
Fig. 2.
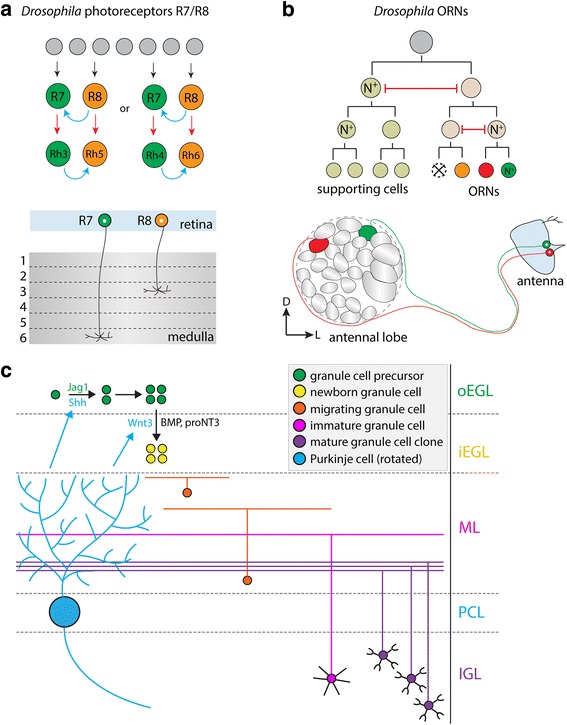
Extrinsic and stochastic regulation of neural wiring. a In the Drosophila retina, photoreceptors R7 and R8 (and R1-R6; not shown) are produced from a pool of progenitors. Cell-cell interactions (blue arrows) and stochastic mechanisms (red arrows) play critical roles in cell fate specification. Mature R7 and R8 cells project their axons to layers 6 and 3, respectively, of the medulla. Rhodopsin: Rh. b In the Drosophila olfactory receptor neuron (ORN) lineage, one progenitor cell within each sensillum undergoes several rounds of asymmetric division, giving rise to four non-neuronal supporting cells and between one and four ORNs depending on other events, such as cell death and glial fate adoption. Binary Notch signaling activation is iteratively employed, and lateral inhibition (red bars) is required for cell fate determination. Notch-ON (N+) and Notch-OFF ORNs send their axons to different glomeruli in the antennal lobe. D: dorsal; L: lateral. c In the developing mouse cerebellum, granule cell precursors (GCPs) in the outer external germinal layer (oEGL) undergo steady proliferation in a process promoted by Purkinje cell-derived Shh and GCP-derived Jag1. GCPs in the inner external germinal layer (iEGL) undergo a rapid burst of cell division before terminal differentiation, a process promoted by Wnt3 (expressed by Purkinje cells), BMP and proNT3. Migrating granule cells (GCs) then extend their parallel fiber axons into the molecular layer (ML), where they contact the dendritic arbors of developing Purkinje cells (rotated 90 degrees). Clones of mature GCs, which are born around the same time, project their parallel fiber axons to restricted depths of the ML. Parallel fibers of early-born GCs thus occupy the deepest depths of the ML while those of late-born GCs occupy the most superficial depths of the ML. PCL: Purkinje cell layer; IGL: internal granule layer