

Abstract
Background
The mosquito resistance to the insecticides threatens malaria control efforts, potentially becoming a major public health issue. Alternative methods like ivermectin (IVM) administration to humans has been suggested as a possible vector control to reduce Plasmodium transmission. Anopheles aquasalis and Anopheles darlingi are competent vectors for Plasmodium vivax, and they have been responsible for various malaria outbreaks in the coast of Brazil and the Amazon Region of South America.
Methods
To determine the IVM susceptibility against P. vivax in An. aquasalis and An. darlingi, ivermectin were mixed in P. vivax infected blood: (1) Powdered IVM at four concentrations (0, 5, 10, 20 or 40 ng/mL). (2) Plasma (0 hours, 4 hours, 1 day, 5, 10 and 14 days) was collected from healthy volunteers after to administer a single oral dose of IVM (200 μg/kg) (3) Mosquitoes infected with P. vivax and after 4 days was provided with IVM plasma collected 4 hours post-treatment (4) P. vivax-infected patients were treated with various combinations of IVM, chloroquine, and primaquine and plasma or whole blood was collected at 4 hours. Seven days after the infective blood meal, mosquitoes were dissected to evaluate oocyst presence. Additionally, the ex vivo effects of IVM against asexual blood-stage P. vivax was evaluated.
Results
IVM significantly reduced the prevalence of An. aquasalis that developed oocysts in 10 to 40 ng/mL pIVM concentrations and plasma 4 hours, 1 day and 5 days. In An. darlingi to 4 hours and 1 day. The An. aquasalis mortality was expressively increased in pIVM (40ng/mL) and plasma 4 hours, 1, 5 10 and 14 days post-intake drug and in An. darlingi only to 4 hours and 1 day. The double fed meal with mIVM by the mosquitoes has a considerable impact on the proportion of infected mosquitoes for 7 days post-feeding. The oocyst infection prevalence and intensity were notably reduced when mosquitoes ingested blood from P. vivax patients that ingested IVM+CQ, PQ+CQ and IVM+PQ+CQ. P. vivax asexual development was considerably inhibited by mIVM at four-fold dilutions.
Conclusion
In conclusion, whole blood spiked with IVM reduced the infection rate of P. vivax in An. aquasalis and An. darlingi, and increased the mortality of mosquitoes. Plasma from healthy volunteers after IVM administration affect asexual P. vivax development. These findings support that ivermectin may be used to decrease P. vivax transmission.
Author summary
Malaria is one of the most important infectious diseases in the world with hundreds of millions of new cases every year. The disease is caused by parasites of the genus Plasmodium where Plasmodium vivax represent most of the cases in the Americas. Current strategies to combat malaria transmission are being implemented; however, widespread insecticide resistance in vectors threatens the effectiveness of vector control programs. Ivermectin (IVM) has arisen as a new potential tool to be added to these programs as it has mosquito-lethal and sporontocidal properties making it a promising transmission reduction drug. Plasmodium vivax was drawn from patients, mixed with powdered IVM and metabolized IVM in plasma collected from healthy volunteers receiving IVM, and fed to mosquitoes via membrane feeding. Powdered and metabolized IVM interrupt P. vivax transmission, reducing oocyst infection and intensity rate of two South American malaria vectors An. aquasalis and An. darlingi. We also demonstrate the effect of IVM on asexual stages development of P. vivax, providing evidence that IVM may affect different parasite life cycle stages. Our findings place IVM as a strong candidate for malaria transmission reducing interventions.
Introduction
The 2016 World Malaria Report (WHO) estimated 212 million cases of malaria worldwide, leading to 429,000 deaths, which illustrates that malaria remains an important public health problem. In the Americas, 389,390 cases and 87 deaths were reported in 2016 with Brazil reporting 24% and Peru 19% of the malaria cases [1]. The majority (92.5%) of the malaria cases occurred in the Amazon sub-region with 69% being Plasmodium vivax [1–4]. Despite considerable efforts, the majority of South American countries are still far from achieving vivax malaria elimination. Current strategies to combat malaria transmission in South America include diagnosis and treatment with artemisinin-based combination therapy (ACT) [5–7] and long-lasting insecticidal nets [8], supported by indoor-residual spraying of insecticide IRS [9, 10]. However, widespread insecticide resistance in vectors threatens the effectiveness of LLINs and IRS [1–15].
Anopheles aquasalis and Anopheles darlingi are known to be susceptible to P. vivax infection ([16–18]). Anopheles aquasalis is considered the primary vector in coastal areas of Central and South America and has been used as a neotropical anopheline vector for laboratory model to evaluate host-parasite interactions [18–20]. Anopheles darlingi is the primary vector in the Amazonian region of South America [21, 22].
The reemergence of attention on transmission blocking strategies for Plasmodium [23, 24] have raised research efforts in attempt to find vaccines [25–27], drugs [28, 29] or microorganisms [30–32] able to disrupt the life cycle of the parasite in the mosquito vector. In this context, the endectocide ivermectin (IVM) has arisen as a new promising tool to be added to malaria control programs.
Ivermectin is a safe drug with activity against a wide range of internal and external parasites and it is used widely in both veterinary and human medicine [33]. Ivermectin as a single oral dose (150–200 μg/kg) is effective for the treatment or control of Onchocerca volvulus, Wuchereria bancrofti and Strongyloides stercoralis. Ivermectin has been widely distributed to humans via mass drug administration (MDA) campaigns against onchocerciasis and lymphatic filariasis in Africa and Latin America [34–36]. Ivermectin has a secondary effect on ectoparasites that feed on recently treated individuals [37], including activity against Anopheles vectors at concentrations present in human blood after standard doses [38, 39]. Consequently, IVM has emerged as a potential tool for malaria control [40, 41]. Ivermectin MDA provides a unique insecticide dissemination route via mosquito ingestion of the compound through a blood meal rather than physical contact as most insecticides are delivered. In Anopheles vectors, IVM acts as an agonist of glutamate-gated chloride channels, causing flaccid paralysis and eventual death [42]. Both in vitro and in vivo studies have shown that a blood meal containing IVM cause a significant reduction in the adult Anopheles lifespan, with secondary sub-lethal effects delaying time to re-feed [38, 43], a sporontocidal effect [38, 44, 45], reductions in fecundity [39, 46, 47] and egg hatch rate [39], and even reduced locomotor activity [48].
Field studies of the effect of MDA with IVM on malaria transmission showed that a single dose of 150–200 μg/kg reduced the survivorship of wild Anopheles gambiae, with an accompanying reduction in the Plasmodium falciparum sporozoite rate [9, 49, 50]. These studies demonstrated that IVM could be a potential additional tool for malaria control.
Several studies have demonstrated that powdered IVM p(IVM) inhibits Plasmodium development in the vector, including P. falciparum in An. gambiae [44, 45] and more recently P. vivax in Anopheles dirus and Anopheles minimus [38] and An. darlingi [43]. Recent evidence suggests the presence of long-lived IVM metabolites that may have mosquito-lethal activity [38]. Thus, it is critical to determine the sporontocidal effect of IVM and potential metabolites in human plasma/blood after administration of IVM at clinically relevant doses.
Ivermectin has also been shown to inhibit the development of asexual blood stages of P. falciparum in vitro and Plasmodium berghei in vivo [51]. Additionally, it has been reported that IVM has liver stage inhibition in a P. berghei rodent model [52]. It remains to be determined if these blood and liver stage effects of IVM have a prophylactic personal protective effect for a human.
In this study, for the first time, we evaluated the IVM effect and its possible metabolites against P. vivax in An. aquasalis and An. darlingi. Additionally, we investigated the potential effects of IVM against asexual blood stages of P. vivax. A better understanding of the effects of IVM and possible metabolites on parasite maturation and transmission to mosquito vectors would contribute to the development of IVM for malaria control strategies.
Methods
Ethics statement
The procedures were approved by the Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (FMT-HVD) Ethics Review Board (ERB) (Approval Number: 296.723 CAAE: 14148813.7.0000.0005) and by the U.S. Naval Medical Research Unit No. 6 (NAMRU-6) and Walter Reed Army Institute of Research Institutional Review Boards (NMRCD.2008.0004 and WRAIR#2175). All subjects provided and signed written informed consent.
P. vivax malaria patients, healthy volunteer recruitment
This study was conducted at FMT-HVD, a tertiary care center for infectious diseases in Manaus, Amazonas State, Brazil and NAMRU-6 in Iquitos (Loreto), Peru. In order to obtain P. vivax samples, adult patients (ages ≥18 years) infected with P. vivax were recruited from the areas surrounding Manaus and Iquitos. P. vivax infection was determined by light microscopy of Giemsa stained blood samples. In Brazil, P. vivax infected patients were identified at FMT-HVD, enrolled and venous blood (15 ml) was collected. In Peru, P. vivax infected patients were identified at Ministry of Health Centers and hospitals in Iquitos, transported to NAMRU-6, enrolled and venous blood (15 ml) was drawn on site for the ivermectin sporogony experiments. The age, gender, history of previous malaria episodes, and place of residence were obtained from the patients and it was verified no signs of severe disease and no previous antimalarial treatment during the preceding 4 weeks. After blood collection, all patients were treated for P. vivax infection following guidelines of the Brazilian Health Ministry or Peruvian Ministry of Health [53].
In order to obtain metabolized (mIVM) plasma samples, healthy volunteers were recruited in Manaus, Brazil. Inclusion criteria were: adults ≥18 years old, history of at least 3 month-negative malaria, and were confirmed as healthy volunteers by health works.
Mosquito colonies
Anopheles aquasalis were reared at Laboratory of Medical Entomology at FMT-HVD in Manaus, Brazil and An. darlingi were reared at NAMRU-6 in Iquitos, Peru. These colonies were kept at a constant temperature (24–26°C) and relative humidity (70–80%) on a 12:12 light-dark cycle. Larvae were hatched in room temperature water and fed ground TetraMin fish food for An. aquasalis and rodent diet for An. darlingi provided daily. The larvae were allowed to pupate and emerge into adults in an enclosed mesh-covered cage with water and 10% sucrose available [18, 54]. Adult mosquitoes used for experiments were between 3–5 days post-emergence.
Drugs and treatment regimens
Powdered pIVM and powdered chloroquine (pCQ) reference material were obtained from Sigma Aldrich (St. Louis, MO, USA). pIVM was dissolved in dimethyl sulfoxide (DMSO) to a concentration of 10 mg/ml and pCQ in Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma Aldrich, St. Louis, MO, USA) to a concentration of 1 mg/ml and aliquots were frozen at -20°C. IVM was serially diluted in PBS to achieve experimental concentrations.
All experimental drug regimens were managed at the FMT-HVD in Manaus, Brazil. IVM tablets (Abbot Laboratórios do Brasil, State, Brazil) were administered at a single dose of 200 μg/kg. Vivax patients received chloroquine (CQ) tablets (Farmaguinhos Laboratórios do Brasil, Rio de Janeiro State, Brazil), administered as a daily dose for three days (i.e., 600 mg in the first day and 450 mg in the second and third day). Primaquine (PQ) tablets (Med Pharma, São Paulo State, Brazil) were administered as a daily dose of 30 mg for 7 days following the Brazilian MoH guideline for vivax malaria treatment [53].
To evaluate the effect of mIVM on P. vivax development, five volunteers, all healthy men from 18 to 50 years, were recruited for the experiments. They received one single dose (200 μg/kg) of IVM. Blood was collected in heparinized tubes at 0 and 4 hours, and 1, 5, 10 and 14 days after drug intake.
For the evaluation of the in vivo mIVM effect on P. vivax, 15 patients with confirmed P. vivax malaria infection were recruited. The patients were divided into four groups and different treatment regimens were provided: (1) IVM plus CQ, (2) CQ alone, (3) PQ plus CQ and (4) IVM plus PQ plus CQ. Before and after 4 hours of drug intake, blood samples were collected. The patients that receive IVM plus CQ or CQ alone have received the first PQ dose after blood was drawn at 4 hours past CQ intake. All patients were treated with PQ and CQ dosage following the Brazilian Ministry of Health Guidelines.
Transmission blocking experimental design
Four experiments were performed to determine the effect of IVM on P. vivax in either An. aquasalis or An. darlingi (Fig 1):
Fig 1. Schematic outline of the experiments of membrane feeding.

Four experiments were performed to determine the transmission blocking potential of IVM in either An. aquasalis or An. darlingi. Abbreviations: Ivermectin (IVM), powdered ivermectin (pIVM), metabolized ivermectin (mIVM), chloroquine (CQ), primaquine (PQ).
Powdered ivermectin experiment (pIVM)—P. vivax infected blood was maintained at 37°C until centrifuged at 1,500 RPM for 5 minutes at 37°C. The plasma was removed and packed red blood cells (RBCs) were washed with RPMI 1640 medium, repeated twice, and reconstituted to 40% hematocrit with non-immune human AB serum (Sigma Aldrich, MO, USA) and mixed with pIVM to achieve final blood concentrations of 5, 10, 20 or 40 ng/mL and a control (0 ng/ml). These concentrations were chosen based on the human pharmacokinetic curve for IVM, corresponding approximately to the concentration founds 4 hours to 60 hours post ingestion [33, 55]. These experiments were only performed with An. aquasalis at FMT-HVD.
Metabolized ivermectin (mIVM) experiment—P. vivax infected blood meal was mixed with plasma from the healthy volunteers administered ivermectin. Ivermectin-treated blood was collected at 0 and 4 hours and 1, 5, 10 and 14 days after administration. Ivermectin-treated blood from the healthy volunteers was centrifuged at 1,500 RPM for 5 min and plasma was removed and stored frozen at -20°C. These experiments were performed with An. aquasalis at FMT-HVD and An. darlingi at NAMRU-6.
Metabolized ivermectin (mIVM) double feed experiment—Mosquitoes were infected with P. vivax and then four days later were blood fed mIVM collected 0 hours (control) or 4 hours after drug administration. These experiments were performed with An. aquasalis at FMT-HVD and An. darlingi at NAMRU-6.
Metabolized drug in vivo experiment—P. vivax infected patients were treated with the following drug regimens described above: IVM+CQ, CQ alone, PQ+CQ and IVM+PQ+CQ. To elucidate the sporontocidal effect of IVM, CQ, and PQ after in vivo exposure in humans, three different blood meal preparations were made from the same patient: reconstituted, unprocessed, and control. For reconstituted blood preparations, P. vivax infected blood was collected just before drug administration, held at 37°C for 4 hours and the blood was reconstituted with drug-treated plasma from the same patient collected 4 hours later. The 4th hour drug-treated blood samples were centrifuged at 1,500 RPM for 5 minutes at 37°C. The 0th hour P. vivax blood was centrifuged, washed with RPMI and reconstituted to a 40% hematocrit with the 4 hours drug-treated plasma and fed to mosquitoes. For unprocessed blood preparations, the blood was collected 4 hours post drug intake and no manipulation of the blood sample was performed prior to feeding to mosquitoes. For control blood preparations, the blood was collected at 0 hours before drug intake and no manipulation of the blood sample was performed prior to feeding to mosquitoes. These experiments were only performed with An. aquasalis at FMT-HVD.
P. vivax infection assessment in mosquitoes
P. vivax infected blood from patients was prepared as described above. The plasma was removed and packed red blood cells (RBCs) were washed with RPMI 1640 medium, repeated twice, and reconstituted to 40% hematocrit with non-immune human AB serum (experiment 1: pIVM) or with plasma from drug-treated volunteers or patients (experiments 2–4: mIVM, double feed mIVM, and in vivo mIVM). Adult female mosquitoes were sugar starved overnight prior to infection via membrane feeding assay (MFA). Blood meals (1ml) prepared as described above were offered to groups of at least 100 mosquitoes for 30 minutes via membrane feeder devices at 37°C as described in detail elsewhere [18, 20]. The fully engorged mosquitoes were separated into different cages and kept until 7 or 14 days post-feeding. Mosquito mortality was monitored on day 7 or 14.
Seven days after P. vivax blood meal ingestion midguts from all experimentally infected mosquito groups were dissected in PBS under a stereo microscope. The midguts were stained with 0.1% commercial Mercurochrome (Merbromin, Sigma- Aldrich, USA), placed under a cover slip and examined for the presence of oocysts with a compound microscope (Optical Microscopy, Olympus, Germany). Infection prevalence was expressed as percentage of mosquitoes with at least one oocyst. Infection intensity was determined as the arithmetic mean of oocysts counted per dissected midgut.
Measurement of ivermectin from human plasma
Plasma samples from healthy volunteers and patients were shipped on dry-ice to the Department of Clinical Pharmacology, Mahidol-Oxford Tropical Medicine Research Unit, Bangkok, Thailand for IVM drug concentration measurements. Plasma concentrations of IVM were determined by a newly developed and validated method using solid-phase extraction and liquid chromatography with tandem mass spectrometry (manuscript in preparation). The linear range for quantification was 0.97–384 ng/m. Three replicates of quality control samples at low, middle, and high concentrations were included in the analysis to ensure precision and accuracy. The observed total assay coefficient of variation was <10% in all quality control samples in accordance with US Food and Drug Administration requirements [56].
Ex vivo schizont maturation inhibition assay
The P. vivax schizont maturation assay was performed as previously described [57] with five P. vivax samples from Manaus, Brazil. The parasitemia was determined by counting the number of parasites per 200 leukocytes. The plasma and buffy coat were separated from RBCs by centrifugation and packed RBCs were washed with RPMI and this was repeated twice. Leukocytes were removed by passing samples in a cellulose column [58] and afterward, the RBC pellet was suspended in McCoy 5A medium supplemented with 3.2% glucose and 20% human AB serum.
Drug plates were freshly prepared to avoid drug degradation. The pIVM and pCQ stock solutions were subsequently diluted in McCoy’s medium to obtain 9 serial dilutions of the drug (1000–3.9 ng/ml). To determine IC50s, 50 μl of pIVM and pCQ solutions were added into 3 wells of 96 well plates, and 9 serial dilutions of each drug were done in triplicate. Three wells were free of drug and served as a control. Additionally, 50 μl of plasma samples from each IVM-treated healthy volunteer 0 and 4 hours after IVM administration were added at 4 different dilutions (1:2, 1:4, 1:8 and 1:16) in triplicate. Then 50 μl of parasite solution was added to each well to achieve a 4% hematocrit. The plates were incubated in a modular incubator chamber containing 5% CO2, 5% O2, and 90% N2 at 37°C. Incubation was stopped when at least 40% of parasites on the ring stage had matured to schizonts in the drug-free control wells.
After incubation, the plates were allowed to stand for 30 minutes in a semi-vertical position. The supernatant was removed, erythrocytes suspended in the remaining fluid, and a thick blood film was made from each well. Thick blood films were stained with Giemsa stain. The number of schizonts containing more than three nuclei per 200 asexual stage parasites was determined in each blood film. Data for analyses prepared as follows: Schizont maturation in relation to control (%) = 100 x (number of schizonts in treated well/number of schizonts in control wells).
Data analysis
Data were entered in Prism 7.03 (GraphPad, USA) and subsequently analyzed by Stata 11.2 (Data Analysis and Statistical Software, Texas, USA). Mosquito oocyst prevalence and schizont proportions were compared by one-way ANOVA and post-hoc analysis using paired T-test to compare each concentration with respect to the control. Mosquito oocyst intensity was compared by Kruskal-Wallis test and post-hoc analysis using Kolmogorov-Smirnov to compare each concentration with respect to the control. P-values <0.05 were considered statistically significant. The P. vivax mosquito-stage oocyst inhibition in An. Aquasalis and An. darlingi after ingestion of whole blood spiked with IVM or whole blood equivalent from volunteers administered ivermectin were analyzed using GraphPad Prism v7.02. Normalized oocyst inhibition versus human volunteer plasma concentrations of IVM was analyzed using a nonlinear dose-response analysis with a variable slope. The maximum inhibition was fixed to 100%, and the minimum inhibition (zero drug concentration) and the drug concentration producing 50% of maximum effect (IC50) was estimated. The P. vivax asexual blood-stage inhibitory concentrations (i.e. IC50, IC90 and IC99) for pIVM and pCQ were determined using the free software ICE estimator available online at http://www.antimalarial-icestimator.net/.
Results
Plasma ivermectin results
Median peak plasma concentrations (i.e., plasma samples collected 4 hours post-dose) was 79.8 (50.6–112) ng/mL after a single oral dose of 200 μg/kg of IVM. Only two out of five volunteers had detectable IVM plasma concentrations 10 days post-dose (i.e., 2.64 and 2.32 ng/mL), while only one volunteer had a detectable IVM plasma concentration on day 14 after dose (i.e., 1.87 ng/mL) (S1 Table).
Powdered Ivermectin (pIVM) inhibits P. vivax development in An. aquasalis
Different concentrations of pIVM were evaluated (5, 10, 20 and 40 ng/mL) on the P. vivax infection prevalence, oocyst intensity, and An. aquasalis mortality (Fig 2A–2C, S1 Dataset). There were significant differences among pIVM treatment groups for infection prevalence [F (3.23) = 11.58, p = 0.001] compared to the control (0ng/ml) group. P. vivax infection prevalence was significantly reduced in mosquitoes that ingested pIVM at 10 ng/ml by 33.2% [32.03% (SD = 5.83%), p = 0.0051, reps = 7, n = 171], 20 ng/ml by 33.7% [31.81% (SD = 5.74%), p = 0.019, reps = 7, n = 68] and 40 ng/ml by 61.3% [18.60% (SD = 5.81%), p<0.0001, reps = 7, n = 178) concentrations but not the 5ng/ml by 12.6% [41.93% (SD = 7.67%), p = 0.141, reps = 7, n = 160) concentration (Fig 2A). Also, the infection intensity (i.e. number of oocysts per mosquito) was reduced in the groups of mosquitoes that fed on infective blood meals containing 10ng/mL by 62.6% [5.89 (SD = 5.87), p = 0.0079, reps = 7, n = 171), 20ng/mL by 72.6% [4.32 (SD = 2.71), p = 0.0298, reps = 7, n = 68] and 40ng/mL by 86.1% [2.20 (SD = 0.97), p<0.0001, reps = 7, n = 178) but not the 5ng/ml by 56.6% [6.84 (SD = 3.20), p = 0.99, reps = 7, n = 160) concentration of pIVM in comparison to the control group (Fig 2B) [15.73 (SD = 12.97), reps = 7, n = 134]. There were significant differences among pIVM treatment groups for mosquito mortality at 7-day [F (3.23) = 2.99, p = 0.052] compared to the control (0ng/ml) group but the mortality rate was higher only in the 40 ng/ml group [78.11% (SD = 7.17%), p = 0.0007] but not 20 ng/ml group [66.34% (SD = 9.72%), p = 0.056], 10 ng/ml group [49.09% (SD = 20.03%), p = 0.646] or 5ng/ml group [63.38% (SD = 13.26%), p = 0.080] compared to the control group [43.93% (SD = 14.46%)] (Fig 2C).
Fig 2. Ivermectin powdered affects Plasmodium vivax development in Anopheles aquasalis.
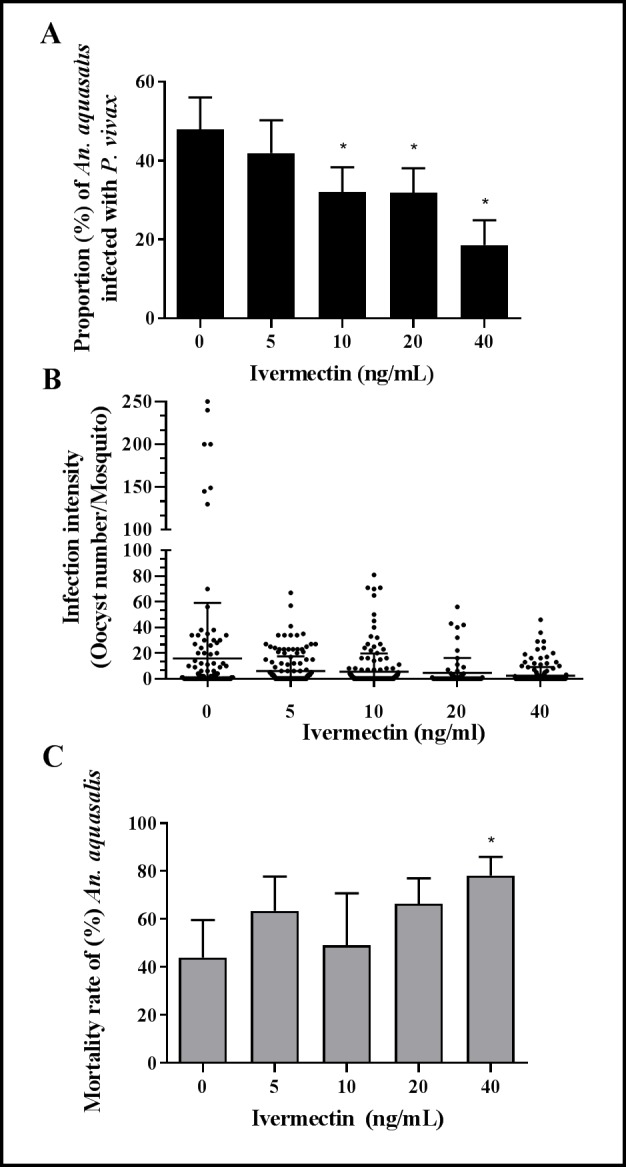
(A) The proportion of P. vivax infected mosquitoes is presented as a mean and standard deviation. (B) The infection intensity is presented as the number of oocysts per a single midgut (black dots) and the black lines represent the mean and standard deviation. (C) Mosquito mortality rate to 7 days post-feeding is presented as mean and standard deviation. Data from 5 independent experiments are presented in A-C. Asterisks (*) represent significant difference (P < 0.05) in relation to the control.
Metabolized ivermectin (mIVM) inhibits P. vivax development in An. aquasalis and An. darlingi
The ingestion of mIVM presented in the infective blood meals affected the infection rate and the infection intensity and the mortality of the vectors An. aquasalis and An. darlingi as seen in the Fig 3A–3F. There were significant differences for among mIVM treatment groups for infection prevalence [F (3.31) = 119.02, p<0.001] in An. aquasalis and [F (2.9) = 9.39, p = 0.0063] in An. darlingi compared to the control (0 hour) group. The oocyst infection rates were reduced in the groups of An. aquasalis fed volunteer mIVM plasma collected at 4 hours (48.0 ng/ml) by 89.2% [7.82% (SD = 7.04%), p<0.0001, reps = 10, n = 232), 1 day (5.55 ng/ml) by 51.5% [35.11% (SD = 8.96%), p = 0.0038, reps = 6, n = 102], and 5 days (1.58 ng/ml) by 24.2% [54.88% (SD = 8.57%), p = 0.0009, reps = 9, n = 300) but not 10 days by 14.3% [62.10% (SD = 9.46%), p = 0.063, reps = 9, n = 258) or 14 days by 2.3% [70.75% (SD = 7.57%), p = 0.073, reps = 9, n = 214) (Fig 3A) compared to the mIVM control [72.38% (SD = 6.05%), reps = 10, n = 448]. The oocyst infection rates were reduced in the groups of An. darlingi fed mIVM plasma collected at 4 hours (43.24 ng/ml) by 91.1% [6.25% (SD = 10.8%), p = 0.019, reps = 4, n = 8] and 1 day (5.69 ng/ml) by 73.9% [18.33% (SD = 18.48%), p = 0.014, reps = 4, n = 11], but increased slightly but not significantly on 5 days (1.35 ng/ml) by 1.95% [71.47% (SD = 22.88%), p = 0.156, reps = 4, n = 67], 10 days (2.48 ng/mL) by 0.48% [70.44% (SD = 28.17%), p = 0.098, reps = 4, n = 97], or 14 days (1.87 ng/mL) by 14.89% [80.54% (SD = 7.5%), p = 0.350, reps = 4, n = 51] compared with the control [70.1% (SD = 25.4%), reps = 4, n = 97] (Fig 3B). The mIVM concentrations imbibed by the mosquitoes were calculated by taking pharmacokinetic values and multiplying by 60% to account for the 40% hematocrit in each blood meal, several volunteers had no detectable ivermectin at days 10 and 14 so no concentrations ingested by mosquitoes could be estimated. We did not find significant differences for infection prevalence among both species in each treatment group: Control (p = 0.9178), 4 horas (p = 0.527), 1 day (0.287), 5 days (0.2883), 10 days (p = 0.734) and 14 days (p = 0.513). However, the mean oocyst intensity for An. darlingi (Fig 3D) is substantially higher than An. aquasalis (Fig 3C).
Fig 3. Ivermectin metabolized affects Plasmodium vivax development in Anopheles aquasalis and Anopheles darlingi.

An. aquasalis (A,C,E) and An. darlingi (B,D,F) mosquitoes were fed P. vivax with mIVM collected at various time points (0 and 4 hours; 1, 5, 10 and 14 days). (A) and (B) Proportion of P. vivax infected mosquitoes are presented as mean and standard deviation. (C) and (D) The intensity of infection is presented as the number of oocysts per a single midgut (black dots) and the black lines represent the median and standard deviation. (E) and (F) The mosquito mortality rate is presented as mean and standard deviation. Data from 5 independent experiments are presented. Asterisks (*) represent significant difference (P < 0.05) in relation to the control.
On the other hand, the oocyst infection intensities were reduced in all the groups: plasma 4 hours (43.24 ng/ml) by 96.1% [0.78 (SD = 2.03), p<0.0001, reps = 6, n = 232], 1 day (5.69 ng/ml) by 56.8% [8.52 (SD = 10.09), p<0.0001, reps = 6, n = 102], 5 days (1.35 ng/ml) by 29.4% [13.91 (SD = 11.39), p<0.0001, reps = 6, n = 300], 10 days by 41.3% [11.57 (SD = 5.50), p = 0.008, reps = 6, n = 258] and 14 days by 58.2% [8.24 (SD = 2.55), p = 0.007, reps = 6, n = 214] after the volunteer pIVM intakes compared with the control [19.70 (SD = 13.09), reps = 6, n = 448] in An. aquasalis (Fig 3C) and in plasma 4 hours (43.24 ng/ml) by 99.9% [0.06 (SD = 0.10), p = 0.003, reps = 4, n = 8], 1 day (5.69 ng/ml) by 97.57% [2.31 (SD = 3.77), p = 0.008, reps = 4, n = 11] and 5 days (1.35 ng/ml) by 56.8% [76.49 (SD = 80.96), p = 0.011, reps = 6, n = 67], but not 10 days by 56.8% [99.77 (SD = 125.9), p = 0.593, reps = 6, n = 97] and 14 days by 56.8% [37.80 (SD = 5.1), p = 0.141, reps = 6, n = 51] after the volunteer pIVM intakes compared with the control [94.8, (SD = 90.18), reps = 4, n = 97] in An. darlingi (Fig 3D). It is important to highlight that An. darlingi showed much less oocyst number on 4 hours and 1 day than An. aquasalis.
At day 7 post blood meal, there were significant differences in the mortality rate among mIVM treatment groups in An. aquasalis [F (5.46) = 16.45, p<0.0001] and mIVM treatment groups in An. darlingi [F (2.9) = 16.92, p<0.0001] compared to the control groups (plasma 0 hours). The mosquito mortality was significant higher 4 hours [78.33% (SD = 17.73%), p = 0.0006], 1 day [60.15% (SD = 6.95%), p = 0.0017], 5 days [36.84% (SD = 11.99%), p = 0.046], 10 days [50.01% (SD = 11.44%), p = 0.014] and 14 days [39.83% (SD = 17.58%), p = 0.043] (Fig 3E) than the control group [21.29% (SD = 15.80%)] in An. aquasalis. However, at day 7 post blood meal only the An. darlingi groups feds on infective blood meals containing plasma collected 4 hours [97.43% (SD = 2.56%), p = 0.027] and 1 day [97.07% (SD = 3.23%), p = 0.025] had significantly increased mortality rates but not when fed plasma from 5 days [71.23% (SD = 14.91%), p = 0.144], 10 days [40.25% (SD = 7.47%), p = 0.855] or 14 days [31.4% (SD = 1.9%), p = 0.873] compared with the control [43% (SD = 22.45%] (Fig 3F).
At 14 days post blood meal, after mIVM plasma from 4 hours, 1 and 5 days intake all the An. darlingi had died, therefore sporozoite prevalence at these time points could not be analyzed. Also, there were no significant differences on the mortality rate with respect to control [36.83% (SD = 20.06%)] in the groups that were blood fed mIVM of 10 days [57.66% (SD = 15.21%), p = 0.265] and 14 days [48.65% (SD = 21.67%), p = 0.200]. An. darlingi sporozoite prevalence at day 14 post blood meal was not reduced with respect to control [90.27% (SD = 13.39%)] in the groups that were blood fed mIVM from 10 days [90.38% (SD = 10.88%), p = 0.992] or 14 days [82.14% (SD = 1.68%), p = 0.824].
Interestingly, the mIVM (IC50 = 5.68 [3.79–7.56] ng/ml [95%CI]) of mosquito-stage P. vivax in An. aquasalis appears to be much lower compared to pIVM (IC50 = 28.15 [16.36–39.94] ng/ml) (Fig 4A and 4B). This demonstrates that mIVM has a more potent sporontocidal effect against P. vivax compared to ivermectin compound.
Fig 4. Activity of IVM on P. vivax infections in An. aquasalis.

P. vivax oocyst inhibition in An. aquasalis when ingesting plasma from human volunteers receiving standard single oral dosing of IVM (A) or whole blood spiked with IVM (B).
mIVM impairs P. vivax development when ingested 4 days post parasites in An. aquasalis and An. darlinigi
Anopheles darlingi and An. aquasalis had significantly reduced oocyst infection prevalence and intensity when ingested a second mIVM (4 hours) non-infective bloodmeal 4 days post P. vivax infection compared to the control. An. aquasalis oocyst prevalence was reduced by 84.5% [10.72% (SD = 6.79%), p = 0.047, reps = 4, n = 50] compared to the control [68.83% (SD = 10.69%), reps = 4, n = 94] (Fig 5A) and An. darlingi by 60.3% [33.98% (SD = 17.8%), p<0.0001, reps = 3, n = 31] compared to the control [85.49% (SD = 13.2%), reps = 3, n = 73] (Fig 5B). Oocyst intensity in An. aquasalis was reduced by 93.6% [0.94 (SD = 0.67), p<0.0001, reps = 4, n = 50] compared to the control [14.65 (SD = 7.71), reps = 4, n = 94] (Fig 5C) and in An. darlingi by 97% [0.73 (SD = 0.2), p<0.0001, reps = 3, n = 31] compared to the control [24.0 (SD = 53.3), reps = 3, n = 73] (Fig 5D).
Fig 5. Ivermectin impairs Plasmodium vivax development in Anopheles aquasalis and Anopheles darlingi when administered in a double feed 4 days after infection.

An. aquasalis and An. darlingi female mosquitos were fed P. vivax blood samples. Four days after infection, two groups of mosquitos received a second uninfected blood meal with mIVM plasma from control (0 hours) or 4 hours after IVM intake. (A) The proportion of P. vivax infected An. aquasalis and (B) An. darlingi are presented as the mean and standard deviation. (C and D) The intensity of infection is presented as the number of oocysts per midgut (dots) and the black lines represent the median and standard error. Data from 5 independent experiments are presented. Asterisks (*) represent significant difference (P < 0.05) in relation to control.
mIVM and metabolized malaria compounds inhibit P. vivax development in An. aquasalis
There were significant reductions in oocyst infection prevalence and intensity compared to control when An. aquasalis ingested the drug-treated infected blood meals with unprocessed [F (3.17) = 159.90, p<0.0001] and reconstituted [F (3.17) = 164.82, p<0.0001] blood. For unprocessed blood, IVM+CQ by 63.7% [29.44% (SD = 4.19%), p = 0.029, reps = 3, n = 64]; PQ+CQ by 71.1% [23.44% (SD = 7.95%), p = 0.02, reps = 3, n = 161] and IVM+PQ+CQ by 66.5% [27.15% (SD = 2.20%), p = 0.046, reps = 3, n = 99] but not CQ alone by 26.5% [59.49% (SD = 10.9%), p = 0.072, reps = 3, n = 58] compared with the control [80.91% (SD = 4.12%), reps = 3, n = 615], (Fig 6A). Similar to observed in mosquitos fed with unprocessed blood, the infection rate was significantly reduced on mosquitos fed with P. vivax blood meal reconstituted with plasma from patients that undertook IVM+CQ by 65.6% [27.86% (SD = 4.44%), p = 0.006, reps = 3, n = 205], PQ+CQ 56.55% [35.21% (SD = 4.17%), p = 0.004, reps = 3, n = 180] and IVM+PQ+CQ 62.9% [30.02% (SD = 1.00%), p = 0.042, reps = 3, n = 140] but not CQ alone by 19.2% [65.40% (SD = 7.63%), p = 0.077, reps = 3, n = 91] in comparison to control [80.91% (SD = 4.12%), reps = 3, n = 615] (Fig 6A). There were no significant differences in oocyst prevalence between the unprocessed and reconstituted treatment regimens [IVM+CQ p = 0.246], [CQ p = 0.784], [PQ+CQ p = 0.120] and [IVM+PQ+CQ p = 0.128]. The infection intensity (oocysts number) was significantly reduced in mosquitos fed with unprocessed: IVM+CQ by 90.1% [1.99 (SD = 2.01), p<0.001, reps = 3, n = 64]; PQ+CQ by 90.8% [1.86 (SD = 1.77), p<0.001, reps = 3, n = 161], IVM+PQ+CQ by 89.1% [2.19 (SD = 0.96), p<0.001, reps = 3, n = 99] and CQ alone by 56.5% [8.71 (SD = 8.92), p<0.001, reps = 3, n = 58] compared with the control [20.02 (SD = 4.05), reps = 3, n = 615] (Fig 6B) and reconstituted with plasma from patients that undertook IVM+CQ by 84.1% [3.19 (SD = 1.43), p<0.001, reps = 3, n = 205], PQ+CQ 71.6% [5.69 (SD = 5.14), p<0.001, reps = 3, n = 180], IVM+PQ+CQ by 87.6% [2.50 (SD = 0.64), p<0.001, reps = 3, n = 140] and [20.02 (SD = 4.05), reps = 3, n = 615]. There were no significant differences in oocyst prevalence between the unprocessed and reconstituted treatment regimens [IVM+CQ p = 0.246], [CQ p = 0.784], [PQ+CQ p = 0.120] and [IVM+PQ+CQ p = 0.128].
Fig 6. Plasmatic ivermectin and malaria drugs have a direct effect on Plasmodium vivax infection in Anopheles aquasalis.

An. aquasalis female mosquitos were fed with unprocessed blood samples from P. vivax patients at time 0 (control) or 4 hours after different treatment regimens intake or with erythrocytes from P. vivax patients’ blood samples (before treatment) reconstituted to a hematocrit of 40% with the respective plasma after 4 hours of different treatment regimens intake. (A) The proportion of P. vivax infected mosquitos are presented as mean and standard deviation. Black bars represent the unprocessed blood and the grey bars represent the reconstitute samples. (B) The intensity of infection is presented as the number of oocysts per a single mosquito midgut (dots), black dots represent the unprocessed blood and the grey dots represent the reconstituted blood samples, and the black lines represent the median and standard error blood. Asterisks (*) represent significant difference (P < 0.05) in relation to the control.
mIVM impairs asexual P. vivax maturation
A total of 5 malaria vivax patients were recruited with high parasitemia (138–245 parasites/200 leucocytes), with at least 80% of parasites in the ring stage. We tested five isolates of P. vivax for pIVM drug sensitivity. The pIVM had some activity against two of five P. vivax isolates tested (Table 1). As expected in our site, chloroquine was fully effective as the blood schizonticidal treatment with pCQ IC50 ranging from 1.96 ng/mL to 7.53ng/mL. Interestingly, when mIVM was added to P. vivax culture in different dilutions, a significant reduction in parasite maturation was observed compared to drug free control 1:2 (34.24 ng/ml) [52.31% (SD = 18.06%), p = 0,011, reps = 5]; 1:4 (17.12 ng/ml) [52.34% (SD = 11.49%), p = 0.0002, reps = 5]; 1:8 (8.56 ng/ml) [54.93% (SD = 11.78%), p = 0.0013, reps = 5] and 1:16 (4.28 ng/ml) [51.77% (SD = 9.92%), p = 0.0001, reps = 5] (Fig 7).
Table 1. Ex vivo activity of ivermectin and chloroquine against P. vivax isolates from Manaus- AM, Brazil.
| Isolate | pIVM (ng/ml) | pCQ (ng/ml) | ||||
|---|---|---|---|---|---|---|
| IC50 | IC90 | IC99 | IC50 | IC90 | IC99 | |
| 1 | 14.83 | 18.48 | 23.49 | 7.53 | 464.13 | 41687.71 |
| 2 | NI | NI | NI | 5.60 | 85.92 | 1693.20 |
| 3 | 3.61 | 4.5 | 5.71 | 1.98 | 2.47 | 3.13 |
| 4 | NI | NI | NI | 1.85 | 2.31 | 2.93 |
| 5 | NI | NI | NI | 1.96 | 2.43 | 3.10 |
Expressed as IC50(ng/ml). NI: no inhibition. pIVM: powdered ivermectin, pCQ: powdered chloroquine
Fig 7. Metabolized ivermectin impairs asexual P. vivax maturation.

Five P. vivax samples containing more than 80% of parasites on ring stage were evaluated. Nine different concentrations of pIVM were added to P. vivax culture. Moreover, plasma from healthy volunteers collected before (0 hours) and after 4 hours of IVM intake was added in 4 different serial dilutions (1:2) with complete medium. A drug free condition was used as the control. The number of schizonts in 200 asexual parasites was evaluated for each condition and the maturation in relation to control were determined. Asterisks (*) represents significant differences (P < 0.05) in relation to control by Kruskal-Wallis test.
Discussion
Effective malaria transmission-blocking tools are an integral element in malaria eradication campaigns. MDA of IVM disrupted malaria parasite transmission in West Africa [9, 49] by killing the vector An. gambiae [9, 50] which shifts the population age structure, thereby reducing the sporozoite rate. Additional effects of ivermectin which likely further reduce transmission include inhibiting sporogony in the vector as demonstrated with P. falciparum in An. gambiae [45] and P. vivax in An. dirus, An. minimus [37], and An. darlingi [43]. Previous findings from our group also showed effects of pIVM and mIVM on survivorship, fecundity and even the locomotor activity of An. aquasalis [39, 48]. Therefore, IVM MDA has strong potential to be a novel tool for malaria transmission control. To our knowledge, this is the first study to evaluate mIVM effects on the P. vivax oocyst infection and intensity in Anopheles. This is also the first study to asses IVM effects against P. vivax asexual blood-stage development.
Herein, it was demonstrated that IVM reduces the oocyst infection and intensity of P. vivax. When An. aquasalis were fed with different concentrations of pIVM, a reduction of oocyst infection rate and infection intensity at the 10, 20 and 40 ng/ml concentrations (Fig 2). These ivermectin concentrations were selected based on the human pharmacokinetic curve for IVM which corresponds to approximately the concentration found in human blood between 4 to 60 hours post ingestion of IVM at 200 μg/kg [33, 55]. A recent report demonstrates that An. darlingi had modest sporontocidal pIVM results wherein oocyst prevalence was significantly reduced by 22.6% at the lethal concentration that kills 50% (LC50) (43.2 ng/ml) and 17.1% at the LC25 (27.8 ng/ml) but not significantly by 11.3% at the LC5 (14.8 ng/ml). Furthermore, there were no reductions in oocyst intensity in An. darlingi at any pIVM concentration tested [43]. Other reports also show the effect of pIVM on P. vivax oocyst prevalence and intensity in Asian vectors Anopheles dirus and Anopheles minimus wherein sporontocidal effect was far more impactful [38].
Interestingly, when An. aquasalis was fed P. vivax with mIVM, a reduction on infection rate and intensity was observed with plasma collected at 4 hours, 1 and 5 days after drug intake by the healthy volunteers. A far more potent sporontocidal effect was observed in An. aquasalis ingesting mIVM concentrations from day 1 (5.55 ng/ml) reduced oocyst prevalence by 51.5% compared to pIVM at 5 ng/ml by 12.6%. A similar effect was observed in An. darlingi ingesting mIVM concentrations from day 1 (5.69 ng/ml) reduced oocyst prevalence by 73.9% compared to pIVM at 14.8 ng/ml reduced oocyst prevalence by 11.3% [43]. Here we demonstrate that metabolized ivermectin has a more potent sporontocidal effect compared to ivermectin compound (Fig 4), which suggests that ivermectin metabolites may enhance the sporontocidal effect. Little is known about ivermectin metabolite production in humans, one small trial (n = 4) indicated that mean peak plasma concentration of metabolites was 2.5 fold greater than that of the parent compound and the effective half-lives of the metabolites were approximately 2.9 days while the parent compound half-life was 11.8 hours [59]. Future studies should be designed to elucidate ivermectin metabolite production in orally-treated volunteers and their impact on mosquito survivorship and Plasmodium sporogony. Interestingly, oocyst prevalence and intensity was more intensely impacted with mIVM plasma from 1 day in An. darlingi compared to An. aquasalis (Fig 3) but no significant reduction occurred when plasma from day 5 was fed to An. darlingi which suggests a shorter duration of but stronger sporontocidal effect compared to An. aquasalis.
Mosquito mortality effect was observed at day 7 post-blood feed with mIVM plasma from 4 hours and 1 day in the two-species studied. This IVM effect on mosquito survival is similar to our previous findings in non-infected An. aquasalis which also used metabolized ivermectin showing the higher impact on the survival and reproductive fitness [39] and with other studies in different anopheline species infected and uninfected with Plasmodium [9, 50, 60, 61]. In a single dose of 200 mcg/kg showed an increase in mosquito mortality in An. aquasalis when fed on mIVM at 1 day (5.55 ng/ml) to 5 days (1.58 ng/ml) the drug intake, but there was no longer effect when fed plasma collected from days 10 or 14.
The reduction in An. aquasalis and An. darlingi oocyst infection and intensity when mosquitoes were blood fed mIVM 4 days after infection expands the window that a blood meal containing IVM has an effect on P. vivax mosquito infection. Moreover, IVM was able to impair parasite development even when it was given to the mosquitoes after the midgut epithelium invasion by parasite ookinete suggesting that IVM has direct effects on already established and developing oocysts. Interestingly, these results were different from Kobylinski et al. [45], wherein no sporontocidal effect was observed on early-stage oocyst development when pIVM was fed 3 days after P. falciparum infection in An. gambiae.
This study also evaluates the in vivo exposure of P. vivax to IVM, CQ, and PQ in the human and its subsequent development in An. aquasalis. Mosquito oocyst infection and intensity were significantly reduced when mosquitoes were fed blood from patients treated with IVM+CQ, PQ+CQ or IVM+PQ+CQ but not CQ alone (Fig 6). Importantly, this is the first study to show that primaquine has a sporontocidal effect on P. vivax infection in the mosquito. However, the reduction on infection rate and intensity were not augmented on IVM +PQ+ CQ treated blood in relation to IVM+CQ only, suggesting that PQ does not have an additive or synergistic effect beyond IVM.
The mosquitoes were exposed in parallel to P. vivax with unprocessed and reconstituted blood samples. Unprocessed blood samples allowed for in vivo exposure of P. vivax to the drugs in the human compared to reconstituted blood samples which investigated the impact of metabolized drugs on P. vivax. The reduction in oocyst infection and intensity found in IVM+CQ and IVM+PQ+CQ treatment groups was similar in mosquitoes fed with the unprocessed and reconstituted blood (Fig 6). These data indicate that the ivermectin transmission blocking effect occurs in the mosquito midgut and not in human blood. Similar results were observed in the PQ+CQ and IVM+PQ+CQ groups, suggesting that primaquine effect on mosquito infection also occurs in the midgut. Since these assays were performed at only one-time point after drug intake (4 hours), we could not discard a possible delayed in vivo effect of primaquine or ivermectin on P. vivax asexual stages and gametocytes in the patient.
It is important to note that all patients who received the different regimens treatment were also supplied with CQ at the same time, following the guidelines of the Brazilian Health Ministry, which recommend all the patients ethically have to receive the CQ treatment at the same time that they are diagnosed. As expected, the mosquitoes fed a blood meal containing only CQ did not show a decrease in the oocyst infection prevalence of An. aquasalis with P. vivax, which is in accordance with other studies, where CQ did not affect the oocyst prevalence of Plasmodium berghei in An. gambiae [62]. However, this is the first report to demonstrate that oocyst intensity was reduced in mosquitoes fed a blood meal containing CQ. A reduction in oocyst intensity by CQ could be due to some direct action on P. vivax, or its immunosuppressive potential [63] may interfere with successful parasite midgut invasion leading to fewer oocysts.
Our results showed the highest mortality rate reduction of infection rate and intensity of An. aquasalis and An. darlingi, two important vectors of South American on plasma 4 hours and considering that in the human pharmacokinetic curve of the IVM the mean peak plasma concentrations is (46.6 ± 21.9ng/ml) at approximately 4 hours after dosing, with a IVM half-life from about 12 to 56 hours [33]. Similar peaks have been found in Primaquine and Chloroquine [64]. We can suggest the use of the IVM as a potent way to administration in combination with the other antimalaria drugs and mainly during the first hours after being detected the infection in the patient, which would have a higher impact in the Malaria elimination and eradication programs, specially, in endemics areas like Amazonas Region, which, have high incidence of Malaria cases by P. vivax.
This is the first study to assess the effect of ivermectin against asexual P. vivax. Two previous studies demonstrated an inhibition of pIVM on P. falciparum asexual stage development but with IC50s in the 1–10 μg/ml range [51, 65]. No effect of pIVM (3.9–1000 ng/ml) was observed in the current study against asexual blood-stage P. vivax, but this may have been due to using too low concentration (Fig 7). On the other hand, when asexual P. vivax was incubated with 4 different dilutions (4.28, 8.56, 17.12 and 34.24 ng/ml) of plasma obtained from healthy volunteers 4 hours after IVM administration, there was a significant decrease in P. vivax maturation in relation to the drug free control and incubations with plasma from healthy volunteers collected before IVM administration. It is important to highlight in the present study that when the asexual P. vivax was incubated with the pIVM (3.9 – 1000ng/ml) the development was not affected, however, a considerable reduction in blood-stage development was observed when the asexual stages were incubated with mIVM (4.28–34.24 ng/ml). This discrepancy might be a result of IVM metabolites conferring the parasite maturation inhibition effect. It is important to note that mIVM concentrations that showed blood-stage inhibition were achieved following oral administration with a standard dose of ivermectin (200 μg/kg). Unfortunately, data collected in this study could not be used to elucidate the ivermectin mechanism of action against asexual P. vivax. Further studies are warranted to evaluate the safety and efficacy of ivermectin as an adjunct during P. vivax antimalarial therapy.
We also have assayed the P. vivax sensitivity to pCQ on the same isolates used to examine the asexual maturation inhibition with IVM. These results showed pCQ IC50 values ranging from 1.96 ng/mL to 7.53ng/mL, similar to other reports [66–69], which demonstrate the chloroquine effect on P. vivax. Our findings also confirm the effect of chloroquine in terms of its pharmacodynamics against P. vivax.
In conclusion, our study shows for the first time the effect of mIVM on the oocyst infection and intensity of P. vivax in the South American malaria vectors An. aquasalis and An. darlingi. In both vectors it appears that mIVM has a stronger sporontocidal effect compared to pIVM, this suggests that ivermectin metabolites have sporontocidal effect. We report for the first time, the effect of IVM on ex vivo cultures of P. vivax and demonstrate that mIVM can inhibit P. vivax development. Moreover, it provides evidence that IVM may affect several parameters of Ross-MacDonald model [70], including parasite life cycle stages, placing it as a strong candidate for malaria transmission reduction.
Supporting information
(DOCX)
(XLSX)
Acknowledgments
We thank all vivax patients, healthy volunteers, and the technical staff from Malaria and Entomology laboratory groups of Fundação de Medicina Tropical Doutor Heitor Viera Dourado and the technical staff for Entomology and Parasitology NAMRU-6, Iquitos, Peru. The material has been reviewed by NAMRU6 and the Walter Reed Army Institute of Research. The study protocol was approved by the Naval Medical Research Center Institutional Review Board (Protocol # NMRCD.2008.0004) in compliance with all applicable Federal regulations governing the protection of human subjects. There is no objection to its presentation and/or publication. The opinions or assertions contained herein are the private views of the authors and are not to be construed as official, or as reflecting true views of the Department of the Navy, Department of the Army, or the Department of Defense. The investigators have adhered to the policies for protection of human subjects as prescribed in AR 70–25.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by National Counsel of Technological and Scientific development (CNPq), Coordination for the Improvement of Higher Education Personnel (CAPES) and Research Support Foundation of Amazonas (FAPEAM) through PPSUS and PAPAC projects supported this study. Bill & Melinda Gates Foundation has also funded this study through TransEpi Project. PFPP and MVGL are level 1 fellows from CNPq. VSS and YTP have fellowships from CAPES. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.WHO. Implications of insecticide resistance for malaria vector control. World Health Organization; http://wwwwhoint/malaria/news/2016/iir-malaria-vectorcontrol-evaluation-nov2016pdf. 2016. [Google Scholar]
- 2.Oliveira-Ferreira J, Lacerda MV, Brasil P, Ladislau JL, Tauil PL, Daniel-Ribeiro CT. Malaria in Brazil: an overview. Malaria journal. 2010;9:115 doi: 10.1186/1475-2875-9-115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Recht J, Siqueira AM, Monteiro WM, Herrera SM, Herrera S, Lacerda MVG. Malaria in Brazil, Colombia, Peru and Venezuela: current challenges in malaria control and elimination. Malaria journal. 2017;16(1):273 doi: 10.1186/s12936-017-1925-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rosas-Aguirre A, Gamboa D, Manrique P, Conn JE, Moreno M, Lescano AG, et al. Epidemiology of Plasmodium vivax Malaria in Peru. Am J Trop Med Hyg. 2016;95(6 Suppl):133–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Abdallah TM, Ali AA, Bakri M, Gasim GI, Musa IR, Adam I. Efficacy of artemether-lumefantrine as a treatment for uncomplicated Plasmodium vivax malaria in eastern Sudan. Malaria journal. 2012;11:404 doi: 10.1186/1475-2875-11-404 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gogtay N, Kannan S, Thatte UM, Olliaro PL, Sinclair D. Artemisinin-based combination therapy for treating uncomplicated Plasmodium vivax malaria. The Cochrane database of systematic reviews. 2013(10):CD008492 doi: 10.1002/14651858.CD008492.pub3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Naing C, Racloz V, Whittaker MA, Aung K, Reid SA, Mak JW, et al. Efficacy and safety of dihydroartemisinin-piperaquine for treatment of Plasmodium vivax malaria in endemic countries: meta-analysis of randomized controlled studies. PLoS One. 2013;8(12):e78819 doi: 10.1371/journal.pone.0078819 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Njoroge MM, Tirados I, Lindsay SW, Vale GA, Torr SJ, Fillinger U. Exploring the potential of using cattle for malaria vector surveillance and control: a pilot study in western Kenya. Parasit Vectors. 2017;10(1):18 doi: 10.1186/s13071-016-1957-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Alout H, Krajacich BJ, Meyers JI, Grubaugh ND, Brackney DE, Kobylinski KC, et al. Evaluation of ivermectin mass drug administration for malaria transmission control across different West African environments. Malaria journal. 2014;13:417 doi: 10.1186/1475-2875-13-417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pluess B, Tanser FC, Lengeler C, Sharp BL. Indoor residual spraying for preventing malaria. The Cochrane database of systematic reviews. 2010(4):CD006657 doi: 10.1002/14651858.CD006657.pub2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Coleman M, Hemingway J, Gleave KA, Wiebe A, Gething PW, Moyes CL. Developing global maps of insecticide resistance risk to improve vector control. Malaria journal. 2017;16(1):86 doi: 10.1186/s12936-017-1733-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Molina D, Figueroa LE. [Metabolic resistance to organophosphate insecticides in Anopheles aquasalis Curry 1932, Libertador municipality, Sucre State, Venezuela]. Biomedica. 2009;29(4):604–15. [PubMed] [Google Scholar]
- 13.Quinones ML, Norris DE, Conn JE, Moreno M, Burkot TR, Bugoro H, et al. Insecticide Resistance in Areas Under Investigation by the International Centers of Excellence for Malaria Research: A Challenge for Malaria Control and Elimination. Am J Trop Med Hyg. 2015;93(3 Suppl):69–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Santacoloma L, Tibaduiza T, Gutierrrez M, Brochero H. [Susceptibility to insecticides of Anopheles darlingi Root 1840, in two locations of the departments of Santander and Caqueta, Colombia]. Biomedica. 2012;32 Suppl 1:22–8. [DOI] [PubMed] [Google Scholar]
- 15.Zamora Perea E, Balta Leon R, Palomino Salcedo M, Brogdon WG, Devine GJ. Adaptation and evaluation of the bottle assay for monitoring insecticide resistance in disease vector mosquitoes in the Peruvian Amazon. Malaria journal. 2009;8:208 doi: 10.1186/1475-2875-8-208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bahia AC, Kubota MS, Tempone AJ, Pinheiro WD, Tadei WP, Secundino NF, et al. Anopheles aquasalis Infected by Plasmodium vivax displays unique gene expression profiles when compared to other malaria vectors and plasmodia. PLoS One. 2010;5(3):e9795 doi: 10.1371/journal.pone.0009795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.da Silva AN, Santos CC, Lacerda RN, Machado RL, Povoa MM. Susceptibility of Anopheles aquasalis and an. darlingi to Plasmodium vivax VK210 and VK247. Mem Inst Oswaldo Cruz. 2006;101(5):547–50. [DOI] [PubMed] [Google Scholar]
- 18.Rios-Velasquez CM, Martins-Campos KM, Simoes RC, Izzo T, dos Santos EV, Pessoa FA, et al. Experimental Plasmodium vivax infection of key Anopheles species from the Brazilian Amazon. Malaria journal. 2013;12:460 doi: 10.1186/1475-2875-12-460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Orfano AS, Nacif-Pimenta R, Duarte AP, Villegas LM, Rodrigues NB, Pinto LC, et al. Species-specific escape of Plasmodium sporozoites from oocysts of avian, rodent, and human malarial parasites. Malaria journal. 2016;15(1):394 doi: 10.1186/s12936-016-1451-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pimenta PF, Orfano AS, Bahia AC, Duarte AP, Ríos-Velásquez CM, Melo FF, et al. An overview of malaria transmission from the perspective of Amazon Anopheles vectors. Mem Inst Oswaldo Cruz. 2015;110(1):23–47. doi: 10.1590/0074-02760140266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hiwat H, Issaly J, Gaborit P, Somai A, Samjhawan A, Sardjoe P, et al. Behavioral heterogeneity of Anopheles darlingi (Diptera: Culicidae) and malaria transmission dynamics along the Maroni River, Suriname, French Guiana. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2010;104(3):207–13. doi: 10.1016/j.trstmh.2009.07.007 [DOI] [PubMed] [Google Scholar]
- 22.Sinka ME, Bangs MJ, Manguin S, Rubio-Palis Y, Chareonviriyaphap T, Coetzee M, et al. A global map of dominant malaria vectors. Parasit Vectors. 2012;5:69 doi: 10.1186/1756-3305-5-69 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Carter R. Transmission blocking malaria vaccines. Vaccine. 2001;19(17–19):2309–14. [DOI] [PubMed] [Google Scholar]
- 24.Carter R, Chen DH. Malaria transmission blocked by immunisation with gametes of the malaria parasite. Nature. 1976;263(5572):57–60. [DOI] [PubMed] [Google Scholar]
- 25.Carter R, Mendis KN, Miller LH, Molineaux L, Saul A. Malaria transmission-blocking vaccines—how can their development be supported? Nat Med. 2000;6(3):241–4. doi: 10.1038/73062 [DOI] [PubMed] [Google Scholar]
- 26.Saul A. Mosquito stage, transmission blocking vaccines for malaria. Curr Opin Infect Dis. 2007;20(5):476–81. doi: 10.1097/QCO.0b013e3282a95e12 [DOI] [PubMed] [Google Scholar]
- 27.Saul A. Efficacy model for mosquito stage transmission blocking vaccines for malaria. Parasitology. 2008;135(13):1497–506. doi: 10.1017/S0031182008000280 [DOI] [PubMed] [Google Scholar]
- 28.Ponsa N, Sattabongkot J, Kittayapong P, Eikarat N, Coleman RE. Transmission-blocking activity of tafenoquine (WR-238605) and artelinic acid against naturally circulating strains of Plasmodium vivax in Thailand. Am J Trop Med Hyg. 2003;69(5):542–7. [PubMed] [Google Scholar]
- 29.Wells TN, Alonso PL, Gutteridge WE. New medicines to improve control and contribute to the eradication of malaria. Nat Rev Drug Discov. 2009;8(11):879–91. doi: 10.1038/nrd2972 [DOI] [PubMed] [Google Scholar]
- 30.Boissière A, Tchioffo MT, Bachar D, Abate L, Marie A, Nsango SE, et al. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012;8(5):e1002742 doi: 10.1371/journal.ppat.1002742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cirimotich CM, Dong Y, Clayton AM, Sandiford SL, Souza-Neto JA, Mulenga M, et al. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science. 2011;332(6031):855–8. doi: 10.1126/science.1201618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fang W, Vega-Rodríguez J, Ghosh AK, Jacobs-Lorena M, Kang A, St Leger RJ. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science. 2011;331(6020):1074–7. doi: 10.1126/science.1199115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Guzzo CA, Furtek CI, Porras AG, Chen C, Tipping R, Clineschmidt CM, et al. Safety, tolerability, and pharmacokinetics of escalating high doses of ivermectin in healthy adult subjects. J Clin Pharmacol. 2002;42(10):1122–33. [DOI] [PubMed] [Google Scholar]
- 34.Hotez PJ. Control of onchocerciasis—the next generation. Lancet. 2007;369(9578):1979–80. doi: 10.1016/S0140-6736(07)60923-4 [DOI] [PubMed] [Google Scholar]
- 35.Omura S. Ivermectin: 25 years and still going strong. Int J Antimicrob Agents. 2008;31(2):91–8. doi: 10.1016/j.ijantimicag.2007.08.023 [DOI] [PubMed] [Google Scholar]
- 36.Ottesen EA, Hooper PJ, Bradley M, Biswas G. The global programme to eliminate lymphatic filariasis: health impact after 8 years. PLoS Negl Trop Dis. 2008;2(10):e317 doi: 10.1371/journal.pntd.0000317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Shmidt E, Levitt J. Dermatologic infestations. Int J Dermatol. 2012;51(2):131–41. doi: 10.1111/j.1365-4632.2011.05191.x [DOI] [PubMed] [Google Scholar]
- 38.Kobylinski KC, Ubalee R, Ponlawat A, Nitatsukprasert C, Phasomkulsolsil S, Wattanakul T, et al. Ivermectin susceptibility and sporontocidal effect in Greater Mekong Subregion Anopheles. Malaria journal. 2017;16(1):280 doi: 10.1186/s12936-017-1923-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sampaio VS, Beltrán TP, Kobylinski KC, Melo GC, Lima JB, Silva SG, et al. Filling gaps on ivermectin knowledge: effects on the survival and reproduction of Anopheles aquasalis, a Latin American malaria vector. Malaria journal. 2016;15(1):491 doi: 10.1186/s12936-016-1540-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chaccour CJ, Kobylinski KC, Bassat Q, Bousema T, Drakeley C, Alonso P, et al. Ivermectin to reduce malaria transmission: a research agenda for a promising new tool for elimination. Malaria journal. 2013;12:153 doi: 10.1186/1475-2875-12-153 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Foy BD, Kobylinski KC, da Silva IM, Rasgon JL, Sylla M. Endectocides for malaria control. Trends in parasitology. 2011;27(10):423–8. doi: 10.1016/j.pt.2011.05.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Meyers JI, Gray M, Foy BD. Mosquitocidal properties of IgG targeting the glutamate-gated chloride channel in three mosquito disease vectors (Diptera: Culicidae). J Exp Biol. 2015;218(Pt 10):1487–95. doi: 10.1242/jeb.118596 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kobylinski KC, Escobedo-Vargas KS, Lopez-Sifuentes VM, Durand S, Smith ES, Baldeviano GC, et al. Ivermectin susceptibility, sporontocidal effect, and inhibition of time to re-feed in the Amazonian malaria vector Anopheles darlingi. Malaria journal. 2017;16(1):474 doi: 10.1186/s12936-017-2125-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kobylinski KC, Alout H, Foy BD, Clements A, Adisakwattana P, Swierczewski BE, et al. Rationale for the coadministration of albendazole and ivermectin to humans for malaria parasite transmission control. Am J Trop Med Hyg. 2014;91(4):655–62. doi: 10.4269/ajtmh.14-0187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kobylinski KC, Foy BD, Richardson JH. Ivermectin inhibits the sporogony of Plasmodium falciparum in Anopheles gambiae. Malaria journal. 2012;11:381 doi: 10.1186/1475-2875-11-381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fritz ML, Siegert PY, Walker ED, Bayoh MN, Vulule JR, Miller JR. Toxicity of bloodmeals from ivermectin-treated cattle to Anopheles gambiae s.l. Annals of tropical medicine and parasitology. 2009;103(6):539–47. doi: 10.1179/000349809X12459740922138 [DOI] [PubMed] [Google Scholar]
- 47.Fritz ML, Walker ED, Miller JR. Lethal and sublethal effects of avermectin/milbemycin parasiticides on the African malaria vector, Anopheles arabiensis. J Med Entomol. 2012;49(2):326–31. [DOI] [PubMed] [Google Scholar]
- 48.Sampaio VS, Rivas G, Kobylinski K, Pinilla YT, Pimenta PFP, Lima JBP, et al. What does not kill it makes it weaker: effects of sub-lethal concentrations of ivermectin on the locomotor activity of Anopheles aquasalis. Parasit Vectors. 2017;10(1):623 doi: 10.1186/s13071-017-2563-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kobylinski KC, Sylla M, Chapman PL, Sarr MD, Foy BD. Ivermectin mass drug administration to humans disrupts malaria parasite transmission in Senegalese villages. Am J Trop Med Hyg. 2011;85(1):3–5. doi: 10.4269/ajtmh.2011.11-0160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sylla M, Kobylinski KC, Gray M, Chapman PL, Sarr MD, Rasgon JL, et al. Mass drug administration of ivermectin in south-eastern Senegal reduces the survivorship of wild-caught, blood fed malaria vectors. Malaria journal. 2010;9:365 doi: 10.1186/1475-2875-9-365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Panchal M, Rawat K, Kumar G, Kibria KM, Singh S, Kalamuddin M, et al. Plasmodium falciparum signal recognition particle components and anti-parasitic effect of ivermectin in blocking nucleo-cytoplasmic shuttling of SRP. Cell Death Dis. 2014;5:e994 doi: 10.1038/cddis.2013.521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mendes AM, Albuquerque IS, Machado M, Pissarra J, Meireles P, Prudencio M. Inhibition of Plasmodium Liver Infection by Ivermectin. Antimicrob Agents Chemother. 2017;61(2). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Saúde Md. Guia prático de tratamento da malária no Brasil. Secretaria de Vigilância em Saúde, Departamento de Vigilância Epidemiológica. Série A Normas e Manuais Técnicos. 2010.
- 54.Villarreal-Trevino C, Vasquez GM, Lopez-Sifuentes VM, Escobedo-Vargas K, Huayanay-Repetto A, Linton YM, et al. Establishment of a free-mating, long-standing and highly productive laboratory colony of Anopheles darlingi from the Peruvian Amazon. Malaria journal. 2015;14:227 doi: 10.1186/s12936-015-0733-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Baraka OZ, Mahmoud BM, Marschke CK, Geary TG, Homeida MM, Williams JF. Ivermectin distribution in the plasma and tissues of patients infected with Onchocerca volvulus. Eur J Clin Pharmacol. 1996;50(5):407–10. [DOI] [PubMed] [Google Scholar]
- 56.Administration FaD. Guidance for industry: bioanalytical method validation. U.S. Department of Health and Human Services, Rockville, Maryland: 2001. [Google Scholar]
- 57.Russell BM, Udomsangpetch R, Rieckmann KH, Kotecka BM, Coleman RE, Sattabongkot J. Simple in vitro assay for determining the sensitivity of Plasmodium vivax isolates from fresh human blood to antimalarials in areas where P. vivax is endemic. Antimicrob Agents Chemother. 2003;47(1):170–3. doi: 10.1128/AAC.47.1.170-173.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Vera O, Brelas de Brito P, Albrecht L, Martins-Campos KM, Pimenta PF, Monteiro WM, et al. Purification Methodology for Viable and Infective Plasmodium vivax Gametocytes That Is Compatible with Transmission-Blocking Assays. Antimicrob Agents Chemother. 2015;59(10):6638–41. doi: 10.1128/AAC.01136-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Laboratories MR. Stromectol new drug application. In: FDA Center for drug evaluation and research. 1996. Available from: http://wwwaccessdatafdagov/drugsatfda_docs/label/2009/050742s026lblpdf
- 60.Butters MP, Kobylinski KC, Deus KM, da Silva IM, Gray M, Sylla M, et al. Comparative evaluation of systemic drugs for their effects against Anopheles gambiae. Acta Trop. 2012;121(1):34–43. doi: 10.1016/j.actatropica.2011.10.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kobylinski KC, Deus KM, Butters MP, Hongyu T, Gray M, da Silva IM, et al. The effect of oral anthelmintics on the survivorship and re-feeding frequency of anthropophilic mosquito disease vectors. Acta Trop. 2010;116(2):119–26. doi: 10.1016/j.actatropica.2010.06.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Abrantes P, Dimopoulos G, Grosso AR, do Rosario VE, Silveira H. Chloroquine mediated modulation of Anopheles gambiae gene expression. PLoS One. 2008;3(7):e2587 doi: 10.1371/journal.pone.0002587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Thome R, Lopes SC, Costa FT, Verinaud L. Chloroquine: modes of action of an undervalued drug. Immunol Lett. 2013;153(1–2):50–7. doi: 10.1016/j.imlet.2013.07.004 [DOI] [PubMed] [Google Scholar]
- 64.Pukrittayakamee S, Tarning J, Jittamala P, Charunwatthana P, Lawpoolsri S, Lee SJ, et al. Pharmacokinetic interactions between primaquine and chloroquine. Antimicrob Agents Chemother. 2014;58(6):3354–9. doi: 10.1128/AAC.02794-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Nasveld P, Russell B, Kotecka B, Rieckmann K. Lack of in vitro effect of ivermectin on Plasmodium falciparum. Southeast Asian J Trop Med Public Health. 2003;34(3):552–3. [PubMed] [Google Scholar]
- 66.Wirjanata G, Handayuni I, Prayoga P, Apriyanti D, Chalfein F, Sebayang BF, et al. Quantification of Plasmodium ex vivo drug susceptibility by flow cytometry. Malaria journal. 2015;14:417 doi: 10.1186/s12936-015-0940-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Rungsihirunrat K, Muhamad P, Chaijaroenkul W, Kuesap J, Na-Bangchang K. Plasmodium vivax drug resistance genes; Pvmdr1 and Pvcrt-o polymorphisms in relation to chloroquine sensitivity from a malaria endemic area of Thailand. Korean J Parasitol. 2015;53(1):43–9. doi: 10.3347/kjp.2015.53.1.43 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Fernandez D, Segura C, Arboleda M, Garavito G, Blair S, Pabon A. In vitro susceptibility of Plasmodium vivax to antimalarials in Colombia. Antimicrob Agents Chemother. 2014;58(11):6354–9. doi: 10.1128/AAC.03191-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Aguiar AC, Pereira DB, Amaral NS, De Marco L, Krettli AU. Plasmodium vivax and Plasmodium falciparum ex vivo susceptibility to anti-malarials and gene characterization in Rondonia, West Amazon, Brazil. Malaria journal. 2014;13:73 doi: 10.1186/1475-2875-13-73 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Smith DL, Battle KE, Hay SI, Barker CM, Scott TW, McKenzie FE. Ross, macdonald, and a theory for the dynamics and control of mosquito-transmitted pathogens. PLoS Pathog. 2012;8(4):e1002588 doi: 10.1371/journal.ppat.1002588 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.