
Fig. 1.
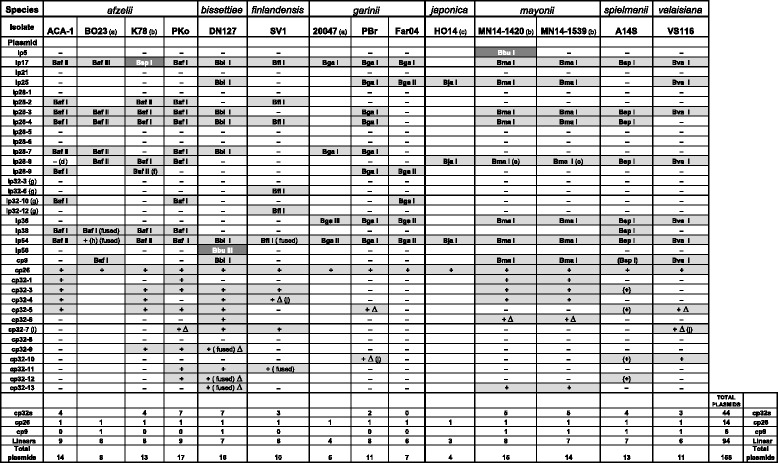
Sequenced plasmids in NBu-Borreliella species. The eleven completely sequenced genomes and three partly sequence genomes are shown as columns where shaded cells indicate the plasmid’s presence, “fused” indicates that the indicated plasmids contain apparently intact PFam32 genes of more than one type (see text); and “∆”, indicates the presence of a substantial deletion relative to other cp32s. Hyphens (−) denote plasmids that are known not to be present, and blank cells indicate that it is not known if that plasmid type is present. For the linear plasmids and cp9, Roman numerals indicate plasmid organizational subtype (see text and [20] for subtype definitions); here subtypes are named by the first letter of the genus name and first two letters of the species name (e.g., B. afzelii subtype I is “Baf I”). Similar subtype numerals in different plasmid types (columns in table) does not imply any relationship, and subtype organizations are always different in the different species except in the three cases that are indicated by dark gray cells (see text). The cp26 plasmids do not exhibit organizational variation and cp32s are too variable, so no subtypes are defined for these plasmids; a “+” indicates that a plasmid of that PFam32 type is present, and parentheses (...) around the subtype name denote the cp9 and four cp32 isolate A14S plasmids whose sequences were not closed. Because the sequences of these plasmids in A14S were not closed, it is possible that some cp32s are fused, so the number of different plasmid DNA molecules is not known precisely in this case. (a) The indicated plasmid sequences from isolates B. afzelii BO23 and B. garinii 20047 have been deposited in GenBank as closed sequences (Additional file 1: Table S1); it is not known what other plasmids these isolates may carry (S. Bontemps-Gallo, Pers. Com.); (b) these plasmids represent the full complement of plasmids of B. afzelii K78 and the two B. mayonii isolates; accession numbers are listed in [15, 21], respectively; (c) the plasmid sequences from isolate B. japonica HO14 (ATCC51557) are “minimal draft” quality sequences; they are included here because they are the right size to be full plasmid sequences but the details of their sequences should be interpreted with caution. (d) plasmid is very likely present in the original ACA-1 isolate, see text; (e) called lp28–10 in reference [15], see text; (f) called lp28–1 in reference [21], see text; (g) the lp32 plasmids very likely have the same compatibility type as cp32 of the same number, see text; (h) the BO23 lp54 sequence does not include the PFam54 gene array that distinguishes the subtypes of this plasmid; (i) previously named cp32–2 and cp32–7 have the same PFam32 protein type; we use cp32–7 to represent this group; and (j) several kbp of typically linear plasmid sequence replaces at least part of the deleted cp32 DNA