
Figure 3.
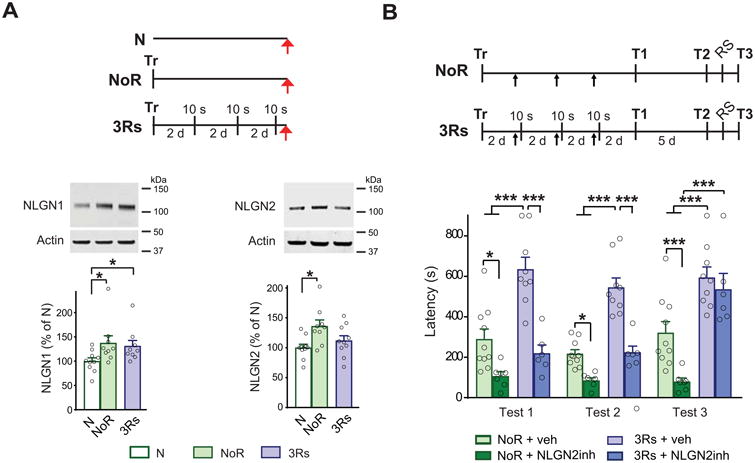
Neuroligin 1 (NLGN1) and NLGN2 play distinct roles in memory consolidation, strengthening, and extinction inhibition. (A) Schema of behavioral procedures, representative images and relative quantitative western blot analyses of PL cortex. Protein extracts were obtained from rats trained (Tr) in inhibitory avoidance (IA) and given 3 brief memory retrievals (3Rs), which consisted of 10 sec exposures to the training context without footshock, every two days. Rats were euthanized (red arrows) one hour after 3Rs or at the matched time point for the group that underwent training without retrievals and remained in the home cage after training (NoR). Naïve rats (N) served as reference controls. Data are presented as mean percentage ± s.e.m. of the mean values of the N group. One-way ANOVA followed by Newman-Keuls post hoc test; NLGN1 n = 9–10, NLGN2 n = 9–10; *p < 0.05 for both comparisons. (B) Schema of behavioral procedures is given above the graphs. Rats were trained (Tr) in IA and given 3Rs every two days or left in the home cage without retrieval (NoR) after training. Thirty minutes before each reactivation, or at matched timepoints in the NoR group, the animals received a bilateral PL cortex injection (black arrows) of NLGN2 recombinant extracellular domain (NLGN2inh) to inhibit NLGN2 function. Animals were tested for memory retention two days after the last retrieval (T1), and again five days later (T2), as shown in the schema. A reminder footshock (RS) was given in a different context with the same shock intensity one day after T2 and memory was tested one day later (T3). Data are expressed as mean latency ± s.e.m. Two-way ANOVA followed by Bonferroni post hoc test; *p<0.05, ***p<0.001 n = 7-10 (Adapted from Ye et al., 2017 [11]).