 | BaSO4 | |||
|---|---|---|---|---|---|---|
| Liver | Kidney | Liver | Kidney | |||
| Inflammatory cytokines and receptors | Ccr2 | Chemokine (C–C motif) receptor 2 |
|
|
||
| Cx3cl1 | Chemokine (C–X3–C motif) ligand 1 |
|
||||
| Cxcl12 | Chemokine (C–X–C motif) ligand 12 |
|
||||
| Cxcr2 | Chemokine (C–X–C motif) receptor 2 |
|
||||
| Cxcr3 | Chemokine (C–X–C motif) receptor 3 |
|
|
|||
| Il2rg | Interleukin 2 receptor, gamma |
|
||||
| Osm | Oncostatin M |
|
|
|||
| Ccl2 | Chemokine (C–C motif) ligand 2 |
|
||||
| Ccl9 | Chemokine (C–C motif) ligand 9 |
|
|
|||
| Cxcr5 | Chemokine (C–X–C motif) receptor 5 |
|
||||
| Spp1 | Secreted phosphoprotein 1 |
|
||||
| Oxidative stress | Gpx2 | Glutathione peroxidase 2 |
|
|
||
| Lpo | Lactoperoxidase |
|
|
|||
| Ncf2 | Neutrophil cytosolic factor 2 |
|
|
|||
| Scd1 | Stearoyl-Coenzyme A desaturase 1 |
|
|
|
|
|
| Alb | Albumin |
|
|
|||
| Epx | Eosinophil peroxidase |
|
||||
| Gclc | Glutamate-cysteine ligase, catalytic SU |
|
||||
| Gclm | Glutamate cysteine ligase, modifier SU |
|
||||
| Hba1 | Hemoglobin alpha, adult chain 2 |
|
||||
| Mpo | Myeloperoxidase |
|
||||
| Ngb | Neuroglobin |
|
|
|||
| Txnip | Thioredoxin interacting protein |
|
|
|||
| DNA repair | Lig4 | Ligase IV, DNA, ATP-dependent |
|
|
|
|
| Smug1 | Single-strand-selective monofunctional uracil-DNA glycosylase 1 |
|
||||
| Xrcc2 | X-ray repair complementing defective repair in Chinese hamster cells 2 |
|
||||
| Brca1 | Breast cancer 1 |
|
||||
| Total | 27 | 13 | 17 | 6 | 6 | |
upregulation;
downregulation;
up- or downregulation (differences between dose groups); cut-off: FR ≤ − 2.0 or ≥ 2.0
Fig. 11.
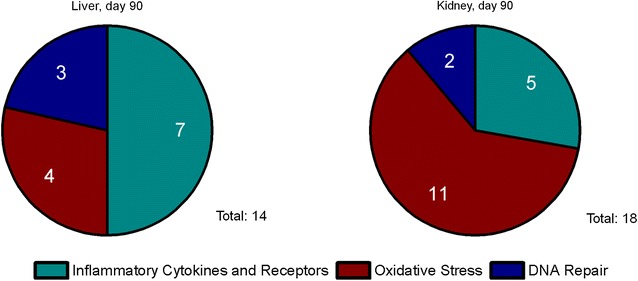
Number and distribution of regulated genes in liver and kidney. The number of regulated genes in liver and kidney tissue after 90-day nanoparticle exposure is illustrated per pathway (pool of all treatment groups)
Nanoparticle retention in liver and kidney tissue
Since some changes in gene expression were detected in liver and kidney tissue after nanoparticle exposure, we determined the amount of Ce in both organs. Figure 12 exemplary shows the level of ionic, particulate and total Ce in the liver during and after 90 day exposure to 3.0 mg/m3 CeO2. The overall amount is low, but there is a significant increase in Ce levels with ongoing exposure and values decrease after the last day of exposure. Differences in the level of ionic and particulate Ce are marginal. A switch from more ionic to more particulate Ce with ongoing nanoparticle exposure and back to higher ionic levels during post-exposure was detected. Table 4 contains all Ce levels measured in liver and kidney for the mid and high dose group. 1.0 mg/m3 CeO2 caused slightly lower values and a similar trend as seen in the high dose group, with peak levels of 2 µg/liver after 90 days exposure. The lower Ce dose groups and BaSO4 were not measured at this point.
Fig. 12.
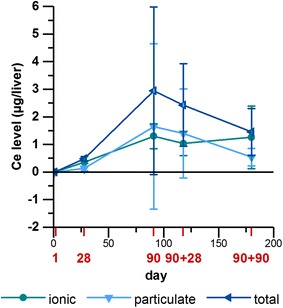
Cerium level in liver tissue after CeO2 nanoparticle exposure. Ionic, particulate and total Ce levels (µg) were determined in liver tissue after one, 28 and 90 days exposure and 28 and 90 days after the end of 90 days exposure to 3.0 mg/m3 CeO2 nanoparticles. Values are expressed as mean ± SD
Table 4.
Amounts of Ce in liver and kidney tissue after CeO2 nanoparticle exposure
| Dose group | Ce | Ce content (µg/organ ± SD) | ||||
|---|---|---|---|---|---|---|
| Day 1 | Days 28 | Days 90 + 1 | Days 90 + 28 | Days 90 + 90 | ||
| 4 (liver) | Ionic | 0.025 ± 0.001 | 0.27 ± 0.068** | 0.717 ± 0.407** | 0.68 ± 0.38** | 0.46 ± 0.70** |
| Particulate | 0.054 ± 0.001 | 0.081 ± 0.04** | 1.308 ± 1.094** | 1.13 ± 1.58** | 0.42 ± 0.24* | |
| Total | 0.079 ± 0.002 | 0.34 ± 0.049** | 2.025 ± 1.182** | 1.81 ± 1.78** | 0.88 ± 0.60* | |
| 5 (liver) | Ionic | < 0.025 | 0.35 ± 0.038** | 1.30 ± 0.45** | 1.03 ± 0.43** | 1.26 ± 1.13** |
| Particulate | < 0.025 | 0.13 ± 0.080 | 1.65 ± 3.00* | 1.40 ± 1.61** | 0.54 ± 0.31** | |
| Total | < 0.025 | 0.48 ± 0.078** | 2.95 ± 3.04** | 2.43 ± 1.50** | 1.45 ± 0.85** | |
| 4 (kidney) | Ionic | 0.11 ± 0.06 | 0.11 ± 0.07 | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.05 ± 0.03 |
| Particulate | 0.04 ± 0.03 | 0.07 ± 0.03 | 0.15 ± 0.10* | 0.07 ± 0.02 | 0.05 ± 0.02 | |
| Total | 0.15 ± 0.07 | 0.18 ± 0.10 | 0.19 ± 0.09 | 0.11 ± 0.02 | 0.09 ± 0.05 | |
| 5 (kidney) | Ionic | 0.018 ± 0.001 | 0.025 ± 0.000 | 0.032 ± 0.012** | 0.041 ± 0.009** | 0.059 ± 0.015** |
| Particulate | 0.025 ± 0.010 | 0.021 ± 0.006 | 0.030 ± 0.003 | 0.033 ± 0.029 | 0.042 ± 0.02 | |
| Total | 0.029 ± 0.011 | 0.033 ± 0.018 | 0.050 ± 0.019 | 0.073 ± 0.033* | 0.084 ± 0.038* | |
Dose groups: 4 = 1.0 mg/m3 CeO2, 5 = 3.0 mg/m3 CeO2; Values are expressed as mean ± SD, * p < 0.05, ** p < 0.01 vs. d 1, n ≤ 5; Kruskal–Wallis–ANOVA with Mann–Whitney U-test as post hoc analysis; detection limit for analysis of liver tissue: 0.025 µg; detection limit for analysis of kidney tissue: 0.015 µg
Discussion
Gene expression analysis is a highly sensitive method, which can yield valuable supportive information on a mechanistic level. Therefore, the gene expression analysis described here was performed as extension of a 90-day inhalation toxicity study on nanomaterial according to OECD TG 413. It aimed on the identification of early biomarkers predicting effects of subacute to chronic nanoparticle exposure. Results of the guideline required parts of the study are discussed in detail elsewhere [1]. Briefly, a marked pulmonary inflammation was detected in response to 3.0 mg/m3 CeO2 nanoparticle inhalation, showing increasing severity with ongoing particle exposure and post-exposure persistency. This particle concentration most likely provoked lung overload, indicating that the inflammation was caused by oversaturated macrophages and related impaired lung clearance. Histopathological findings suggest the risk of long-term effects like fibrosis, resulting from the chronic inflammation reaction. For the mid concentration (1.0 mg/m3 CeO2) also signs of inflammation accompanied by slightly impaired lung clearance were detected. Depending on the calculation method, overload is assumed to be present or absent and therefore making a clear conclusion difficult. At lower CeO2 concentrations (0.1 and 0.3 mg/m3) no signs of inflammation and overload were detected. BaSO4 nanoparticles (50.0 mg/m3) also caused mild inflammatory cell infiltrations in the alveolar space, although major effects were restricted to the nasal cavity, due to particle accumulation. Interestingly, despite the high concentration administered to the animals, BaSO4 was cleared rapidly from the lung without provoking overload.
Local alveolar effects of CeO2 nanoparticles
An overview of CeO2 nanoparticles’ influence on the alveolar compartment in terms of inflammation, oxidative stress and tissue degradation is depicted in Fig. 15.
The described inflammation was reflected by gene expression analysis. In AEII cells a total number of 34 genes were regulated after nanoparticle exposure and thereof 18 were chemokines or interleukins, responsible for immune cell activation and recruitment. This matches inflammatory cell infiltrations in lung tissue and increasing neutrophil and lymphocyte levels in BAL detected in the main study [1]. A major part of the effects exacerbate time- and concentration-dependent, which was reflected by two different phenomena on mRNA level: (1) increase in total number of regulated genes, (2) increase in fold regulation of the individual genes.
Pulmonary immune responses imply the complex interaction between different inflammatory cell types, but also respiratory epithelial cells. In particular AEII cells are suggested to release inflammatory mediators. Most distinct regulation in these cells was detected for the C–C motif chemokines Ccl2, Ccl7, Ccl17 and Ccl22 after 1.0 and 3.0 mg/m3 CeO2 nanoparticle exposure. Ccl2 and Ccl7 are closely related chemoattractant proteins, which bind to the same receptor (Ccr2) and cause similar responses, like macrophage, lymphocyte and neutrophil recruitment [44]. Ccl17 and Ccl22 also react via a common receptor (Ccr4). They are involved in activation of CD4+ T-lymphocytes and monocytes [45, 46]. Our results show upregulation of all these chemokines in response to CeO2 nanoparticle exposure and correlations between the related mediators Ccl2/Ccl7 and Ccl17/Ccl22 (Fig. 13a, b). At exposure conditions resulting in high gene regulation (90 days exposure, CeO2 concentration ≥ 1.0 mg/m3) also significant changes in the amount of target cells for those mediators have been detected in histopathology and/or BAL (particle-laden macrophage accumulations, increased lymphocyte levels) [1]. Until now less was known regarding the expression of the mentioned chemokines in AEII cells in general or in response to nanoparticles. Several studies indicate Ccl2 production in isolated AEII cells after different stimuli [3, 47–52]. Cxcl2 or Ccl2 mRNA expression in either isolated AEII cells or alveolar epithelial cell lines has been described already in response to quartz exposure [6, 53, 54] and Chen et al. [8] demonstrated the expression and release of different Cxcl-motif chemokines by AEII cells after intra-tracheal instillation of carbon nanoparticles and assumed AEII cells to be the major key player in this neutrophil-driven response.
Fig. 13.
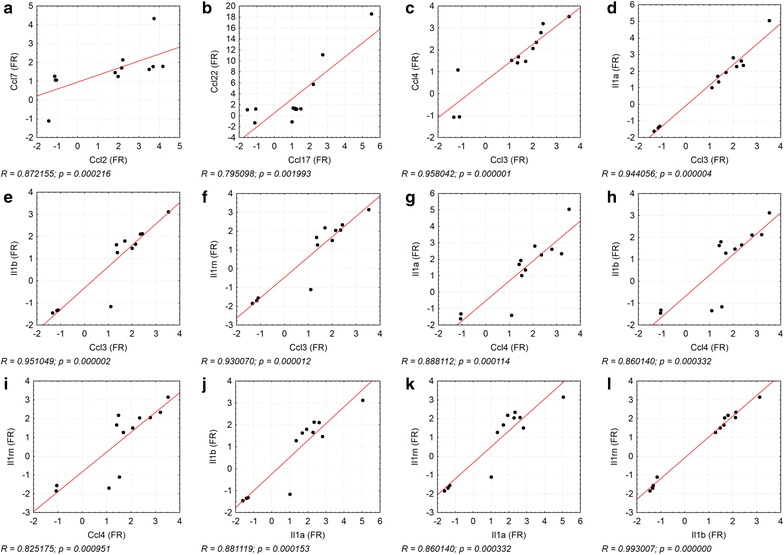
Correlation between fold regulations of chemokines. The correlation between gene expression fold regulation (FR) values of a Ccl2/Ccl7, b Ccl17/Ccl22, c Ccl3/Ccl4, d Ccl3/Il1a, e Ccl3/Il1b, f Ccl3/Il1rn, g Ccl4/Il1a, h Ccl4/Il1b, i Ccl4/Il1rn, j Il1a/Il1b, k Il1a/Il1rn, and l Il1b/Il1rn is illustrated. The individual data points display the FR values of each dose groups 0.1, 0.3, 1.0, and 3.0 mg/m3 CeO2 per time point (day 1, 28, 90). R = Spearman’s correlation coefficient, level of significance p ≤ 0.05; Spearman’s rank correlation analysis
Lower, non-overload concentration levels should help to evaluate potential substance related effects. Our previous investigations indicated that at a concentration of 1.0 and even 3.0 mg/m3 CeO2 effects are likely not exclusively overload-related but also dependent on material-specific characteristics, because they did not induce a volumetric or specific surface-related lung overload [1]. The upregulation of chemokines is therefore to some extent substance related either. CeO2 nanoparticle exposure at 0.1 and 0.3 mg/m3 revealed gene regulation of inflammatory mediators, not markedly affected at higher dose levels (Ccl3, Ccl4, Il-1α, Il-1β, Il-1rn). Ccl3 and Ccl4 are two related mediators with the shared receptor Ccr5. They function as chemoattractant for mainly monocytes and lymphocytes and are expressed by a variety of cells, including inflammatory and epithelial cells [55]. Il-1α and Il-1ß both bind to the interleukin receptor and trigger chemokine release from target cells and immune cell activation. The antagonist Il-1rn competes with Il-1α and Il-1ß for interleukin receptor binding and by this balances induced immune reactions. Evidence is given that Il-1α and Il-1ß stimulate Ccl2 expression in AEII cells [3, 4, 51, 56]. Brabcová et al. [57] demonstrated in addition to Ccl2 effects on Ccl3, Ccl4, and Ccl17 levels in A549 cell cultures in response to Il-1ß and graded this interleukin as a quite potent chemokine stimulant. Il-1ß synthesis has also been found in stem cell-derived lung epithelial cells [58] and isolated mouse AEII cells exposed to TiO2 nanoparticles [59]. So far, data on the influence of nano-CeO2 on the release of the discussed mediators by AEII cells has not been reported by others. Only some measurements of Il-1ß have been performed in BAL [19, 26] or lung tissue [30] of rodents treated with CeO2. However, since no cell type specific analysis was performed the cellular source of Il-1ß remains unknown.
90 days exposure to concentrations of up to 0.3 mg/m3 revealed significant upregulation of the respective mediators followed by a decrease to baseline levels in higher dose groups and highly correlated expression patterns between all mediators (Fig. 13c–l). Since the chemokines and interleukins indicate inflammation which was for this concentration not yet measureable in BAL analysis but clearly present at higher dose levels (BAL and histopathology) [1], the respective mediators might function as promising sensitive biomarkers for nanoparticle exposure on a gene expression level. Also, this gene regulation might indicate an early risk of induced pulmonary inflammation during long-term low dose exposure to CeO2 nanoparticles. In parallel to our 90-day study respective investigations will be provided by a long-term setup with the same CeO2 nanomaterial and concentrations as part of the European program on the regulatory testing of manufactured nanomaterials (NANoREG), in which potential toxic and carcinogenic effects during and after exposure periods of up to 1 year were investigated [BASF, Ludwigshafen, Germany, “combined chronic inhalation toxicity and carcinogenicity study with CeO2 and BaSO4”, (81|0661/10|170)]. The combination of both studies constitutes the ideal base to prove and classify the meanings of the potential early signs for inflammatory effects discussed here.
It could be assumed that AEII cells support the defense against nanoparticles in second instance since upregulation of inflammatory mediators started at later stages of exposure and was dominated by chemokines stimulating monocytes and cells of the adaptive immune system (e.g. lymphocytes). Constant monocyte recruitment supports elimination of the rising particle amount and handling of overload conditions, lymphocytes are usually recruited for delayed defenses. Our histopathology and BAL analysis match this theory as we found neutrophilic inflammation at early stages of exposure, followed by increasing levels of lymphocytes [1]. Respective observations were accompanied by sustained presence of particle-laden macrophages.
Although inflammatory effects of CeO2 nanoparticle exposure were dominating signs of induced oxidative stress have additionally been detected. A marked time-dependent upregulation of Lpo mRNA and protein was measured in AEII cells. Protein analysis further identified the increased Lpo levels as being related to areas of particle-laden macrophage accumulations and inflammatory cell infiltrations. Lpo catalyzes the conversion of thiocyanate to antibacterial hypothiocyanite by use of hydrogen peroxide and belongs to the non-immunological airway defense system. The respiratory Lpo defense system highly depends on H2O2 and is influenced by the presence of inflammatory mediators [60]. Based on research with oxidized SWCNT evidence is given that Lpo functions in nanomaterial degradation [61]. Since CeO2 particles bear redox activity, an interaction with the Lpo defense system seems reasonable and supports substance related effects. The particles might be able to process H2O2 by Ce3+/Ce4+ redox cycling. Although respective properties were reviewed from an anti-oxidative perspective, in terms of mimicking superoxide dismutase (SOD) or catalase activities, CeO2 obviously has the potential to affect H2O2 balance. The SOD like reaction includes the generation of H2O2 from superoxide anion. This would favor the increase of extracellular H2O2 and might cause elevated Lpo levels. Also other theories of oxidative stress generation, including lipid peroxidation [19] by the particles are conceivable. The catalytic activity thus claims consideration of both, pro- and anti-oxidative capacities.
Furthermore, an inflammation-related hypothesis for elevated Lpo levels could be based on respiratory burst. Inflammatory cells eliminate internalized material by the production of ROS. Neutrophils for instance release O2− and H2O2 in response to various stimuli [62]. Lung epithelial cells are further known to physiologically produce H2O2 [63]. Bronchial epithelial cells have been shown to increase H2O2 release in response to the inflammatory mediator IFNγ, leading to stimulation of the Lpo defense system [60]. Such conditions might thus be responsible for imbalance of the system during inflammation (1.0 or 3.0 mg/m3 CeO2), manifested by increased Lpo levels in AEII cells at hotspots of inflammation and the correlation between the amount of neutrophils in BAL and Lpo gene expression levels (Fig. 14a).
Fig. 14.
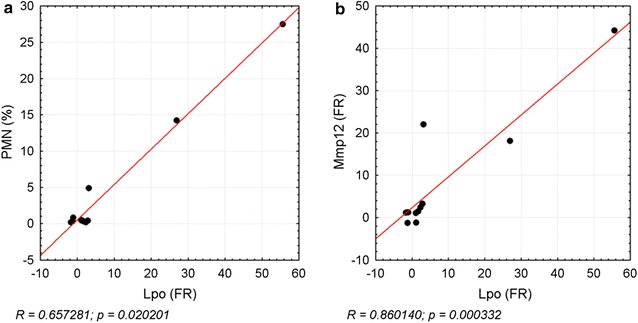
Correlation of Lpo to neutrophils and Mmp12 expression. The correlation of Lpo gene expression fold regulation (FR) values to the percentage of PMN levels in a BAL, and the FR of b Mmp12 is illustrated. Data points of the dose groups 0.1, 0.3, 1.0, and 3.0 mg/m3 CeO2 for each time point (day 1, 28, 90) are depicted. R = Spearman’s correlation coefficient, level of significance p ≤ 0.05; Spearman’s rank correlation analysis
Another upregulated factor participating in ROS generation is Noxo1, which is part of the transmembrane Nox enzyme complex and contributes to extracellular ROS production by intracellular consumption of NADPH [64]. It was significantly upregulated in response to 3.0 mg/m3 CeO2 when also a marked Lpo expression was observed. Van Klaveren et al. [65] demonstrated activation of a membrane bound NAPDH oxidase-like system in isolated rat AEII cells and enzymatic generation of O2− and H2O2 in response to different stimuli. The potential contribution of this system to H2O2 levels suggests a functional relationship between Noxo1 and Lpo levels after CeO2 nanoparticle exposure.
No clear evidence was given for effects on genotoxicity and apoptosis in AEII cells after CeO2 nanoparticle exposure since no distinct gene regulation was measured for the tested DNA repair- or apoptosis-related genes.
However, we further found some indications for chronic particle related effects, including fibrosis and lung cancer. Mmp12 plays an important role in lung tissue remodeling and serves as marker for related acute and chronic lung diseases like fibrosis and COPD [66, 67]. Activated alveolar macrophages produce Mmp12 for neutrophil activation and release of neutrophil elastase. This leads to tissue degradation and finally emphysema development. The persistent neutrophil infiltrations measured during our previous investigations [1], accompanied by increased Mmp12 mRNA in lung epithelia indicate such mechanisms and the risk of developing chronic pathological changes during long-term exposure. Although immunohistochemical staining of lung tissue for Mmp12 did not reveal significantly elevated protein levels, Mmp12 might serve as a valid marker for substance related mechanisms of action as mRNA upregulation occurs at absent lung overload with CeO2 nanoparticles. Overexpression of Mmp12 has been described in mouse lungs after application of asbestos or TiO2 nanoparticles [68, 69]. The latter caused parallel upregulation of Ccl2, 3, 4 and 7, which were also affected in our studies. Ma et al. [16] demonstrated the influence of CeO2 nanoparticles on the development of fibrosis by induction of epithelial–mesenchymal transition of AEII cells after intra-tracheal instillation. Interestingly, disturbance of the Lpo defense system was demonstrated in relation to the pathology of cystic fibrosis [70]. The high correlation of both mediators (Fig. 14b) therefore might function as marker for fibrosis induction in AEII cells triggered through the early inflammation events. This hypothesis is further supported by very slight interstitial fibrosis measured in the main study (90 days post-exposure, 3.0 mg/m3 CeO2) [1].
Figure 15 summarizes the potential relationship between the analyzed mediators and different cell types in response to nanoparticle exposure. Increased gene expression in AEII cells is assumed to affect inflammatory cells and the lung’s oxidative balance. This might further lead to tissue degradation and long-term effects like fibrosis. Lpo, Mmp12 and the inflammatory mediators Ccl2, 7, 17, 22, 3, 4, Il-1α, Il-1β and Il-1rn are promising marker genes for the mentioned effects.
Local alveolar effects of BaSO4 nanoparticles
Considering BaSO4 classification as chemically inert and non-toxic, a generally low and non-adverse response was expected. Therefore BaSO4 NM-220 was selected as reference material without any specific toxicity in the NANoREG long-term study performed by BASF (81|0661/10|170). We measured effects less severe compared to CeO2, despite the at least 16-fold higher concentration, which supports the low effect hypothesis. However, some unexpected findings in BaSO4 exposed lungs have been detected and need to be evaluated.
Pulmonary inflammation and some changes in AEII cell gene expression levels were measured. Major histopathological findings in the nasal cavity with minor inflammation of the alveolar compartment were attributed to particle deposition favoring the upper respiratory tract rather than the alveoli because of high particle accumulation [1]. This might be the reason for the lower impact on gene regulation in AEII cells compared to CeO2.
Interestingly, the majority of genes affected by BaSO4 were also affected by CeO2. Similar to CeO2 the highest response in the group of inflammatory mediators was measured for the C–C-motif chemokine Ccl22. In vitro data on the influence of BaSO4 on macrophages or neutrophils regarding expression of inflammatory mediators and cell migration did not show activation of respective mechanisms [71, 72]. Particles like silica and TiO2 in contrast, had stimulating effects [72]. All in all, the impact of BaSO4 on inflammatory mediator release compared to CeO2 is rather low. Post-exposure decrease of Ccl2 protein levels in BAL supported by decreases in severity of histopathological findings after the end of 90 days exposure [1] further suggest reversibility of the inflammatory reaction.
Although BaSO4 NM-220 was demonstrated to bear no intrinsic oxidative potential [73], the oxidative stress marker Lpo was highly upregulated. Like for CeO2 a marked increase of Lpo protein in AEII cells was identified in areas of inflammation. This verifies a mechanistic relationship between the particle-induced inflammation and oxidative stress. Further, the quantification of the Lpo signal displayed a lower, but significant elevation in comparison to CeO2.
Signs for genotoxicity and apoptosis were minimal, but a strong effect of BaSO4 nanoparticle exposure on Mmp12 was detected. Considering the relation of Mmp12 to chronic pulmonary effects like fibrosis, potential adverse reactions due to long-term BaSO4 become more likely. However, such findings were not detected in previous histopathological investigations [1]. Also, Mmp12 protein levels in lung tissue remain unaffected. Since Mmp12 further indicates inflammation, the elevated gene expression levels in AEII cells detected for BaSO4 might be related to the inflammatory reaction without notifying chronic adverse effects.
Although BaSO4 was cleared from the respiratory tract quite rapid and no overload situation was provoked during 90-day particle exposure [1], the very high amount of particles entering the respiratory tract might be a reason for detection of unexpected effects like increased Lpo, Ccl22 or Mmp12. The phenomenon of fast clearance was pointed out in our previous research [1]. The dissolution of BaSO4 nanoparticles is assumed to be generally low. However, according to Konduru et al. [74], who stated a dissolution rate of < 1% in different media, the fast clearance could be attributed to changes in the dissolution rate because of e.g. switches in surface charge of the nanoparticles. Nevertheless an explicit reason for the phenomenon of fast BaSO4 clearance remains unknown and needs to be further investigated. Our results further show that CeO2 and BaSO4 caused similar responses in AEII cells while the response pattern still allowed differentiation between both substances. The higher inflammatory potential of CeO2 was appropriately reflected. Mediators displaying quite high responses (Lpo, Ccl22, Mmp12) might be suitable markers for nanomaterial toxicity testing. To further clarify the specific function of Lpo or Mmp12 as response to CeO2 and BaSO4 nanoparticles, investigations of the respective markers after exposure to other nanoparticles with a known oxidative, genotoxic or tumorigenic potential are needed.
Systemic effects of CeO2 and BaSO4 nanoparticles on liver and kidney tissue
Nanomaterial generally bears a certain potential to translocate to extra-pulmonary organs after inhalation and organs responsible for excretion are common targets. We investigated gene expression in liver and kidney tissue after 90 days exposure. The number of regulated genes was lower compared to AEII cells with less overlap between pulmonary and extra-pulmonary compartments. Distribution of genes over the different pathways however was similar especially in liver tissue where effects on inflammatory mediators were dominating. Interestingly in the kidney a great portion of genes associated with oxidative stress were downregulated. CeO2 had a higher impact than BaSO4 on both organs.
It has been demonstrated by Aalapati et al. [19] that a small fraction of CeO2 nanoparticles accumulate in liver and kidney and cause histopathological changes after inhalation in mice. The kidney was assumed to be the major extra-pulmonary target organ for nanoparticles. Nemmar et al. [30] comprehensively investigated inflammation, oxidative stress and DNA damage in extrapulmonary tissues after CeO2 intratracheal instillation and reported adverse findings for all endpoints in liver and kidney.
Our findings show upregulation of different C–X–C motif chemokines and receptors responsible for neutrophil recruitment in liver tissue in response to CeO2, indicating inflammatory reactions. In kidney tissue almost all genes affected by CeO2 displayed downregulation, therefore suggesting a rather anti-inflammatory and anti-oxidative effect in this organ. Since potential effects of CeO2 are controversial and positive health effects have been described [75], a positive impact of translocating particles to the kidneys could be conceivable.
As basis for discussion of the present findings, we exemplary determined the level of Ce in liver and kidney tissue during and after nanoparticle exposure to the higher concentrations. We found a small amount of ionic and particulate Ce in the liver, showing an expected development of increase during and decrease after exposure. In the previous study we measured peak levels of 1300 µg Ce retained per lung [1]. This amount derived from a concentration of 3.0 mg/m3 CeO2 NM-212 administered over 90 days (6 h/day, 5 days/week) at a deposition fraction of 9.6%. In the liver we found 3 µg/organ at these conditions. The fraction translocated to the liver from the amount present in the lung was thus 0.23%. However, the increase of Ce levels is significant over time and a contribution to the changes in gene expression is conceivable. It also needs to be considered that modulated gene expression can be caused by other systemically distributed mediators as a response to the present lung inflammation. Also the Ce level was slightly lower after 1.0 mg/m3 CeO2 exposure compared to 3.0 mg/m3 CeO2. This indicates a concentration-dependency which has clearly been detected in the lung as well [1]. Even lower levels are thus assumed for the lower dose groups. At earlier stages of exposure the amount of ionic Ce was higher than the particulate proportion. This indicates that translocation of particles occurs with delay due to barriers and defense mechanisms. As described earlier CeO2 nanoparticles likely influence the cellular balance of ROS due to redox activity. This ability is contributed to ion formation on the nanoparticle surface [75]. The amount of ionic Ce could be a result of this ion formation. The high Lpo levels detected in this study indicate oxidative stress in the lung. The amount of ionic Ce in this tissue however, was negligible [1]. The ion content could therefore not be correlated to the level of oxidative stress. In liver and kidney tissue the overall modulation of gene regulation was too low to adequately assess a potential correlation between ion formation and oxidative stress in these organs. The effects on gene expression and the amount of Ce in extra-pulmonary organs was overall very low. The question in how far these findings contribute to potential adverse effects thus remains unanswered. No significant increases of Ce levels were detected in kidney tissue. It could be assumed that the downregulation of several genes was rather caused by other signals, like endogenous mediators, than the nanoparticles. Due to the low response to BaSO4 in terms of gene expression after 90 days and the less clear results for CeO2 we did not include barium retention in our investigations at this point.
Conclusion
Our results suggest that CeO2 related lung inflammation was supported by chemokine release of AEII cells and characterized by increased monocyte activation with subsequent neutrophil and lymphocyte infiltrations. Stimulation of rat AEII cells is likely not exclusively overload but also substance related. At low concentrations it is dominated by interleukins like Il-1α, Il-1ß and chemokine expression. With increasing concentrations and induction of lung overload gene regulation exacerbates and the chemokine pattern switches. AEII cells further respond with signs of oxidative stress indicated by increased Lpo and Noxo1 in the presences of CeO2 and the mediators can be considered as potential early marker for nanoparticle oxidative stress induction after inhalation. Genes related to genotoxicity and apoptosis were not markedly affected by CeO2 exposure. However, due to the upregulation of Mmp12 a relation to potential long-term effects like fibrosis and lung cancer could be assumed. Respective findings need to be related to the long-term inhalation toxicity study (BASF, Ludwigshafen, Germany) to further support significance.
BaSO4 nanoparticle exposure has a rather low influence on the expression of inflammatory mediators in AEII cells. However, we detected some changes in gene regulation, which supports the assumption that the nanomaterial affects the organism after inhalation although it is considered as chemically inert. Interestingly, mediators like Lpo, Ccl22 and Mmp12, highly affected by CeO2 exposure also responded to BaSO4. Such mediators are promising biomarkers for nanomaterial toxicity testing.
The overall effect on gene expression in liver and kidney was low. However, especially in the liver some inflammatory mediators were upregulated. Also, low but significant Ce levels were present in the liver. A contribution of the particles to changes in gene expression of extra-pulmonary organs therefore still needs to be taken into consideration and further research should focus on this issue.
Methods
Test substances
The test materials cerium dioxide NM-212 and barium sulfate NM-220 belong to the Nanomaterial Repository of the European Commission Joint Research Center (JRC, Ispra, Italy) and were provided by the Fraunhofer Institute for Molecular Biology and Applied Ecology (Schmallenberg, Germany).
Animals
For this study female Wistar rats [Crl:WI (Han)] at the age of 10 weeks during start of exposure were used (Charles River, Sulzfeld, Germany). Beforehand, animals were acclimatized to laboratory conditions for 1 week, trained for restraint in nose-only tubes over 3 weeks and were randomly distributed to control and treatment groups, respectively. The following conditions were set for animal housing: 20–24 °C, 40–70% relative humidity, 12 h light/dark cycle. Laboratory diet (“V1534”, sniff Spezialdiaeten GmbH, Soest, Germany) and water was supplied ad libitum. All experiments and animal handling was conducted in compliance with the Regulations of the German Animal Protection Law (Tierschutzgesetz of May 18, 2006).
Nose-only inhalation
The examinations described here were performed as additional part to the conventional investigation of a 90-day inhalation toxicity study according to OECD TG 413. For gene expression analysis, 90 animals were exposed to clean air or the test substances via nose-only inhalation for one, 28 and 90 days in concentrations of 0.1, 0.3, 1.0 and 3.0 mg/m3 for CeO2 nanoparticles and 50.0 mg/m3 for BaSO4 nanoparticles (5 rats/dose group/time point). Aerosol generation was done via dry powder dispersion using a dispersion nozzle developed at Fraunhofer ITEM and the respective aerosol generation system [76]. Aerosol concentrations were constantly checked via an aerosol photometer (Fraunhofer ITEM, Hannover, Germany). The MMAD of the samples was determined by gravimetry (Marple 298 Personal Cascade Impactor, Thermo Fisher Scientific, Dreieich, Germany). Animals were kept in the inhalation system 5 days a week for 6 h each day. They were checked for clinical signs daily before and after exposure. Bodyweight as well as food and water consumption was recorded weekly.
Gene expression analysis
Alveolar epithelial type II cell isolation and organ extraction
After one, 28 and 90 days of exposure to either clean air, the different CeO2 NM-212 concentrations or BaSO4 NM-220, rat lungs were prepared on the following day for isolation of AEII cells and subsequent gene expression analysis. The method of AEII isolation is based on the work of Richards et al. [77] and Dobbs et al. [78] and well established in our institute [79]. We further performed comprehensive methodical research on this topic in mice [80]. Minor modifications on the current protocol have been taken for the work with rats. Briefly, animals were anesthetized (Ketamine/Xylazine, 10:1; bela-pharm, Vechta, Germany/Bayer, Leverkusen, Germany) via intraperitoneal injection. For lung perfusion, the trachea was uncovered and cut to introduce a cannula for artificial ventilation. Abdominal cavity and thorax were opened and a cannula was inserted via the vena cava inferior into the right ventricle. Pulmonary perfusion was done with PBS (5 mL/min, 25 mL Biochrom, Berlin, Germany) and a peristaltic pump while the lung was constantly manually ventilated. An incision in the left atrium served as perfusate outflow. Finally, the lung was explanted for subsequent tissue digestion and cell isolation. During sacrifice the caudate lobe of the liver as well as the right kidney were removed and stored at − 80 °C for RNA isolation. The isolated lung was flushed with PBS, treated with dispase (Sigma-Aldrich, Taufkirchen, Germany) and manually dissected. Respective steps were followed by DNase digestion (Sigma-Aldrich, Taufkirchen, Germany) and passage through two nylon sieves with different mesh size (250, 60 µM). After several steps of centrifugation IgG panning (Sigma-Aldrich, Taufkirchen, Germany) was performed to remove unwanted cell types, including macrophages. One hour incubation was followed by another DNase digestion. Finally, the isolated cells were separated by centrifugation and the pellet was stored at − 80 °C for RNA isolation. AEII cell extraction from rat lungs yielded 5.3 ± 2.5 × 106 cells per individual which was sufficient for subsequent processing.
RNA Isolation
RNA isolation from AEII cells, liver and kidney tissue was performed using the RNeasy Mini Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions including a DNase treatment procedure (RNase-Free DNase Set, QIAGEN, Hilden, Germany). RNA extraction and subsequent gene expression analysis was done for AEII cells after one, 28 and 90 days of nanoparticle exposure (90 samples in total). For liver and kidney tissues 90-day exposure samples were processed (in total 30 samples each). Amount and quality of RNA were measured with the NanoDrop ND-1000 Spectrophotometer, Version 3.7 (Thermo Fisher Scientific, Dreieich, Germany) and the Agilent 2100 Bioanalyzer (Agilent Technologies, Ratingen, Germany), respectively. The amount of RNA extracted from liver tissue was highest (78.4 ± 21.3 µg), followed by kidney (45.2 ± 6.5 µg) and AEII cells (15.6 ± 5.6 µg). Measurements of RNA quality yielded mean RNA integrity numbers (RIN) > 7.0 for liver, kidney and AEII cells, respectively, indicating RNA samples of good quality.
RT2 profiler PCR arrays
To screen a broad spectrum of potential effects of nanoparticle exposure, the following RT2 profiler PCR arrays (SABiosciences/QIAGEN, Hilden, Germany) were applied for AEII samples: inflammatory cytokines and receptors (PARN-011Z), oxidative stress (PARN-065Z), DNA repair (PARN-042Z), apoptosis (PARN-012Z), lung cancer (PARN-134Z). For RNA isolated from liver and kidney the same arrays were used except of the lung cancer array. The commercially available arrays contain qPCR assays for the analysis of 84 different genes related to the respective end points in a 96 well format (84 genes, 5 potential reference genes, 7 quality controls). The quality controls consist of a genomic DNA contamination control (1×), a reverse transcription efficiency control (3×) and a PCR efficiency control (3×). For the present study we created customized arrays, by pooling four commercial arrays in a 384 well format. The fifth array was spotted four times on a 384 well plate for simultaneous analysis of four samples. Some genes are overlapping between the pathways and therefore in total 391 different genes were analyzed per AEII RNA sample and 325 genes per RNA sample extracted from liver and kidney tissues (Additional file 2: Table S2). In a first step, cDNA synthesis was performed using the RT2 First Strand Kit (SABiosciences/QIAGEN, Hilden, Germany), following the manufacturer’s instructions. Subsequently, RT2 Profiler PCR arrays were conducted according to the manufacturer’s instructions using the real-time PCR system ViiA™7 (Applied Biosystems/Thermo Fisher Scientific Inc., Dreieich, Germany). An amount of 1.19 ng was applied per single RTqPCR assay.
Data evaluation
Data analysis of exported “Cycle Threshold” (CT) values was performed based on the comparative ∆∆CT method described by Schmittgen and Livak [81], using the provided SABiosciences PCR Array Data Analysis Template Excel® and web-based software (SABiosciences/QIAGEN, Hilden, Germany; https://www.qiagen.com/de/products/genes%20and%20pathways/data-analysis-center-overview-page/?UID=70f8b4af-789d-4788-af6f-b12ebd800888#). One to three stable expressed reference genes were chosen for each time point and tissue for normalization using NormFinder [82] and geNorm algorithm [83] as part of the GenEx Professional 6 Software (bioMCC, Freising, Germany). The fold regulation (FR) values generated for each gene describe the factor of up- or downregulation in a certain treatment group related versus the control group. Genes displaying FR values ≤ − 2 or ≥ 2 were considered as relevant. Genes with high CT values (≥ 35) or abnormal melting curves were excluded. Statistical significance of FR values was checked with Student’s T test (p ≤ 0.05). CT values, if normal distributed, were further checked for outliers within the group of five animals for each experimental condition and gene, according to Grubbs [84]. Respective values were eliminated if necessary, FR values were recalculated and statistical significance testing was repeated as described above. Genes with FR values ≤ − 2 or ≥ 2 were selected for further analysis concerning function and relevance in the context of the study focus, independent of their statistical significance. Already changes in individual animals are important, because a completely identical genetic background of the animals should not reasonably be assumed.
Correlation analysis was performed using fold regulation (FR) values (gene expression data) or mean percentages (BAL data, main study) of all CeO2 dose groups and time points for the two compared variables. Data were correlated using Spearman rank correlation analysis (p ≤ 0.05).
Cytokine levels in bronchoalveolar lavage
Levels of the cytokines Ccl2, Ccl20, Il-1α, and Il-1β were measured in bronchoalveolar lavages (BAL) of rat lungs after one and 28 days as well as one, 28 and 90 days post-90-day exposure to clean air, the different CeO2 NM-212 concentrations or BaSO4 NM-220. BAL was performed according to Henderson et al. [85] with minor modifications. Lungs were lavaged twice using 4 mL 0.9% NaCl. For cytokine analysis 1 mL of the BAL fluid was removed. Cytokine levels were determined using a MULTI-SPOT® 4 Spot Cytokine Custom Rat 4-Plex kit for the four analytes (Meso Scale Discovery, Rockville, USA), according to the manufacturer’s instructions.
Immunohistochemistry
For immunohistological analysis, two consecutive lung tissue sections were prepared. Examinations were done in rats exposed to either clean air, 3.0 mg/m3 CeO2 NM-212 or 50.0 mg/m3 BaSO4 NM-220 for one, 28 or 90 days. Lungs were fixed in 10% formalin and trimmed based on the work of Kittel et al. [86]. To detect the presence of lactoperoxidase (Lpo, rabbit polyclonal, Acris, NBP1-87010) and matrixmetalloproteinase 12 (Mmp12, rabbit polyclonal, Abcam, ab66157) antibodies directed against those markers were applied as described by Rittinghausen et al. [87]. Immunohistochemically stained lung sections were counterstained with hematoxylin and digitalized using slide scanner (MiraxScanner, Zeiss, Germany) and quantified using image analyzing software (ZeissZen, Zeiss, Germany). Quantified Lpo and Mmp12 levels were statistically evaluated using Kruskal–Wallis–ANOVA with Mann–Whitney U-test as post hoc analysis.
Retention analytics
Cerium contents were measured in liver and kidney tissue of five animals of the mid and high dose group after one and 28 exposure days and one, 28 and 90 days post-90-day exposure. The isotopes 140Ce/142Ce were quantified via inductively coupled plasma mass spectrometry (ICP-MS) using a quadrupole ICP-MS system (X-Serie II, Thermo Fisher Scientific). The shredded tissue was lyophilized for at least 6 h (0.37 mbar). Prior and subsequently to freeze-drying organ weights were recorded. Plasma ashing (cool plasma conditions, 400 W, 1 mbar O2, 24 h) and subsequent microwave digestion (H2SO4, 96%, supra quality, max. 500 W) was performed to further remove organic material. To distinguish between CeO2 particles and soluble CeO2 a semiquantitative technique was implemented using nuclear pore filters (diameter: 0.1 µm).
Additional files
Additional file 1: Table S1. Fold regulation of regulated genes. List of fold regulation and standard deviation values of the regulated genes listed in Tables 2 and 3 of the manuscript. Values are shown for every dose group and time point.
Additional file 2: Table S2. Analyzed genes per array. List of all analyzed genes (short and long name) separated per PCR profiler array.
Authors’ contributions
DS was responsible for study planning and management. She was further involved in laboratory work including animal exposure, AEII cell isolation and gene expression analysis. She processed all gene expression data, performed statistical evaluations and wrote major parts of the manuscript. MN reviewed gene expression analyses and highly contributed to manuscript writing with valuable scientific and structural input. DS performed immunohistochemical tissue staining and analysis, provided the photographs and was involved in interpretation and discussion of immunohistochemical data. HK performed chemical analytics of the substances and was involved in retention data analysis. TH supervised AEII cell isolation and contributed to manuscript reviewing. CD initiated and supervised the study and importantly contributed to manuscript reviewing. OC was involved in project initiation and management as well as interpretation and discussion of results. All authors read and approved the final manuscript.
Acknowledgements
The authors thank all other employees of Fraunhofer ITEM, involved in the project for excellent technical assistance and intellectual input. Special thanks to Janosch Kretzschmann for creative support and critical review of the overview figure.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All experiments were conducted and approved according to the German Animal Welfare Act by the local authority at the LAVES Niedersachsen, Hannover, Germany, No. 33.12-42502-04-14/1564.
Funding
All experiments were funded by the German Federal Ministry of Education and Research (BMBF) within the project InhalT-90: “90 days inhalation testing with CeO2 in the rat and subsequent analysis of gene expression profiles for the early detection of toxic/carcinogenic effects” (tag: 03X0149).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- 8-OHdG
8-oxo-2′-deoxyguanosine
- AEI cells
alveolar epithelial cells type I
- AEII cells
alveolar epithelial cells type II
- Alb
albumin
- BAL
bronchoalveolar lavage
- Birc5
baculoviral IAP repeat-containing 5
- Brca1
breast cancer 1
- Ccl11
C–C motif chemokine ligand 11
- Ccl17
C–C motif chemokine ligand 17
- Ccl2
C–C motif chemokine ligand 2
- Ccl20
C–C motif chemokine ligand 20
- Ccl22
C–C motif chemokine ligand 22
- Ccl24
C–C motif chemokine ligand 24
- Ccl3
C–C motif chemokine ligand 3
- Ccl4
C–C motif chemokine ligand 4
- Ccl7
C–C motif chemokine ligand 7
- Ccl9
chemokine (C–C motif) ligand 9
- Ccr2
chemokine (C–C motif) receptor 2
- Cd40lg
CD40 ligand
- COPD
chronic obstructive lung disease
- Cx3cl1
chemokine (C–X3–C motif) ligand 1
- Cx3cr1
C–X3–C motif chemokine receptor 1
- Cxcl12
chemokine (C–X–C motif) ligand 12
- Cxcl9
C–X–C motif chemokine ligand 9
- Cxcr2
chemokine (C–X–C motif) receptor 2
- Cxcr3
chemokine (C–X–C motif) receptor 3
- Cxcr5
chemokine (C–X–C motif) receptor 5
- Cyp1b1
cytochrome P450, family 1, subfamily b, polypeptide 1
- Epx
eosinophil peroxidase
- Exo1
exonuclease 1
- Fabp4
fatty acid binding protein 4
- FR
fold regulation
- Gclc
glutamate-cysteine ligase, catalytic SU
- Gclm
glutamate cysteine ligase, modifier SU
- Gpx2
glutathione peroxidase 2
- Hba1
hemoglobin alpha, adult chain 2
- IFNγ
interferon gamma
- Il1rn
interleukin 1 receptor antagonist
- Il1α
interleukin 1 alpha
- Il1β
interleukin 1 beta
- Il2rb
interleukin 2 receptor, beta
- Il2rg
interleukin 2 receptor, gamma
- Krt1
keratin 1
- LDH
lactate dehydrogenase
- Lig4
ligase IV, DNA, ATP-dependent
- Lpo
lactoperoxidase
- LYMPH
lymphocyte
- Mmp12
matrix metallopeptidase 12
- MPH
macrophage
- Mpo
myeloperoxidase
- Msh5
mutS homolog 5
- Mutyh
MutY homolog (E. coli)
- NADPH
nicotineamide adenine dinucleotide phosphate
- Ncf2
neutrophil cytosolic factor 2
- Ngb
neuroglobin
- Noxo1
NADPH oxidase organizer 1
- OECD
Organization for Economic Co-operation and Development
- Opcml
opioid binding protein/cell adhesion molecule-like
- Osm
oncostatin M
- Pf4
Platelet factor 4
- PMN
polymorphonuclear neutrophil
- ROS
reactive oxygen species
- Scd1
stearoyl-Coenzyme A desaturase 1
- Smug1
single-strand-selective monofunctional uracil-DNA glycosylase 1
- SOD
superoxide dismutase
- Spp1
secreted phosphoprotein 1
- SWCNT
single walled carbon nanotubes
- Tcf21
transcription factor 21
- Thbs2
thrombospondin 2
- Tnfsf4
tumor necrosis factor superfamily member 4
- Top2a
topoisomerase (DNA) II alpha
- Txnip
thioredoxin interacting protein
- Xrcc2
X-ray repair complementing defective repair in Chinese hamster cells 2
- γ-H2AX
histon γ-H2AX
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s12951-018-0343-4) contains supplementary material, which is available to authorized users.
Contributor Information
Daniela Schwotzer, Email: daniela.schwotzer@item.fraunhofer.de.
Monika Niehof, Email: monika.niehof@item.fraunhofer.de.
Dirk Schaudien, Email: dirk.schaudien@item.fraunhofer.de.
Heiko Kock, Email: heiko.kock@item.fraunhofer.de.
Tanja Hansen, Email: tanja.hansen@item.fraunhofer.de.
Clemens Dasenbrock, Email: clemens.dasenbrock@item.fraunhofer.de.
Otto Creutzenberg, Email: otto.creutzenberg@item.fraunhofer.de.
References
- 1.Schwotzer D, Ernst H, Schaudien D, Kock H, Pohlmann G, Dasenbrock C, Creutzenberg O. Effects from a 90-day inhalation toxicity study with cerium oxide and barium sulfate nanoparticles in rats. Part Fibre Toxicol. 2017;14:23. doi: 10.1186/s12989-017-0204-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Pison U, Max M, Neuendank A, Weißbach S, Pietschmann S. Host defence capacities of pulmonary surfactant: evidence for ‘non-surfactant’ functions of the surfactant system. Eur J Clin Invest. 1994;24:586–599. doi: 10.1111/j.1365-2362.1994.tb01110.x. [DOI] [PubMed] [Google Scholar]
- 3.Paine R, Rolfe MW, Standiford TJ, Burdick MD, Rollins BJ, Strieter RM. MCP-1 expression by rat type II alveolar epithelial cells in primary culture. J Immunol. 1993;150:4561–4570. [PubMed] [Google Scholar]
- 4.Standiford TJ, Kunkel SL, Phan SH, Rollins BJ, Strieter RM. Alveolar macrophage-derived cytokines induce monocyte chemoattractant protein-1 expression from human pulmonary type II-like epithelial cells. J Biol Chem. 1991;266:9912–9918. [PubMed] [Google Scholar]
- 5.van Berlo D, Knaapen AM, van Schooten F-J, Schins RP, Albrecht C. NF-kappaB dependent and independent mechanisms of quartz-induced proinflammatory activation of lung epithelial cells. Part Fibre Toxicol. 2010;7:13. doi: 10.1186/1743-8977-7-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Driscoll KE, Howard BW, Carter JM, Asquith T, Johnston C, Detilleux P, et al. Alpha-quartz-lnduced chemokine expression by rat lung epithelial cells. Am J Pathol. 1996;149:1627–1637. [PMC free article] [PubMed] [Google Scholar]
- 7.Barlow PG, Clouter-Baker A, Donaldson K, Maccallum J, Stone V. Carbon black nanoparticles induce type II epithelial cells to release chemotaxins for alveolar macrophages. Part Fibre Toxicol. 2005;2:11. doi: 10.1186/1743-8977-2-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chen S, Yin R, Mutze K, Yu Y, Takenaka S, Konigshoff M, Stoeger T. No involvement of alveolar macrophages in the initiation of carbon nanoparticle induced acute lung inflammation in mice. Part Fibre Toxicol. 2016;13:33. doi: 10.1186/s12989-016-0144-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kuhn DA, Vanhecke D, Michen B, Blank F, Gehr P, Petri-Fink A, Rothen-Rutishauser B. Different endocytotic uptake mechanisms for nanoparticles in epithelial cells and macrophages. Beilstein J Nanotechnol. 2014;5:1625–1636. doi: 10.3762/bjnano.5.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Thorley AJ, Ruenraroengsak P, Potter TE, Tetley TD. Critical determinants of uptake and translocation of nanoparticles by the human pulmonary alveolar epithelium. ACS Nano. 2014;8:11778–11789. doi: 10.1021/nn505399e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kemp SJ, Thorley AJ, Gorelik J, Seckl MJ, O’Hare MJ, Arcaro A, et al. Immortalization of human alveolar epithelial cells to investigate nanoparticle uptake. Am J Respir Cell Mol Biol. 2008;39:591–597. doi: 10.1165/rcmb.2007-0334OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schumann C, Schubbe S, Cavelius C, Kraegeloh A. A correlative approach at characterizing nanoparticle mobility and interactions after cellular uptake. J Biophotonics. 2012;5:117–127. doi: 10.1002/jbio.201100064. [DOI] [PubMed] [Google Scholar]
- 13.Schaudien D, Knebel J, Creutzenberg O. In vitro study revealed different size behavior of different nanoparticles. J Nanopart Res. 2012;14:593. doi: 10.1007/s11051-012-1039-6. [DOI] [Google Scholar]
- 14.Geiser M, Quaile O, Wenk A, Wigge C, Eigeldinger-Berthou S, Hirn S, et al. Cellular uptake and localization of inhaled gold nanoparticles in lungs of mice with chronic obstructive pulmonary disease. Part Fibre Toxicol. 2013;10:19. doi: 10.1186/1743-8977-10-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Conhaim RL, Dovi WF, Watson KE, Spiegel CA, Harms BA. Bacteremia does not affect cellular uptake of ultrafine particles in the lungs of rats. Anat Rec (Hoboken) 2011;294:550–557. doi: 10.1002/ar.21326. [DOI] [PubMed] [Google Scholar]
- 16.Ma J, Bishoff B, Mercer RR, Barger M, Schwegler-Berry D, Castranova V. Role of epithelial–mesenchymal transition (EMT) and fibroblast function in cerium oxide nanoparticles-induced lung fibrosis. Toxicol Appl Pharmacol. 2017;323:16–25. doi: 10.1016/j.taap.2017.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Oberdörster G. Lung particle overload: implications for occupational exposure to particles. Regul Toxicol Pharmacol. 1995;27:123–135. doi: 10.1006/rtph.1995.1017. [DOI] [PubMed] [Google Scholar]
- 18.Schins RPF. Mechanisms of genotoxicity of particles and fibers. Inhal Toxicol. 2002;14:57–78. doi: 10.1080/089583701753338631. [DOI] [PubMed] [Google Scholar]
- 19.Aalapati S, Ganapathy S, Manapuram S, Anumolu G, Prakya BM. Toxicity and bio-accumulation of inhaled cerium oxide nanoparticles in CD1 mice. Nanotoxicology. 2014;8:786–798. doi: 10.3109/17435390.2013.829877. [DOI] [PubMed] [Google Scholar]
- 20.Demokritou P, Gass S, Pyrgiotakis G, Cohen JM, Goldsmith W, McKinney W, et al. An in vivo and in vitro toxicological characterisation of realistic nanoscale CeO(2) inhalation exposures. Nanotoxicology. 2013;7:1338–1350. doi: 10.3109/17435390.2012.739665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gosens I, Mathijssen LE, Bokkers BGH, Muijser H, Cassee FR. Comparative hazard identification of nano- and micro-sized cerium oxide particles based on 28-day inhalation studies in rats. Nanotoxicology. 2014;8:643–653. doi: 10.3109/17435390.2013.815814. [DOI] [PubMed] [Google Scholar]
- 22.Keller J, Wohlleben W, Ma-Hock L, Strauss V, Groters S, Kuttler K, et al. Time course of lung retention and toxicity of inhaled particles: short-term exposure to nano-ceria. Arch Toxicol. 2014;88:2033–2059. doi: 10.1007/s00204-014-1349-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Landsiedel R, Ma-Hock L, Hofmann T, Wiemann M, Strauss V, Treumann S, et al. Application of short-term inhalation studies to assess the inhalation toxicity of nanomaterials. Part Fibre Toxicol. 2014;11:16. doi: 10.1186/1743-8977-11-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Larsen ST, Jackson P, Poulsen SS, Levin M, Jensen KA, Wallin H, et al. Airway irritation, inflammation, and toxicity in mice following inhalation of metal oxide nanoparticles. Nanotoxicology. 2016;10:1254–1262. doi: 10.1080/17435390.2016.1202350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Morimoto Y, Izumi H, Yoshiura Y, Tomonaga T, Oyabu T, Myojo T, et al. Pulmonary toxicity of well-dispersed cerium oxide nanoparticles following intratracheal instillation and inhalation. J Nanopart Res. 2015;17:442. doi: 10.1007/s11051-015-3249-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Srinivas A, Rao PJ, Selvam G, Murthy PB, Reddy PN. Acute inhalation toxicity of cerium oxide nanoparticles in rats. Toxicol Lett. 2011;205:105–115. doi: 10.1016/j.toxlet.2011.05.1027. [DOI] [PubMed] [Google Scholar]
- 27.Eom H-J, Choi J. Oxidative stress of CeO2 nanoparticles via p38-Nrf-2 signaling pathway in human bronchial epithelial cell, beas-2B. Toxicol Lett. 2009;187:77–83. doi: 10.1016/j.toxlet.2009.01.028. [DOI] [PubMed] [Google Scholar]
- 28.Lin W, Huang Y-W, Zhou X-D, Ma Y. Toxicity of cerium oxide nanoparticles in human lung cancer cells. Int J Toxicol. 2006;25:451–457. doi: 10.1080/10915810600959543. [DOI] [PubMed] [Google Scholar]
- 29.Mittal S, Pandey AK. Cerium oxide nanoparticles induced toxicity in human lung cells: role of ROS mediated DNA damage and apoptosis. Biomed Res Int. 2014;2014:891934. doi: 10.1155/2014/891934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nemmar A, Yuvaraju P, Beegam S, Fahim MA, Ali BH. Cerium oxide nanoparticles in lung acutely induce oxidative stress, inflammation, and DNA damage in various organs of mice. Oxid Med Cell Longev. 2017;2017:9639035. doi: 10.1155/2017/9639035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rubio L, Annangi B, Vila L, Hernandez A, Marcos R. Antioxidant and anti-genotoxic properties of cerium oxide nanoparticles in a pulmonary-like cell system. Arch Toxicol. 2016;90:269–278. doi: 10.1007/s00204-015-1468-y. [DOI] [PubMed] [Google Scholar]
- 32.Xia T, Kovochich M, Liong M, Madler L, Gilbert B, Shi H, et al. Comparison of the mechanism of toxicity of zinc oxide and cerium oxide nanoparticles based on dissolution and oxidative stress properties. ACS Nano. 2008;2:2121–2134. doi: 10.1021/nn800511k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kumari M, Kumari SI, Grover P. Genotoxicity analysis of cerium oxide micro and nanoparticles in Wistar rats after 28 days of repeated oral administration. Mutagenesis. 2014;29:467–479. doi: 10.1093/mutage/geu038. [DOI] [PubMed] [Google Scholar]
- 34.Kumari M, Kumari SI, Kamal SSK, Grover P. Genotoxicity assessment of cerium oxide nanoparticles in female Wistar rats after acute oral exposure. Mutat Res Genet Toxicol Environ Mutagen. 2014;775–776:7–19. doi: 10.1016/j.mrgentox.2014.09.009. [DOI] [PubMed] [Google Scholar]
- 35.El Yamani N, Collins AR, Runden-Pran E, Fjellsbo LM, Shaposhnikov S, Zielonddiny S, Dusinska M. In vitro genotoxicity testing of four reference metal nanomaterials, titanium dioxide, zinc oxide, cerium oxide and silver: towards reliable hazard assessment. Mutagenesis. 2017;32:117–126. doi: 10.1093/mutage/gew060. [DOI] [PubMed] [Google Scholar]
- 36.Cordelli E, Keller J, Eleuteri P, Villani P, Ma-Hock L, Schulz M, et al. No genotoxicity in rat blood cells upon 3- or 6-month inhalation exposure to CeO2 or BaSO4 nanomaterials. Mutagenesis. 2016 doi: 10.1093/mutage/gew005. [DOI] [PubMed] [Google Scholar]
- 37.Ma JY, Mercer RR, Barger M, Schwegler-Berry D, Scabilloni J, Ma JK, Castranova V. Induction of pulmonary fibrosis by cerium oxide nanoparticles. Toxicol Appl Pharmacol. 2012;262:255–264. doi: 10.1016/j.taap.2012.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ma JYC, Young S-H, Mercer RR, Barger M, Schwegler-Berry D, Ma JK, Castranova V. Interactive effects of cerium oxide and diesel exhaust nanoparticles on inducing pulmonary fibrosis. Toxicol Appl Pharmacol. 2014;278:135–147. doi: 10.1016/j.taap.2014.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Geraets L, Oomen AG, Schroeter JD, Coleman VA, Cassee FR. Tissue distribution of inhaled micro- and nano-sized cerium oxide particles in rats: results from a 28-day exposure study. Toxicol Sci. 2012;127:463–473. doi: 10.1093/toxsci/kfs113. [DOI] [PubMed] [Google Scholar]
- 40.He X, Zhang H, Ma Y, Bai W, Zhang Z, Lu K, et al. Lung deposition and extrapulmonary translocation of nano-ceria after intratracheal instillation. Nanotechnology. 2010;21:285103. doi: 10.1088/0957-4484/21/28/285103. [DOI] [PubMed] [Google Scholar]
- 41.Li D, Morishita M, Wagner JG, Fatouraie M, Wooldridge M, Eagle WE, et al. In vivo biodistribution and physiologically based pharmacokinetic modeling of inhaled fresh and aged cerium oxide nanoparticles in rats. Part Fibre Toxicol. 2016;13:45. doi: 10.1186/s12989-016-0156-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Molina RM, Konduru NV, Jimenez RJ, Pyrgiotakis G, Demokritou P, Wohlleben W, Brain JD. Bioavailability, distribution and clearance of tracheally instilled, gavaged or injected cerium dioxide nanoparticles and ionic cerium. Environ Sci Nano. 2014;1:561–573. doi: 10.1039/C4EN00034J. [DOI] [Google Scholar]
- 43.Nalabotu SK, Kolli MB, Triest WE, Ma JY, Manne Nandini D P K, Katta A, et al. Intratracheal instillation of cerium oxide nanoparticles induces hepatic toxicity in male Sprague–Dawley rats. Int J Nanomed. 2011;6:2327–2335. doi: 10.2147/IJN.S25119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Proost P, Wuyts A, van Damme J. Human monocyte chemotactic proteins-2 and -3: structural and functional comparison with MCP-1. J Leukoc Biol. 1996;59:67–74. doi: 10.1002/jlb.59.1.67. [DOI] [PubMed] [Google Scholar]
- 45.Nakanishi T, Imaizumi K, Hasegawa Y, Kawabe T, Hashimoto N, Okamoto M, Shimokata K. Expression of macrophage-derived chemokine (MDC)/CCL22 in human lung cancer. Cancer Immunol Immunother. 2006;55:1320–1329. doi: 10.1007/s00262-006-0133-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Imai T, Nagira M, Takagi S, Kakizaki M, Nishimura M, Wang J, et al. Selective recruitment of CCR4-bearing Th2 cells toward antigen-presenting cells by the CC chemokines thymus and activation-regulated chemokine and macrophage-derived chemokine. Int Immunol. 1999;11:81–88. doi: 10.1093/intimm/11.1.81. [DOI] [PubMed] [Google Scholar]
- 47.Neagos J, Standiford TJ, Newstead MW, Zeng X, Huang SK, Ballinger MN. Epigenetic regulation of tolerance to toll-like receptor ligands in alveolar epithelial cells. Am J Respir Cell Mol Biol. 2015;53:872–881. doi: 10.1165/rcmb.2015-0057OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bello-Irizarry SN, Wang J, Olsen K, Gigliotti F, Wright TW. The alveolar epithelial cell chemokine response to pneumocystis requires adaptor molecule MyD88 and interleukin-1 receptor but not toll-like receptor 2 or 4. Infect Immun. 2012;80:3912–3920. doi: 10.1128/IAI.00708-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chuquimia OD, Petursdottir DH, Rahman MJ, Hartl K, Singh M, Fernandez C. The role of alveolar epithelial cells in initiating and shaping pulmonary immune responses: communication between innate and adaptive immune systems. PLoS ONE. 2012;7:e32125. doi: 10.1371/journal.pone.0032125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang J, Gigliotti F, Bhagwat SP, Maggirwar SB, Wright TW. Pneumocystis stimulates MCP-1 production by alveolar epithelial cells through a JNK-dependent mechanism. Am J Physiol Lung Cell Mol Physiol. 2007;292:L1495–L1505. doi: 10.1152/ajplung.00452.2006. [DOI] [PubMed] [Google Scholar]
- 51.Manzer R, Wang J, Nishina K, McConville G, Mason RJ. Alveolar epithelial cells secrete chemokines in response to IL-1beta and lipopolysaccharide but not to ozone. Am J Respir Cell Mol Biol. 2006;34:158–166. doi: 10.1165/rcmb.2005-0205OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Christensen PJ, Du M, Moore B, Morris S, Toews GB, Paine R. Expression and functional implications of CCR2 expression on murine alveolar epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2004;286:L68–L72. doi: 10.1152/ajplung.00079.2003. [DOI] [PubMed] [Google Scholar]
- 53.Barrett EG, Johnston C, Oberdörster G, Finkelstein JN. Silica-induced chemokine expression in alveolar type II cells is mediated by TNF-alpha. Am J Physiol Lung Cell Mol Physiol. 1998;275:1110–1119. doi: 10.1152/ajplung.1998.275.6.L1110. [DOI] [PubMed] [Google Scholar]
- 54.Tao F, Kobzik L. Lung macrophage–epithelial cell interactions amplify particle-mediated cytokine release. Am J Respir Cell Mol Biol. 2002;26:499–505. doi: 10.1165/ajrcmb.26.4.4749. [DOI] [PubMed] [Google Scholar]
- 55.Driscoll KE. Macrophage inflammatory proteins: biology and role in pulmonary inflammation. Exp Lung Res. 1994;20:473–490. doi: 10.3109/01902149409031733. [DOI] [PubMed] [Google Scholar]
- 56.O’Brien AD, Standiford TJ, Christensen PJ, Wilcoxen SE, Paine R. Chemotaxis of alveolar macrophages in response to signals derived from alveolar epithelial cells. J Lab Clin Med. 1998;131:417–424. doi: 10.1016/S0022-2143(98)90142-1. [DOI] [PubMed] [Google Scholar]
- 57.Brabcová E, Kolesár L, Thorburn E, Stríz I. Chemokines induced in human respiratory epithelial cells by IL-1 family of cytokines. Folia Biol. 2014;60:180–186. [PubMed] [Google Scholar]
- 58.McIntyre BAS, Kushwah R, Mechael R, Shapovalova Z, Alev C, Bhatia M. Innate immune response of human pluripotent stem cell-derived airway epithelium. Innate Immun. 2015;21:504–511. doi: 10.1177/1753425914551074. [DOI] [PubMed] [Google Scholar]
- 59.Finkelstein JN, Johnston C, Barrett T, Oberdörster G. Particulate-cell interactions and pulmonary cytokine expression. Environ Health Perspect. 1997;105:1179–1182. doi: 10.1289/ehp.97105s51179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Gattas MV, Forteza R, Fragoso MA, Fregien N, Salas P, Salathe M, Conner GE. Oxidative epithelial host defense is regulated by infectious and inflammatory stimuli. Free Radic Biol Med. 2009;47:1450–1458. doi: 10.1016/j.freeradbiomed.2009.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bhattacharya K, El-Sayed R, Andón FT, Mukherjee SP, Gregory J, Li H, et al. Lactoperoxidase-mediated degradation of single-walled carbon nanotubes in the presence of pulmonary surfactant. Carbon. 2015;91:506–517. doi: 10.1016/j.carbon.2015.05.022. [DOI] [Google Scholar]
- 62.Hoffstein ST, Gennaro DE, Manzi RM. Neutrophils may directly synthesize both H2O2 and O2—since surface stimuli induce their release in stimulus-specific ratios. Inflammation. 1985;9:425–437. doi: 10.1007/BF00916342. [DOI] [PubMed] [Google Scholar]
- 63.Fischer H. Mechanisms and function of DUOX in epithelia of the lung. Antioxid Redox Signal. 2009;11:2453–2465. doi: 10.1089/ars.2009.2558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Brandes RP, Weissmann N, Schröder K. Nox family NADPH oxidases: molecular mechanisms of activation. Free Radic Biol Med. 2014;76:208–226. doi: 10.1016/j.freeradbiomed.2014.07.046. [DOI] [PubMed] [Google Scholar]
- 65.van Klaveren RJ, Roelant C, Boogaerts M, Demedts M, Nemery B. Involvement of an NAD(P)H oxidase-like enzyme in superoxide anion and hydrogen peroxide generation by rat type II cells. Thorax. 1997;52:465–471. doi: 10.1136/thx.52.5.465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Nénan S, Boichot E, Lagente V, Bertrand CP. Macrophage elastase (MMP-12): a pro-inflammatory mediator? Mem Inst Oswaldo Cruz. 2005;100(1):167–172. doi: 10.1590/S0074-02762005000900028. [DOI] [PubMed] [Google Scholar]
- 67.Lagente V, Manoury B, Nénan S, Le Quément C, Martin-Chouly C, Boichot E. Role of matrix metalloproteinases in the development of airway inflammation and remodeling. Braz J Med Biol Res. 2005;38:1521–1530. doi: 10.1590/S0100-879X2005001000009. [DOI] [PubMed] [Google Scholar]
- 68.Park E-J, Yoon J, Choi K, Yi J, Park K. Induction of chronic inflammation in mice treated with titanium dioxide nanoparticles by intratracheal instillation. Toxicology. 2009;260:37–46. doi: 10.1016/j.tox.2009.03.005. [DOI] [PubMed] [Google Scholar]
- 69.Shukla A, Barrett TF, Nakayama KI, Nakayama K, Mossman BT, Lounsbury KM. Transcriptional up-regulation of MMP12 and MMP13 by asbestos occurs via a PKCdelta-dependent pathway in murine lung. FASEB J. 2006;20:997–999. doi: 10.1096/fj.05-4554fje. [DOI] [PubMed] [Google Scholar]
- 70.Conner GE, Wijkstrom-Frei C, Randell SH, Fernandez VE, Salathe M. The lactoperoxidase system links anion transport to host defense in cystic fibrosis. FEBS Lett. 2007;581:271–278. doi: 10.1016/j.febslet.2006.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Loza K, Fohring I, Bunger J, Westphal GA, Koller M, Epple M, Sengstock C. Barium sulfate micro- and nanoparticles as bioinert reference material in particle toxicology. Nanotoxicology. 2016 doi: 10.1080/17435390.2016.1235740. [DOI] [PubMed] [Google Scholar]
- 72.Schremmer I, Brik A, Weber DG, Rosenkranz N, Rostek A, Loza K, et al. Kinetics of chemotaxis, cytokine, and chemokine release of NR8383 macrophages after exposure to inflammatory and inert granular insoluble particles. Toxicol Lett. 2016;263:68–75. doi: 10.1016/j.toxlet.2016.08.014. [DOI] [PubMed] [Google Scholar]
- 73.Delaval M, Wohlleben W, Landsiedel R, Baeza-Squiban A, Boland S. Assessment of the oxidative potential of nanoparticles by the cytochrome c assay: assay improvement and development of a high-throughput method to predict the toxicity of nanoparticles. Arch Toxicol. 2017;91:163–177. doi: 10.1007/s00204-016-1701-3. [DOI] [PubMed] [Google Scholar]
- 74.Konduru N, Keller J, Ma-Hock L, Groters S, Landsiedel R, Donaghey TC, et al. Biokinetics and effects of barium sulfate nanoparticles. Part Fibre Toxicol. 2014;11:55. doi: 10.1186/s12989-014-0055-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Celardo I, Pedersen JZ, Traversa E, Ghibelli L. Pharmacological potential of cerium oxide nanoparticles. Nanoscale. 2011;3:1411–1420. doi: 10.1039/c0nr00875c. [DOI] [PubMed] [Google Scholar]
- 76.Koch W. Application of aerosols. In: Uhlig S, Taylor AE, editors. Methods in pulmonary research. Basel: Birkhäuser Basel; 1998. pp. 485–507. [Google Scholar]
- 77.Richards RJ, Davies N, Atkins J, Oreffo VIC. Isolation, biochemical characterization, and culture of lung type II cells of the rat. Lung. 1987;165:143–158. doi: 10.1007/BF02714430. [DOI] [PubMed] [Google Scholar]
- 78.Dobbs LG, Gonzalez R, Williams MC. An improved method for isolating type II cells in high yield and purity. Am Rev Respir Dis. 1986;134:141–145. doi: 10.1164/arrd.1986.134.1.141. [DOI] [PubMed] [Google Scholar]
- 79.Hansen T, Blickwede M, Borlak J. Primary rat alveolar epithelial cells for use in biotransformation and toxicity studies. Toxicol In Vitro. 2006;20:757–766. doi: 10.1016/j.tiv.2005.10.011. [DOI] [PubMed] [Google Scholar]
- 80.Hansen T, Chougule A, Borlak J. Isolation and cultivation of metabolically competent alveolar epithelial cells from A/J mice. Toxicol In Vitro. 2014;28:812–821. doi: 10.1016/j.tiv.2014.03.009. [DOI] [PubMed] [Google Scholar]
- 81.Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative CT method. Nat Protoc. 2008;3:1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 82.Andersen CL, Jensen JL, Ørntoft TF. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004;64:5245–5250. doi: 10.1158/0008-5472.CAN-04-0496. [DOI] [PubMed] [Google Scholar]
- 83.Vandesompele J, de Preter K, Pattyn F, Poppe B, van Roy N, de Paepe A, Speleman F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3:1–11. doi: 10.1186/gb-2002-3-7-research0034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Grubbs FE. Procedures for detecting outlying observations in samples. Technometrics. 1969;11:1–21. doi: 10.1080/00401706.1969.10490657. [DOI] [Google Scholar]
- 85.Henderson RF, Mauderly JL, Pickrell JA, Hahn FF, Muhle H, Rebar AH. Comparative study of bronchoalveolar lavage fluid: effect of species, age, and method of lavage. Exp Lung Res. 1987;13:329–342. doi: 10.3109/01902148709069597. [DOI] [PubMed] [Google Scholar]
- 86.Kittel B, Ruehl-Fehlert C, Morawietz G, Klapwijk J, Elwell MR, Lenz B, et al. Revised guides for organ sampling and trimming in rats and mice—part 2. A joint publication of the RITA and NACAD groups. Exp Toxicol Pathol. 2004;55:413–431. doi: 10.1078/0940-2993-00349. [DOI] [PubMed] [Google Scholar]
- 87.Rittinghausen S, Bellmann B, Creutzenberg O, Ernst H, Kolling A, Mangelsdorf I, et al. Evaluation of immunohistochemical markers to detect the genotoxic mode of action of fine and ultrafine dusts in rat lungs. Toxicology. 2013;303:177–186. doi: 10.1016/j.tox.2012.11.007. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1. Fold regulation of regulated genes. List of fold regulation and standard deviation values of the regulated genes listed in Tables 2 and 3 of the manuscript. Values are shown for every dose group and time point.
Additional file 2: Table S2. Analyzed genes per array. List of all analyzed genes (short and long name) separated per PCR profiler array.
Data Availability Statement
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.