

Abstract
18O discrimination in CO2 stems from the oxygen exchange between 18O-enriched water and CO2 in the chloroplast, a process catalyzed by carbonic anhydrase (CA). A proportion of this 18O-labeled CO2 escapes back to the atmosphere, resulting in an effective discrimination against C18OO during photosynthesis (Δ18O). By constraining the δ18O of chloroplast water (δe) by analysis of transpired water and the extent of CO2-H2O isotopic equilibrium (θeq) by measurements of CA activity (θeq = 0.75–1.0 for tobacco, soybean, and oak), we could apply measured Δ18O in a leaf cuvette attached to a mass spectrometer to derive the CO2 concentration at the physical limit of CA activity, i.e. the chloroplast surface (ccs). From the CO2 drawdown sequence between stomatal cavities from gas exchange (ci), from Δ18O (ccs), and at Rubisco sites from Δ13C (cc), the internal CO2 conductance (gi) was partitioned into cell wall (gw) and chloroplast (gch) components. The results indicated that gch is variable (0.42–1.13 mol m−2 s−1) and proportional to CA activity. We suggest that the influence of CA activity on the CO2 assimilation rate should be important mainly in plants with low internal conductances.
Natural variation in 18O content (δ18O) of CO2 is a useful tracer for photosynthetic activity. This is due to a sequence of events: first, δ18O of chloroplast water is high due to evaporative effects (Gonfiantini et al., 1965); second, in the chloroplasts, exchange of oxygen between CO2 and H2O is catalyzed by carbonic anhydrase (CA); and third, a large fraction of this 18O-labeled CO2 diffuses from the chloroplast back to the atmosphere. On a leaf scale, this “retroflux” of 18O-enriched CO2 from the leaf back to the atmosphere is observed as an enrichment in the C18OO in air passing over the leaf or as discrimination against C18OO by the leaf (Δ18O) (Farquhar and Lloyd, 1993). Notably, Δ18O is also observed on a global scale as latitudinal and seasonal changes in the δ18O of atmospheric CO2. The quantitative use of such large-scale signals, however, still critically depends on better understanding of the basic processes influencing Δ18O (Francey and Tans, 1987; Farquhar et al., 1993; Ciais et al., 1997).
To interpret Δ18O measured during leaf-atmosphere CO2 exchange, an estimate of CO2 concentration at the site of CO2-H2O is required (Farquhar and Lloyd, 1993). The chloroplast CO2 concentration (cc) may be derived from comparing measured and modeled discrimination against 13CO2 (Δ13C) (Farquhar et al., 1982; Evans et al., 1986; von Caemmerer and Evans, 1991). Since both the photosynthetic enzyme Rubisco (responsible for 13C discrimination) and CA (responsible for Δ18O) are similarly distributed within the chloroplast stroma (Anderson et al., 1996), the 13C-derived value of cc was also applied to Δ18O (Farquhar et al., 1993; Flanagan et al., 1994). However, it was suggested (Yakir, 1998) that the CO2 concentration pertaining to Δ18O may be associated with the chloroplast surface, i.e. the limit of CA activity, and not the mean CO2 concentration at sites of CO2 fixation by Rubisco. This is because CA acts to cancel out any gradients in 18O of CO2 within its domain. We now suggest that with adequate estimates of chloroplast water δ18O and of the extent of CO2-H2O isotopic equilibrium in the chloroplast (i.e. CA activity), it should be possible to use Δ18O to accurately estimate the effective CO2 concentration at the sites of CO2-H2O equilibrium. This approach is somewhat similar to that using observed and predicted Δ13C to compare ci and cc (von Caemmerer and Evans, 1991).
Using 13C-derived estimates of cc, the internal leaf conductance to CO2 (gi) and its influence on leaf photosynthesis have been well characterized (von Caemmerer and Evans, 1991; Lloyd et al., 1992; Loreto et al., 1992; Syvertsen et al., 1995). However, evaluating the relative importance of the major components of gi, the wall conductance (gw) and the chloroplast conductance (gch) has been restricted (Cowan, 1986; Evans et al., 1994). The association of Δ18O with CO2 concentration at the chloroplast surface should enable this partitioning. Information on CA activity directly from assays or through Δ18O measurements should also provide insight into the role of CA in facilitating diffusion within the chloroplast (Cowan, 1986; Makino et al., 1992; Price et al., 1994; Williams et al., 1996).
By comparing the CO2 concentration and isotopic composition of air entering and leaving a leaf chamber, dis-crimination against C18OO (Δ18O) may be measured “on-line” in a method equivalent to Δ13C (Evans et al., 1986):
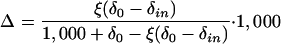 |
1 |
where ξ = cin/(cin-co), cin, co, and δin, δo referring to the CO2 concentration (corrected to the same humidity) and isotopic composition of air entering and leaving the cuvette, respectively. Δ18O can also be predicted (Farquhar and Lloyd, 1993) as
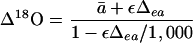 |
2 |
where Δea = 1,000·[(δe/1,000 +1)/(δa/1,000 + 1) −1]; ε = cc/(ca − cc); δa and δe represent the δ18O of CO2 in the overlying air and in full isotopic equilibrium with water in the chloroplast, respectively, and ca and cc the respective CO2 concentrations (see Fig. 1); ā is the weighted-mean diffusional fractionation through the boundary layer, 5.8‰, stomata, 8.8‰, and aqueous leaf media, 0.8‰, (Farquhar and Lloyd, 1993). Despite general agreement between modeled and measured Δ18O (Farquhar et al., 1993; Flanagan et al., 1994; Williams et al., 1996), large quantitative discrepancies often occur (Yakir et al., 1994; Williams et al., 1996; Harwood et al., 1998; Wang et al., 1998). There are three main assumptions in Equation 2: (a) chloroplast water (and hence CO2 in equilibrium with this water) is assumed to be isotopically similar to water at the evaporating sites (δe); (b) CO2 and H2O in the chloroplast reach full isotopic equilibrium; and (c) cc correctly represents the CO2 concentration at the site of oxygen exchange.
Figure 1.

Diagram showing the 18O content in fluxes of CO2 (Fout) and H2O (E) from leaf to atmosphere. H2O enters the leaf with isotopic composition δs, evaporates from the cell surfaces, and diffuses from the leaf, experiencing both phase-change (ε*) and diffusional (εk) fractionation, giving rise to depleted transpiring water (δt) and enriched evaporating surfaces (δe). Similarly, CO2 from the atmosphere (Fin) dissolves in the chloroplast, equilibrates (εeq) to composition δc depending on the δ18O of water in the chloroplast and the extent of isotopic equilibrium (θeq), and then approximately two-thirds retro-diffuses outward (Fout) with fractionation during diffusion (ā). This can be observed as an 18O enrichment in outgoing CO2 (δout) relative to incoming CO2 (δin), which is proportional to discrimination against C18OO, termed Δ18O. CO2 reference points along the leaf-atmosphere pathway are marked (with average values in μmol mol−1) as cc, ccs, ci, and ca, referring to the CO2 concentration in the chloroplast, chloroplast surface, substomatal cavity, and air, respectively.
The isotopic composition of water at evaporating surfaces (δe) may be estimated from the Craig and Gordon model of evaporative enrichment (Craig and Gordon, 1965):
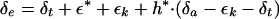 |
3 |
where h* is the relative humidity at leaf temperature; δa and δt are the isotopic composition of water vapor in the air and transpired by the leaf, respectively; εk is the combined diffusional fractionation through stomata and turbulent boundary layer (Farquhar and Lloyd, 1993; Buhay et al., 1996); and ε* is the temperature-dependent liquid-vapor fractionation. The measurement of δ18O of transpired water vapor (δt) allows estimation of δe under non-steady-state conditions (Harwood et al., 1998). While the proximity of chloroplasts to the liquid-air interface in leaves implies good mixing between evaporating sites and chloroplasts, isotopic gradients in leaf water can occur (Yakir et al., 1989, 1994; Luo and Sternberg, 1992; Wang and Yakir, 1995) and need to be considered.
Considering oxygen isotope exchange between CO2-H2O, current estimates have suggested isotopic exchange to be almost complete, approximately 95% (Farquhar and Lloyd, 1993; Flanagan et al., 1994; Williams et al., 1996). However, given the potential uncertainties in δ18O of water and CO2 concentration in the chloroplast when interpreting Δ18O, an independent method is still required to test this assumption. Alternatively, the extent of isotopic equilibrium (θeq) in the CO2-H2O system may be derived from Mills and Urey (1940) as:
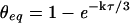 |
4 |
which describes the fractional approach to full equilibrium (where θeq = 1) as a function of the number of hydration reactions achieved per CO2 molecule (kτ). This “coefficient of CO2 hydration” may be calculated for a leaf by calculating the rate constant (k) from biochemical measurements of CA activity and the residence time of CO2 in the leaf (τ) from photosynthetic flux measurements of CO2 (see “Materials and Methods”). In this way, the extent of isotopic equilibrium may be directly determined.
We sought to determine the CO2 concentration relevant to CO2-H2O in leaves from measurements of Δ18O by constraining both the δ18O of exchangeable water and the extent of isotopic equilibrium as above. The subsequent implications toward internal CO2 conductance are discussed in the context of CA activity and its role in facilitating diffusion of CO2 within the chloroplast.
RESULTS AND DISCUSSION
Interpreting C18OO discrimination requires information on δ18O of water in the chloroplasts, the extent of isotopic equilibrium between CO2-H2O and the CO2 concentration in the chloroplast. The δ18O value of chloroplast water is often derived from the Craig and Gordon model for evaporating water (δe in Eq. 3; Flanagan et al., 1994; Williams et al., 1996; Yakir and Wang, 1996; Wang et al., 1998; Harwood et al., 1998). This is due to the proximity of chloroplasts to the liquid-air interfaces within leaves. This leaves two options in using Equation 2 and measurements of Δ18O: (a) to use Δ13C-derived estimates of cc as the CO2 concentration relevant to the site of oxygen exchange in the chloroplast and solve for the extent of isotopic equilibrium (e.g. Flanagan et al., 1994); and (b) to independently measure the extent of isotopic equilibrium and solve for cc. We argue that cc does not refer to the site of CO2-H2O equilibrium and so took the latter approach to estimate its true value. Using constrained estimates of δe and direct assays of CA activity, we solved both Δ18O and Δ13C discrimination equations for cc. We found the Δ18O-derived cc to be always higher than Δ13C-derived values, and define the CO2 concentration relevant to Δ18O as ccs (for [CO2] at the chloroplast surface). We then use ccs to partition the internal conductance into its two major components. In the following sections we show how the interpretations were constrained and discuss their implications.
Observed Δ18O
Consistent with previous observations and predictions, a clear dependence of Δ18O on cc/ca was observed (Fig. 2; Farquhar et al., 1993; Flanagan et al., 1994; Williams and Flanagan, 1996; Williams et al., 1996). As expected, Δ18O was also larger when measured using 18O-depleted source CO2, which generated a larger isotopic difference between source and leaf CO2 (Δca) (increasing the precision of measurement). No difference in the response was observed under different photorespiratory conditions in oak leaves (Fig. 2c). Low pO2 did, however, induce higher assimilation rates and lower ci and cc (Table I) due to reduced photorespiration. Estimates of internal CO2 conductance (gi) derived from Δ13C measurements were 0.50 (±0.12), 0.32 (±0.05), and 0.27 (±0.09) mol m−2 s−1 for tobacco, soy, and oak respectively, which were used to calculate cc/ca. Our gi estimates were in line with previous estimates on similar herbaceous species (von Caemmerer and Evans, 1991; Evans et al., 1994), although in the case of oak, gi was higher than the range previously quoted for oak species (0.15, Loreto et al., 1992; 0.08–0.22 mol m−2 s−1, Roupsard et al., 1996; 0.07–0.08, Hanba et al., 1999).
Figure 2.

Discrimination against C18OO (Δ18O) as a function of chloroplast CO2 concentration (calculated from Δ13C) and expressed as cc/ca for tobacco (a), soy (b), and oak (c). In a and b, experiments were conducted under depleted source CO2 (−30‰, white symbols) and ambient CO2 (0‰, black symbols). For oak, experiments were conducted all in depleted CO2, but at 2% (squares), 21% (circles), and 35% (triangles) oxygen.
Table I.
Gas exchange (at max PPFD), CA activity, and isotopic water data (average at all PPFDs) for tobacco, soy (n = 3 and 2), and oak (n = 3, at different O2%)
| Species | Emax | gs(max) | Amax | ci(min) | Tmax | CAassay | CAleaf | VPD | δt | δe | δLW | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| × 103 | ||||||||||||
| Tobacco 1 | 5.5 | 244 | 14.1 | 274 | 27.2 | n.d | 1,623a | 1.37 (0.19) | −6.3 (0.9) | +12.0 (1.1) | +11.0 | |
| Tobacco 2 | 4.8 | 189 | 13.5 | 240 | 29.3 | 54.8 (13) | 598 | 1.48 (0.17) | −5.5 (0.6) | +15.6 (1.0) | +11.9 | |
| Tobacco 3 | 5.0 | 241 | 15.8 | 265 | 28.5 | 71.3 (6.3) | 670 | 1.27 (0.18) | −2.3 (0.7) | +16.4 (1.1) | +13.6 | |
| Soy 1 | 4.3 | 206 | 12.2 | 255 | 29.3 | 25.3 (2.9) | 294 | 1.51 (0.11) | −4.7 (0.9) | +11.0 (1.1) | +11.0 | |
| Soy 2 | 5.1 | 300 | 20.0 | 237 | 28.5 | 37.1 (1.6) | 342 | 1.32 (0.14) | −5.1 (0.9) | +12.7 (0.3) | +11.9 | |
| Oak - 40% | 3.3 | 142 | 11.8 | 237 | 28.9 | n.d. | – | 1.62 (0.18) | −5.6 (1.9) | +13.4 (1.4) | n.d | |
| Oak - 20% | 4.2 | 166 | 15.5 | 207 | 29.2 | n.d. | – | 1.57 (0.17) | −6.6 (0.6) | +11.8 (0.8) | n.d. | |
| Oak - 2% | 4.1 | 172 | 17.5 | 174 | 27.8 | n.d. | – | 14.6 (2.2) | −6.2 (0.7) | +13.1 (1.3) | n.d. | |
Units are: evaporation rate (Emax, mmol m−2 s−1), stomatal CO2 conductance (gs(max), mmol CO2 m−2 s−1), CO2 assimilation (Amax, μmol m−2 s−1), sub-stomatal CO2 concentration (ci, μmol mol−1), leaf temperature (Tmax, °C), carbonic anhydrase CO2 hydration rate, under assay (CAassay, at 2°C and 35 mm [CO2], mean of three leaves), and in vivo conditions (CAleaf, at ccs and Tleaf, μmol CO2 m−2 s−1). For VPD (kPa) and isotopic data (‰), nos. are averages (+sd) during the entire light response. Symbols as in text.
In vivo CA hydration based on the mean CAassay of the other two tobacco plants.
δ18O of Water at the Site of Oxygen Exchange
Estimates of evaporating surface water (δe) were based on direct measurements of transpired water vapor (δt) applying the isotopic fractionation during evaporation (Eq. 3). The maintenance of constant vapor pressure deficit (VPD) during the complete photosynthetic photon flux density (PPFD) curve kept both δt and δe constant throughout each experiment (Table I). Determining δe in this way from δt avoids uncertainties that arise in substituting source water δ18O for δt in Equation 3, as shown in Table I; although δt is at steady state, the absolute value may differ from source water (−4.5‰) by several per mill.
δe is assumed to provide a close approximation to δ18O of water in the chloroplasts. The proximity of chloroplasts to evaporating surfaces is sufficient to ensure good isotopic mixing between the two. In particular, this assumption would be safe when 18O heterogeneity in the entire leaf water is small. As a precautionary measure, such heterogeneity was evaluated by comparing δe with measured bulk leaf water (δLW) both at the end of each light response experiment (Table I; Fig. 3, white symbols) and across a range of evaporation rates in an independent test (Fig. 3, black symbols). On average, bulk leaf water was lower than δe by 2‰ (±1‰ in Fig. 3). This phenomenon has been observed extensively (Wang et al., 1998, and refs. therein) and has been partly explained by the inclusion of unenriched vein water, estimated to represent 2% to 5% of total leaf water (Yakir et al., 1989; Flanagan et al., 1991), and/or by a Peclet effect proposed by Farquhar and Lloyd (1993).
Figure 3.

The difference between δe from the Craig and Gordon equation and bulk leaf water (δLW) as a function of evaporation rate (E) for soy (triangles), tobacco (circles), maize (diamonds), and sorghum (squares). White symbols are data from the last measurement of the light response study; black symbols are additional points from the leaf water heterogeneity test. The three marked points excluded from statistical analysis are thought to represent non-steady-state conditions.
The difference between δe and δLW increased with the evaporation rate (Fig. 3, excluding the three marked data points, from leaves thought not to be at isotopic steady state), which is consistent with a Peclet effect (Flanagan et al., 1991; Farqhuar and Lloyd, 1993). In this case, the large advective flux of water through the leaf at higher evaporation rates restricts back-mixing of 18O-enriched water from the evaporation sites with the bulk leaf water. The maximal Peclet effect observed here was 3‰ (excluding marked points), which was much smaller than those reported previously (Flanagan et al., 1991, 1994; Wang et al., 1998). The small Peclet effect in this study was probably due to low evaporation rates (<5 mmol H2O m−2 s−1, Table I). In addition, although not measured here, Peclet effects in oak (K.G. Harwood, D. Yakir, J.S. Gillon, and H. Griffiths, unpublished data) and other woody species (birch and poplar; Roden and Ehleringer, 1999) have been consistently equally low in a wide range of conditions. Therefore, the absence of significant isotopic gradients in leaf water over the whole leaf is a good indication that isotopic gradients are unlikely to occur across the much smaller distances (<0.01 mm) between evaporating surfaces and chloroplasts.
Extent of CO2-H2O Isotopic Equilibrium
We measured CA activity in the experimental plants and used the results to estimate the in vivo extent of isotopic equilibrium between CO2 and H2O in the chloroplast. Previously, close to full isotopic equilibrium has been assumed due to the high rates of CA catalysis expected in most plants. We tested this assumption by measuring CA activities under assay conditions and estimating in vivo rates under leaf conditions (at chloroplastic [CO2] and leaf temperature). Assay rates showed significant variation, with the lowest rates in soy (Table I). Although CA activity for the oak plants used here was not determined, measurements in oak species from previous studies revealed very high CA activity (mean and sd for Quercus bosserii −288 ± 36; Quercus robur −261 ± 25; Quercus pedunculata −201 ± 30 mmol CO2 m−2 s−1; J.S. Gillon and D. Yakir, unpublished survey data). Further differences were introduced when calculating in vivo CO2 hydration rates, due to small variabilities in leaf temperature and internal [CO2] between the experiments. Notably, during a light response curve, CO2 hydration rates in vivo did not vary considerably. Most likely, reductions in the calculated CO2 hydration rate associated with decreased internal [CO2] at high light were compensated for by increased leaf temperatures and enhancement of catalytic activity.
Using the data on CA activity during leaf gas exchange, we could assess the extent of CO2-H2O isotopic equilibrium independently of Δ18O measurements. The efficiency of CO2-H2O isotopic equilibrium depends upon the product kτ, which is residence time (τ) times the rate constant of CO2 hydration within the chloroplast (k) (see “Materials and Methods”). Combining with the isotope exchange theory of Mills and Urey (1940) for the CO2-H2O reaction, the fractional extent of isotopic equilibrium may be described by θeq (Eq. 4), so that full equilibrium occurs when θeq = 1 (corresponding to kτ greater than 15; Fig. 4). Thus, as CO2 (with isotopic signal of δa) passes through a leaf, the δ18O of CO2 changes, approaching equilibrium with leaf water represented by δe. The δ18O value of CO2 in the chloroplast (δc) should lie between δa and δe, at some point depending on θeq (Fig. 4). This effect has already been demonstrated qualitatively in genetically modified plants with low CA activity, where Δ18O, and therefore δc, were dramatically reduced compared with the values expected at full isotopic equilibrium (δe) (Price et al., 1994; Williams et al., 1996).
Figure 4.

Diagram showing the dynamics of oxygen isotope exchange between atmospheric CO2 (δa) and leaf water (δe) and the resulting δ18O of CO2 (δc). Isotopic equilibrium (θeq) from Equation 4, solid line was calculated from the CA activity and CO2 residence time (kτ), which represents the number of hydrations per CO2 molecule and is related to Δca/Δea.
Calculating kτ and the corresponding extent of isotopic equilibrium for each data point, we observed more than 95% isotopic equilibrium (θeq >0.95) for tobacco (Fig. 5). The lower CA activity in soy suggested that the equilibrium was 75% complete. In the extreme case, potentially high CA activity in oak corresponded to complete isotopic equilibrium throughout the light response, where kτ was always greater than 15 (data not plotted in Fig. 5). Note that in other plants, including C3 and C4 species, θeq values were found to span the whole range from 0 to 1 (J.S. Gillon and D. Yakir, unpublished data).
Figure 5.

The number of hydration reactions per CO2 molecule (kτ) calculated from CAleaf/Fin as a function of the CO2 assimilation rate. Shown on the second axis is the equivalent extent of isotopic equilibrium from Equation 4, in which full equilibrium (>99.5%) occurs above kτ = 15. White symbols, Soy; black symbols, tobacco (different symbols refer to different light responses). All values for oak were above kτ = 15 because of the assumed high CA rates.
CO2 Concentration at the Site of Isotopic Equilibrium
Based on the above discussion that δc lies between δa and δe and reflects θeq, it is possible to show that θeq is related to Δca/Δea (Fig. 4), since algebraically:
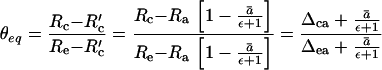 |
5 |
This describes the 18O/16O ratio of CO2 at the site of oxygen exchange (Rc) relative to that in full equilibrium with leaf water (Re), and that for un-equilibrated CO2 inside the leaf, Rc′. The term Rc′ = Ra · (1−ā/[ε+1]) allows for the variable expression of ā under non-equilibrium conditions (as in 13C discrimination). Thus, (1−ekτ/3) = θeq ≈ Δca/Δea (Fig 5). We now incorporate the extent of isotopic equilibrium into Eq. 2, and C18OO discrimination is then given as:
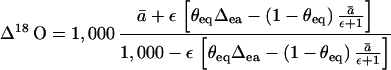 |
6 |
With measured values of Δ18O, Δea, and θeq, we may thus derive ε, and hence cc(eff), the effective CO2 concentration at the site of CO2-H2O equilibrium.
The cc(eff) values obtained from Equation 6 were always intermediate between ci (from gas exchange) and cc (from Δ13C), as shown in Figure 6. These results were obtained from eight experiments in three species, measured on two separate systems, reducing the likelihood of bias introduced via system or species effects. Typically, all values of internal [CO2] dropped at high assimilation rates, as CO2 demand increased, albeit with variation due to some non-correlated changes in stomatal conductance, particularly for soy. Species differences were evident: cc(eff) was generally closer to ci in soy, closer to cc in oak, and intermediate in tobacco.
Figure 6.

Data shown for all light response curves, showing internal CO2 concentration (μmol mol−1) as a function of CO2 assimilation rate (A) (μmol m−2 s−1). Symbols refer to CO2 concentration in internal air space (ci) (from gas exchange measurements, ▵), ccs (from Δ18O, δe, and CA activity; ♦), and cc (from Δ13C; ○). Species are tobacco (a–c), soy (d–e), and oak (f–h). Light responses in c and e were conducted using ambient δ18O CO2, whereas the rest used CO2 depleted in 18O. f through h, Experiments in 40%, 20%, and 2% ) O2, respectively.
The values of cc(eff) represent the CO2 concentration at the effective site of CO2-H2O equilibrium, which we term ccs (Yakir, 1998), indicating that the chloroplast surface is the likely site. This assumes that the limit of CA activity occurs at the chloroplast surface, since the majority of CA resides within the chloroplast (Everson, 1970). These results are consistent with the difference between the effects of Rubisco on Δ13C and CA on Δ18O: although Rubisco and CA show the same distribution within the chloroplast (Anderson et al., 1996), Rubisco removes 12CO2 from the system, creating a 13C gradient between cc and ca; CA only acts to cancel out any 18O gradients in CO2 throughout the domain of its activity, so that an 18O gradient only exists from the chloroplast surface (ccs) to the atmosphere (ca) (Fig. 7).
Figure 7.

Diagram representing the backflux of CO2 from sites of CO2 fixation (cc) and sites of oxygen exchange (ccs) in the chloroplast, showing the partitioning of total internal conductance (gi) (relevant to Δ13C) into chloroplast (gch) and wall (gw) conductance (from Δ18O).
Note that in interpreting Δ18O, the best-constrained value is δe. Consequently, testing the model for Δ18O usually involved deriving δc values and comparing them with δe values. Assuming that we appropriately adjust δc for θeq and correctly estimate ccs, the two values should match. In previous studies, 13C-derived values of cc were used and the observed difference between δe and δc was explained in terms of incomplete isotopic equilibrium. The effects of incomplete equilibrium was addressed in those cases by applying a certain ρ value (ρ = A/CAleaf), which was incorporated within the Δ18O model (Farquhar and Lloyd, 1993; Flanagan et al., 1994; Williams and Flanagan, 1996; Williams et al., 1996). Furthermore, the method of cc determination based on individual Δ13C measurements (and not the trend of Δ13C across the full range of A) generated a range of cc values (up to 40 μmol mol−1), so that co-adjustment of cc and ρ was required to resolve δe versus δc differences.
In some cases, estimates of δc were as much as 10‰ below δe in both laboratory and field studies (Yakir et al., 1994; Harwood et al., 1998; Wang et al., 1998). Such differences cannot be explained by only considering ccs, and probably imply large heterogeneity in leaf water isotopic composition. Especially in the two latter field studies, Peclet effects may be much larger than observed in this study. It is possible that these discrepancies represent isotopic leaf water heterogeneity between water in the chloroplast and at the evaporating sites. Better characterization of the oxygen exchange site may help future studies of significant leaf water gradients and Peclet effects.
Partitioning Internal CO2 Conductance
The association of Δ18O with the [CO2] at the chloroplast surface (ccs) provides us with another reference point in the diffusion pathway from atmosphere to chloroplast in addition to probing ci via gas exchange (von Caemmerer and Farquhar, 1981) and cc from Δ13C analysis (von Caemmerer and Evans, 1991). From Fick's law of diffusion, CO2 concentration gradients are related to conductance by the general expression A = gx (c1 − c2). Applying values of ccs, we may partition the total conductance (gi) into its components before and after the chloroplast surface by plotting A versus (ci − ccs) and versus (ccs − cc). In each case, the inverse of the gradient refers to cell wall/plasmalemma conductance (gw) and conductance within the chloroplast (gch), respectively (Fig. 7), assuming no significant resistance to CO2 diffusion in the gaseous leaf interior (Evans et al., 1994). Despite a larger error in determining conductances from Δ18O compared with Δ13C, gw was significantly higher than gi for both tobacco and soy, and on the borderline of significance for oak (Table II). Comparing the values of gch relative to gw, the chloroplast conductance was estimated to be 0.8 (tobacco), 0.3 (soy), and 3.2 (oak) times the wall conductance (gch/gw, Table II). The magnitude and species variability of gch was much lower than previous theoretical estimations, where the wall conductance was thought to be the major limitation to diffusion, such that gch/gw was predicted to be from 4.8 (Evans et al., 1994) to 7.4 (Cowan, 1986). The occurrence of low chloroplast conductance was associated with low in vivo CA activities (soy), while potentially high CA activity in oak may be associated with high gch.
Table II.
The breakdown of total leaf CO2 conductance (gleaf) into its components, stomatal (gs) (from gas exchange) and internal (gi) (from Δ13C, plus error from 95% confidence limits of the slopes)
| Species | gleaf | ||||||
|---|---|---|---|---|---|---|---|
|
gs |
gi |
||||||
|
gw |
gch |
gch/gw | CAleaf | ||||
| Tobacco | 0.155 | 0.224 | 0.50 (0.12) | 1.12 (0.33) | 0.90 | 0.8 | 623 |
| Soy | 0.141 | 0.253 | 0.32 (0.05) | 1.31 (0.45) | 0.42 | 0.3 | 318 |
| Oak | 0.102 | 0.166 | 0.27 (0.09) | 0.35 (0.04) | 1.13 | 3.2 | 2,016a |
With Δ18O, gi is further partitioned into wall conductance to Ccs, gw, and the residual conductance within the chloroplast, (gch). All units are mol CO2 m−2 s−1. The ratio of chloroplast to wall conductance is also shown gch/gw. Average CAleaf activity is shown for comparison (μmol m−2 s−1).
In the absence of direct CA measurements in the oak species used here, CAleaf is estimated from the average CA activity observed other oak species (see text).
Importance of CA-Mediated Diffusion in gch
It is becoming increasingly evident that CA facilitates diffusion and therefore CO2 conductance within the chloroplast (Cowan, 1986; Makino et al., 1992; Price et al., 1994; Williams et al., 1996; Sasaki et al., 1998). This may be further supported by the association of CA activity with the relative magnitude of chloroplast conductance in the three species used here (Table II). However, in the past, modification of CA activity has revealed little or no change in photosynthetic rate (Price et al., 1994; Williams et al., 1996), so that the benefit to photosynthesis from CA remains unclear. We now propose that the relative contribution from CA to photosynthetic efficiency may be species dependent and not always clearly apparent. In particular, CA-mediated diffusion may be more important when total internal conductance is low, as is the case for woody species (von Caemmerer and Evans, 1991; Lloyd et al., 1992; Loreto et al., 1992; Syvertsen et al., 1995). In such cases, photosynthetic limitations attributable to low wall conductance (gw), which occur due to the cellular architecture of schlerophyllous leaves, may be offset by optimizing chloroplast conductance (gch).
This species effect on CA-mediated gch is illustrated by estimating CO2 assimilation as a function of chloroplast conductance (Fig. 8). Assimilation was described as A = k(cc − Γ*) − Rd, where k and Γ* are the carboxylation efficiency and compensation point (k = 0.121 and 0.073, ci = 208 and 252, Γ* = 40 and 45 for oak and tobacco, respectively) and cc = ci − A/gi. We calculated the change in CO2 assimilation rate relative to observed values (Fig. 8) due to varying the chloroplast component of internal conductance (while keeping gw constant, plotted as gch/gw in Fig. 8). In oak, with lower wall conductance (high gch/gw), the current assimilation rate is 20% higher compared with that which would occur at chloroplast conductance values found in tobacco. Conversely, in tobacco, increasing gch to the extent found in oak would result in only a 5% increase in A. This example is also consistent with the gas exchange measurements from tobacco plants with genetically reduced CA activity (Price et al., 1994; Williams et al., 1996). Internal conductance was lower (approximately 0.25 mol m−2 s−1) in the CA mutant compared with wild-type plants (approximately 0.4 mol m−2 s−1).
Figure 8.

The potential change in CO2 assimilation rate (A) as a function of gch (oak and tobacco, solid lines). gch is normalized relative to a constant wall conductance (gw) (0.35 and 1.12 mol m−2 s−1 for oak and tobacco, respectively). The changes in A are expressed relative to measured assimilation rates at actual conductance values, gch/gw = 0.8 and 3.2 for tobacco (A = 12.7 μmol mol−1) and oak (A = 13.7 μmol mol−1), respectively. Also marked is the estimated gch/gw (see text) of tobacco mutants lacking CA (Price et al., 1994; Williams et al., 1996), indicating only a small effect on assimilation relative to the wild-type tobacco.
Applying the present ratio of gch/gw (0.8) for wild-type tobacco plants, we may calculate the wall conductance for wild-type plants from their Δ13C-derived gi values. Assuming this physical wall conductance is unchanged between wild-type and CA mutant plants (the antisense CA gene should have no other effects on leaf physiology and structure), we estimate a lower value of gch/gw = 0.5 for the CA mutant tobacco plants, i.e. the reduction in gi is due to reduction in gch only. Indicating the position of the CA-mutant plants on Figure 8, we predict only a 5% drop in CO2 assimilation for the 90% to 95% reduction in CA activity, in agreement with reported results.
Two main points arise from this simple analysis. First, it appears that relative chloroplast conductance is proportional to CA activity across almost three orders of magnitude of CA activity, with a possible minimum at gch/gw = 0.5, where all residual CO2 diffusion will be un-facilitated (i.e. no CA effect). This strongly supports the occurrence of CA-mediated diffusion in the chloroplast. Second, although the oak plants used may not be completely representative of woody species, CA activity in woody plants in general may have been optimized over evolutionary time to compensate for low wall conductance (J.S. Gillon and D. Yakir, unpublished data). For example, in a preliminary survey, mean in vivo CA hydration rates were 1,090 and 390 μmol m−2 s−1 for trees/shrubs (n = 16) and herbaceous species (n = 12), respectively, which may correspond to a three-times increase in gch relative to gw. By extending such surveys to include conductance estimates (both internal and stomatal), or by manipulating CA activity in species with low internal conductance, the potential importance of CA in photosynthesis may prove to be substantially greater than currently assumed.
MATERIALS AND METHODS
Plant Material
Soy (Glycine max), tobacco (Nicotiana tabacum), maize (Zea mays), and sorghum (Sorghum bicolor) were grown from seed in a greenhouse under ambient light and temperature at Weizmann Institute of Science (WIS). The latter two species were only used in the test of leaf water heterogeneity to increase the scope of the test. Oak seedlings (Quercus robur) were provided by the Forestry Commission (UK) in 1991, and kept outside and well-watered in 1-L pots at Moorbank Botanical Gardens (University of Newcastle-upon-Tyne, UK), until required. Measurements were conducted on 6- to 10-week-old plants of soy and tobacco plants and on 5-year-old oak seedlings. Transfer of plant material was several days prior to the experiment to allow acclimatization to laboratory conditions.
System 1 (WIS): Gas Exchange
Figure 9 shows a scheme of the on-line isotope/gas exchange system at WIS. Synthetic air was mixed from N2, CO2, and O2 cylinders using mass flow controllers (MKS Instruments, Andover, MA), and humidified by bubbling a variable portion of the airstream through water at room temperature (δ18O = −4.5‰, therefore, vapor ≈ −14.5‰), acidified with two drops of 80% (v/v) H3PO4. The airflow was split into reference and analysis airstreams, the latter flow range, 800 to 1,500 mL min−1, was passed to a Parkinson “conifer pod” leaf cuvette (PLC) (model PLC3C, ADC Scientific, Hoddeson, UK), and flow was measured via another mass flow controller. Illumination was from a 250 W projector lamp (GEC, Cleveland), passing through a 3-cm depth of water to reduce infrared radiation. Incident radiation on the leaf was controlled by shading with a predetermined number of Miracloth filters (Calbiochem, San Diego). Absolute CO2 and H2O concentration in reference and analysis airstreams were monitored alternately via an infrared gas analyzer (model Li-6262, LI-COR, Lincoln, NE).
Figure 9.

Arrangement of on-line CO2 trapping and off-line H2O trapping apparatus for continuous flow CO2 isotopic analysis, in conjunction with leaf chamber and gas exchange system.
Isotopic Measurement of CO2
The outflow of the leaf chamber after passing through the infrared gas analyzer (minimum 700 mL min−1) was split, 100 mL min−1 was pumped first through a dryer (Nafion, Perma Pure, Toms River, NJ), and then a sample loop (0.85 mL) was fitted onto a six-port, two-position valve (Valco Instruments, Houston). CO2 was trapped at liquid N2 temperatures for 30 s. After warming to room temperature, the sample was swept with helium carrier gas (120 mL min−1; ultrapure, Gordon Gas and Chemicals, Tel Aviv) through a magnesium perchlorate drying trap and a 2-m packed column (sieve 5A, 80/100 mesh, Alltech, Deerfield, IL) at 60°C. The large peaks of N2 and O2 that eluted first from the column were diluted via a gas diluter (Micromass, Manchester, UK), followed by the non-diluted sample CO2. The gas was introduced into the source of a mass spectrometer (OPTIMA, Micromass) via an open split. 13C to 12C and 18O to 16O isotope ratios were measured from the integrated peak areas of masses 44, 45, and 46 normalized against a 30-s CO2 reference pulse injected prior to each sample. Sample size was standardized by adjusting the cryogenic trapping time according to the CO2 concentration in the outflow from the leaf chamber. N2O was assumed to be constant in air (310 nmol mol−1) and absent from “synthetic” air, so δ values were corrected accordingly (Freidli and Siegenthaler, 1988) and expressed in the small delta notation versus Vienna Pee Dee Belemnite (VPDB) for 13C and VPDB-CO2 for 18O. Precision for repeated sampling of CO2 was 0.06‰ (δ13C) and 0.07‰ (δ18O).
Isotopic Measurement of Water Vapor
The remaining airflow from the leaf chamber was passed at positive pressure to a 0.61-cm o.d. stainless steel vacuum line (pressure <1 × 10−3 torr) in which CO2 and water vapor were trapped from the airstream (3 min at 500 mL min−1) in a coil cooled with liquid N2. After trapping, the line was evacuated and the trap was heated with a flame, distilling both CO2 and H2O into a Pyrex side arm immersed in liquid N2. After quantitative transfer the pyrex tube was flame sealed. The sample was left for CO2-H2O equilibrium at constant temperature (29°C, Labline Instruments, Melrose Park, IL) for 72 h. The CO2 was then dried in a vacuum line with an ethanol trap at −70°C before isotopic analysis on a dual inlet mass spectrometer (MAT 250, Finnigan-MAT, Bremen, Germany). δ18O of water vapor was calculated from that of the CO2 according to the method of Scrimgeour (1995), correcting for the amount CO2 and H2O (calculated from the concentration, flow rate, and time of trapping) and the δ18O of the pre-equilibration CO2, taken from the corresponding measurement of the continuous flow system. Precision of δ13C CO2 and δ18O water vapor was 0.04‰ and 0.11‰, respectively.
Experimental Procedure
Light responses were conducted from high to low PPFD (1,200–100 μmol photons m−2 s−1, 10 intervals) in 21% O2. Collections of CO2 for isotopic analyses were carried out for 3 min, while water vapor was trapped continuously (i.e. two samples of CO2 and one of water were analyzed per light level). Photosynthesis measurements were averaged for the collection period. At the end of the experiment, the portion of leaf inside the cuvette was excised and placed in a 15-mL vacuum container (Becton-Dickinson, Rutherford, NJ) for extraction of leaf water. In addition, three leaf discs (1.8 cm2) were cut from the same leaf, and stored in liquid N2 for subsequent determination of CA activity. The complete light response analysis (approximately 10 determinations) was first conducted with CO2 relatively depleted in 13C and 18O (δ13C = −30‰ and δ18O = −30‰) to maximize the precision of measurement. Subsequently, ambient air pumped through a 50-L external buffering volume (δ13C ≈ −8‰ and δ18O ≈ 0‰) was used to replicate the experiment. Run-replicate numbers were n = 3 for tobacco (two “depleted” and one ambient air) and n = 2 for soy (one of each).
System 2 (UNUT)
Photosynthesis measurements and cryogenic trapping of CO2 and H2O for the experiments on oak were conducted using the CIRAS-1 (PP Systems, Hitchin, UK) and collection system at UNUT, which is described in Gillon and Griffiths (1997). CO2 isotopic composition was δ13C = −42‰, δ18O = −30‰, with δ18O water vapor approximately = −18‰. In addition, trapped CO2 and H2O were separated via distillation of CO2 from the mixture using an acetone/liquid N2 slush at −80°C, as described in Harwood et al. (1998). Precision for dry CO2 was 0.04‰ (δ13C) and 0.07‰ (δ18O). Precision for δ18O H2O determinations was 0.09‰.
Experimental Procedure
A portion of leaf was placed in the chamber and illuminated for 1 to 2 h before beginning measurements. During sampling, CO2 and water vapor were cryogenically trapped for 15 min from an airstream of 200 mL min−1, during which time photosynthetic parameters were averaged. This was repeated at various PPFDs (500–100 μmol photons m−2 s−1, 10–12 steps) to cover the range of CO2 assimilation from approximately 5 μmol m−2 s−1 to saturation, allowing the photosynthetic rate to stabilize between each change in PPFD (approximately 20 min). The leaf-to-air VPD was maintained as constant as possible (approximately 1.5 ± 0.2 kPa) throughout the experiment by drying a portion of the reference airstream with Drierite (W.A. Hammond, Xenia, OH). Reference CO2 was collected between every three to four samples. The complete light response and isotopic analyses were conducted on the same leaf three times, once each at 2%, 21%, and 35% O2 to check the influence of photorespiration rate (all other experiments at WIS were conducted at 21% O2).
Leaf Water Heterogeneity
A separate experiment was carried out at WIS to determine the suitability of the Craig and Gordon model to estimate the δ18O of bulk leaf water (δLW). A leaf was placed in the cuvette and left for 1 h (the minimum time for the first measurement in the above light response experiments). A dry CO2 sample from the leaf chamber was first collected in the stainless steel line by passing the airstream through an additional acetone/liquid N2 trap at −70°C in the vacuum line. This was used to derive the δ18O CO2 to be used for equilibration. Next, a water vapor sample from the leaf chamber was collected, as before, and the leaf portion in the cuvette was then excised immediately afterward, and placed in a vacutainer. Finally a reference water vapor sample, bypassing the chamber, was collected (δ18O ≈ −15‰). This was repeated at different PPFD (100–1,500 μmol photons m−2 s−1) to generate a range of evaporation rates, and repeated for soy, tobacco as well as sorghum and maize, to increase the species range.
Determination of δ18O Leaf Water
Leaf water was extracted by vacuum distillation at 60°C. δ18O values were determined by equilibration of 0.2 mL with CO2 (70 kPa) at 29°C for 24 h, followed by cryogenic separation of a CO2 aliquot, prior to mass spectrometric analysis. Values were calibrated on the Vienna standard mean oceanic water (VSMOW) scale by simultaneously running internal water standards (WIS H2O = −4.5‰ VSMOW).
CA Activity
Leaf discs were ground in a pestle and mortar at 4°C with approximately 1 mL of extraction buffer per square centimeter of leaf disc (adapted from Makino et al., 1992). The buffer contained 50 mm 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES)-NaOH (set to pH 8.3) 0.5 mm EDTA, 10 mm dithiothreitol, 10% (v/v) glycerol, and 1% (v/v) Triton X-100 to ensure solubilization of any membrane-bound CA. Extracts were spun at 5,000 rpm for 10 min, and the supernatant was decanted into Eppendorf tubes and frozen at −20°C until assayed. The assay was conducted in a stirred flat-bottomed tube at 2°C. Assay error for the same extract was much smaller than between extracts, so extracts were only assayed once. To 3 mL of assay buffer (20 mm Na-barbitol at pH 8.3), 15 to 50 μL of extract was added, and the assay was started by adding 1 mL of distilled water previously saturated with CO2 at 0°C. The time for the change from pH 8.3 to 7.3 was recorded. To convert this into a molar rate of CO2 hydration, the same pH change was titrated with 0.2 n H2SO4, assuming the stoichiometry of 1 mole of H+ formed for every mole of CO2 hydrated (Hatch and Burnell, 1990). To calculate the rate of enzymic CO2 hydration, the rate of a blank assay (i.e. just adding extraction buffer) was subtracted from the rate with leaf extract. The activity (CAassay) was expressed on a leaf area basis (micromoles of CO2 hydrated per m2 per second), representing activity at 2°C and 17.5 mm CO2. In vivo rates (CAleaf) were calculated at leaf temperature by CAleaf = (17.5 × F·CAassay·[CO2])/{[17.5 + Km]([CO2]+Km)}, where [CO2] is the concentration at the site of catalysis (from ccs—see “Discussion”—and converted from micromoles per mole to micromolar via Henry's law), Km is the concentration at half maximal activity, taken as 5 mm for dicot CA (data not shown), and the factor F was used to correct the rate to leaf temperature, where F = [(tleaf − tassay)/10]Q10, assuming Q10 = 2 (Hatch and Burnell, 1990).
Coefficient of CO2 Hydration (kτ)
Two main factors control the exchange of 18O between leaf water and CO2. During photosynthesis, the gross CO2 influx rate (Fin) regulates the residence time (τ) of CO2 in the aqueous leaf medium, while the CA-catalyzed hydration of CO2 (CAleaf) determines the efficiency of oxygen exchange during that time (Eq. 4). The rate constant for CA (k) is equivalent to CAleaf/ccs, and the CO2 residence time (τ) is given by ccs/Fin. Thus the product, kτ = CAleaf/Fin, relates to the number of hydration reactions per CO2 molecule. Fin was determined from the product of external CO2 concentration (ca) and total conductance to the site of CO2-H2O equilibrium (ca·gleaf), where gleaf is the combination of boundary (gb), stomatal (gs), and internal conductance to ccs (gw). Using the resistance analogy, gleaf = 1/(1/gb + 1.6/gs + 1.6/gw). Boundary values are quoted in the methods, stomatal values were taken from gas exchange measurements (converting from water to CO2 via the factor of 1.6), and the internal conductance estimate is described below (and see “Discussion”). Note that Fin may be also defined as A(ε + 1) from rearranging A = gleaf (ca − ccs), where ε = ccs/(ca − ccs).
Photosynthetic Calculations
Photosynthetic parameters were calculated according to the method of von Caemmerer and Farquhar (1981). Due to the influence of oxygen on water vapor determination from infrared gas analyzers (Ludwig et al., 1998), measurements in 2% and 35% O2 were corrected with an O2-dependent calibration coefficient (determined from separate tests). Conductance was corrected for the ratio of stomatal density between upper and lower surfaces (K. Parkinson, CIRAS manual, PP Systems, Hitchin, UK), which were measured from epidermal impressions as 0.32, 0.21, and 0.0 for tobacco, soy, and oak, respectively. Boundary layer conductance to H2O was measured for each species by dipping a leaf in a weak detergent solution, removing excess water, then measuring the evaporation rate in a darkened cuvette (1.1, 0.9 mol m−2 s−1 for tobacco and soy in a PLC conifer pod, and 3.0 mol m−2 s−1 for oak in PLC leaf chamber, respectively). Leaf temperature was allowed to vary with PPFD, so that maximal Tleaf at saturating PPFD was between 27°C and 29°C for all experiments (minimum Tleaf = 23°C). Tleaf was calculated from the energy balance, where radiation and transmission characteristics were either taken from the CIRAS manual for the system at UNUT or measured directly at WIS. The ratio of PPFD (Delta-Ohm, Padova, Italy) to total radiation (LI-COR) was determined for the light source, as well as the transmission of the cuvette windows. All of the above parameters can greatly influence the energy budget and hence the calculation of photosynthetic parameters, especially ci, hence we stress that such rigorous determination of all parameters was essential for interpreting plant isotope discriminations.
Δ13C Estimation of cc
The additional reduction in CO2 concentration from ci to the chloroplast (cc) was estimated from the difference between the simple model and the measured discrimination (Δi − Δobs) (Evans et al., 1986) as:
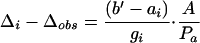 |
7 |
where Δobs is the discrimination measured in Equation 1, Δi = a + (b′ − a) ci/ca (Farquhar et al., 1982), where ci and ca refer to CO2 concentration in the substomatal cavity and atmosphere, respectively, a is the fractionation during diffusion in air (4.4‰), and b′ is the fractionation during carboxylations (29‰); gi refers to the total internal conductance, ai is the combined fractionation (+1.8‰) during dissolution (+1.1‰) and diffusion through the liquid phase (+0.7‰). Internal conductance was derived from the gradient of the Δi − Δobs response versus A/ca measured concurrently with Δ18O during the light responses. This method avoids any influence from photorespiration (Gillon and Griffiths, 1997) and uncertainty in b′. Measurements where A < 8 μmol m−2 s−1 were excluded to avoid the influence of dark respiration on Δ13C (Gillon and Griffiths, 1997).
Statistical Analysis
For the determination of total internal conductance (gi) (from A/ca versus Δi − Δobs) and wall conductance (gw) (from A versus ci − ccs), linear regressions were obtained by the least square method, also deriving 95% confidence limits for slopes, from which the error for each conductance estimate was derived (Sokal and Rohlf, 1981).
ACKNOWLEDGMENTS
We are grateful for the technical support of Emanuela Negreanu and Ruti Yam, and for the reviewers' comments.
Footnotes
This research was supported by the Israel Science Foundation (grant no. 308/96). J.S.G. was supported by a fellowship from the Leverhulme Trust, UK (no. SAS/30317) while at the Weizmann Institute of Science, and by the Natural Environment Research Council, UK (grant no. GT4/94/379) while at University of Newcastle upon Tyne, Department of Agriculture and Environmental Sciences, Ridley Building, Newcastle upon Tyne, UK.
LITERATURE CITED
- Anderson LE, Gibbons JT, Wang X. Distribution of ten enzymes of carbon metabolism in pea (Pisum sativum) Int J Plant Sci. 1996;157:525–538. [Google Scholar]
- Buhay WM, Edwards TWD, Aravena R. Evaluating kinetic fractionation factors use for ecologic and paleoclimatic reconstruction from oxygen and hydrogen isotope ratios in plant water and cellulose. Geochim Cosmochim Acta. 1996;60:2209–2218. [Google Scholar]
- Ciais P, Denning AS, Tans PP, A BJ, Randall DA, Collatz GJ, Sellers PJ, White JWC, Trolier M, Meijer HAJ, Francey RJ, Monfray P, Heimann M. A three dimensional synthesis study of δ18O in atmospheric CO2. Part 1: Surface fluxes. J Geophy Res. 1997;102:5873–5883. [Google Scholar]
- Cowan IR. Economics of carbon fixation in higher plants. In: Givnish TJ, editor. On the Economy of Plant Form and Function. Cambridge, UK: Cambridge University Press; 1986. pp. 133–170. [Google Scholar]
- Craig H, Gordon HI. Deuterium and oxygen-18 variations in the ocean and marine atmoshpere. In: Tongiorgi E, editor. Proceedings of a Conference on Stable Isotopes in Oceanographic Studies and Palaeotemperatures. Laboratory of Geology and Nuclear Science, Pisa. 1965. pp. 9–130. [Google Scholar]
- Evans JR, Sharkey TD, Berry JA, Farquhar GD. Carbon isotope discrimination measured concurrently with gas exchange to investigate carbon dioxide diffusion in leaves of higher plants. Aust J Plant Physiol. 1986;13:281–292. [Google Scholar]
- Evans JR, von Caemmerer S, Setchell BA, Hudson GS. The relationship between CO2 transfer conductance and leaf anatomy in transgenic tobacco with a reduced content of Rubisco. Aust J Plant Physiol. 1994;21:475–495. [Google Scholar]
- Everson RG. Carbonic anhydrase and CO2 fixation in intact chloroplasts. Phytochemistry. 1970;9:25–32. [Google Scholar]
- Farquhar GD, Lloyd J. Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere. In: Ehleringer JR, Hall AE, Farquhar GD, editors. Stable Isotopes and Plant Carbon-Water Relations. San Diego: Academic Press; 1993. pp. 47–70. [Google Scholar]
- Farquhar GD, Lloyd J, Taylor JA, Flanagan LB, Syvertsen JP, Hubick KT, Wong SC, Ehleringer JR. Vegetation effects on the isotope composition of oxygen in atmospheric carbon dioxide. Nature. 1993;363:439–443. [Google Scholar]
- Farquhar GD, O'Leary MH, Berry JA. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust J Plant Physiol. 1982;9:121–137. [Google Scholar]
- Flanagan LB, Compstock JP, Ehleringer JR. Comparison of modeled and observed environmental influences on the stable oxygen and hydrogen isotope composition of leaf water in Phaseolus vulgaris L. Plant Physiol. 1991;96:588–596. doi: 10.1104/pp.96.2.588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flanagan LB, Phillips SL, Ehleringer JR, Lloyd J, Farquhar GD. Effect of changes in leaf water oxygen isotopic composition on discrimination against C18O16O during photosynthetic gas exchange. Aust J Plant Physiol. 1994;21:221–234. [Google Scholar]
- Francey RJ, Tans PP. Latitudinal variation in oxygen-18 of atmospheric CO2. Nature. 1987;327:495–497. [Google Scholar]
- Freidli H, Siegenthaler U. Influence of N2O on isotope analysis in CO2 and mass-spectrometric determination of N2O in air samples. Tellus. 1988;40B:129–133. [Google Scholar]
- Gillon JS, Griffiths H. The influence of (photo) respiration on carbon isotope discrimination in plants. Plant Cell Environ. 1997;20:1217–1230. [Google Scholar]
- Gonfiantini R, Gratziu S, Tongiogi E. Use of Isotopes and Radiation in Soil-Plant Nutrition Studies. IAEA, Vienna. 1965. Oxygen isotope composition of water in leaves; pp. 405–410. [Google Scholar]
- Hanba YT, Miyazawa S, Terashima I (1999) The influence of leaf thickness on the CO2 transfer conductance and leaf stable carbon isotope ratio for some evergreen tree species in Japanese warm temperate forests. Funct Ecol (in press)
- Harwood KG, Gillon JS, Griffiths H, Broadmeadow MSJ. Diurnal variation of Δ13CO2, ΔC18O16O and evaporative site enrichment of δH218O in Piper aduncum under field conditions in Trinidad. Plant Cell Environ. 1998;21:269–283. [Google Scholar]
- Hatch MD, Burnell JN. Carbonic anhydase activity in leaves and its role in the first step of C4 photosythesis. Plant Physiol. 1990;93:825–828. doi: 10.1104/pp.93.2.825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lloyd J, Syvertsen JP, Kriedemann PE, Farquhar GD. Low conductances for carbon dioxide diffusion from stomata to the sites of carboxylation in leaves of woody species. Plant Cell Environ. 1992;15:873–899. [Google Scholar]
- Loreto F, Harley PC, Di MG, Sharkey TD. Estimation of mesophyll conductance to carbon dioxide flux by three different methods. Plant Physiol. 1992;98:1437–1443. doi: 10.1104/pp.98.4.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludwig M, von Caemmerer S, Price D, Badger M, Furbank RT. Expression of tobacco carbonic anhydrase in the dicot Flaveria bidentis leads to increased leakiness of the bundle sheath and a defective CO2 concentrating mechanism. Plant Physiol. 1998;117:1071–1081. doi: 10.1104/pp.117.3.1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo Y, Sternberg LSL. Spatial D/H heterogeneity of leaf water. Plant Physiol. 1992;99:348–350. doi: 10.1104/pp.99.1.348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makino A, Sakashita H, Hidema J, Mae T, Ojima K, Osmond B. Distinctive responses of ribulose-1,5-bisphosphate carboxylase and carbonic anhydrase in wheat leaves to nitrogen nutrition and their possible relationship to CO2 transfer resistance. Plant Physiol. 1992;100:1737–1743. doi: 10.1104/pp.100.4.1737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mills GA, Urey HC. The kinetics of isotopic exchange between carbon dioxide, bicarbonate ion, carbonate ion and water. J Am Chem Soc. 1940;62:1019–1026. [Google Scholar]
- Price D, von Caemmerer S, Evans JR, Yu JW, Lloyd J, Oja V, Kell P, Harrison K, Gallagher A, Badger M. Specific reduction of chloroplast carbonic anhydrase activity by antisense RNA in transgenic tobacco plants has a minor effect on photosynthetic CO2 assimilation. Planta. 1994;193:193–331. [Google Scholar]
- Roden JS, Ehleringer JR. Observations of hydrogen and oxygen isotopes in leaf water confirm the Craig-Gordon model under wide-ranging environmental conditions. Plant Physiol. 1999;120:1165–1173. doi: 10.1104/pp.120.4.1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roupsard O, Gross P, Dreyer E. Limitation of photosynthetic activity by CO2 availability in the chloroplasts of oak leaves from different species and during drought. Ann Sci Forest. 1996;53:243–254. [Google Scholar]
- Sasaki H, Hirose T, Watanabe Y, Ohsugi R. Carbonic anhydrase activity and CO2 transfer resistance in Zn-deficient rice leaves. Plant Physiol. 1998;118:929–934. doi: 10.1104/pp.118.3.929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scrimgeour CM. Measurement of plant and soil water isotope composition by direct equilibration methods. J Hydrol. 1995;172:261–274. [Google Scholar]
- Sokal RR, Rohlf FJ. Biometry. Ed 2. New York: W.H. Freeman; 1981. [Google Scholar]
- Syvertsen JP, Lloyd J, McConchie C, Kriedemann PE, Farquhar GD. On the relationship between leaf anatomy and CO2 diffusion through the mesophyll of hypostomatous leaves. Plant Cell Environ. 1995;18:149–157. [Google Scholar]
- von Caemmerer S, Evans JR. Determination of the average partial pressure of carbon dioxide in chloroplasts from leaves of several C3 plants. Aust J Plant Physiol. 1991;18:287–306. [Google Scholar]
- von Caemmerer S, Farquhar GD. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta. 1981;153:376–387. doi: 10.1007/BF00384257. [DOI] [PubMed] [Google Scholar]
- Wang XF, Yakir D. Temporal and spatial variations in the oxygen-18 content of leaf water in different plant species. Plant Cell Environ. 1995;18:1377–1385. [Google Scholar]
- Wang XF, Yakir D, Avishai M. Non-climatic variations in the oxygen isotopic compositions of plants. Glob Change Biol. 1998;4:835–849. [Google Scholar]
- Williams TG, Flanagan LB. Effect of changes of water content on photosynthesis, transpiration and discrimination against 13CO2 and C18O16O in Pleurozium and Sphagnum. Oecologia. 1996;108:38–46. doi: 10.1007/BF00333212. [DOI] [PubMed] [Google Scholar]
- Williams TG, Flanagan LB, Coleman JR. Photosynthetic gas exchange and discrimination against 13CO2 and C18O16O in tobacco plants modified by an antisense construct to have low chloroplastic carbonic anhydrase. Plant Physiol. 1996;112:319–326. doi: 10.1104/pp.112.1.319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yakir D. Oxygen-18 of leaf water: a crossroad for plant-associated isotopic signals. In: Griffiths H, editor. Stable Isotopes and the Integration of Biological, Ecological and Geochemical Cycles. Oxford: BIOS Scientific Publishers; 1998. pp. 147–168. [Google Scholar]
- Yakir D, Berry JA, Giles L, Osmond CB. Isotopic heterogeneity of water in transpiring leaves: identification of the component that controls the δ18O of atmospheric O2 and CO2. Plant Cell Environ. 1994;17:73–80. [Google Scholar]
- Yakir D, De Niro MJ, Rundel PW. Isotopic inhomogeneity of leaf water: evidence and implications for the use of isotopic signals transduced by plants. Geochim Cosmochim Acta. 1989;53:2769–2773. [Google Scholar]
- Yakir D, Wang XF. Estimation of CO2 and water fluxes between terrestrial vegetation and the atmosphere from isotope measurements. Nature. 1996;380:515–517. [Google Scholar]