
Extended data Figure 6. Ascorbate regulates HSC function in vivo and HSC differentiation in culture.
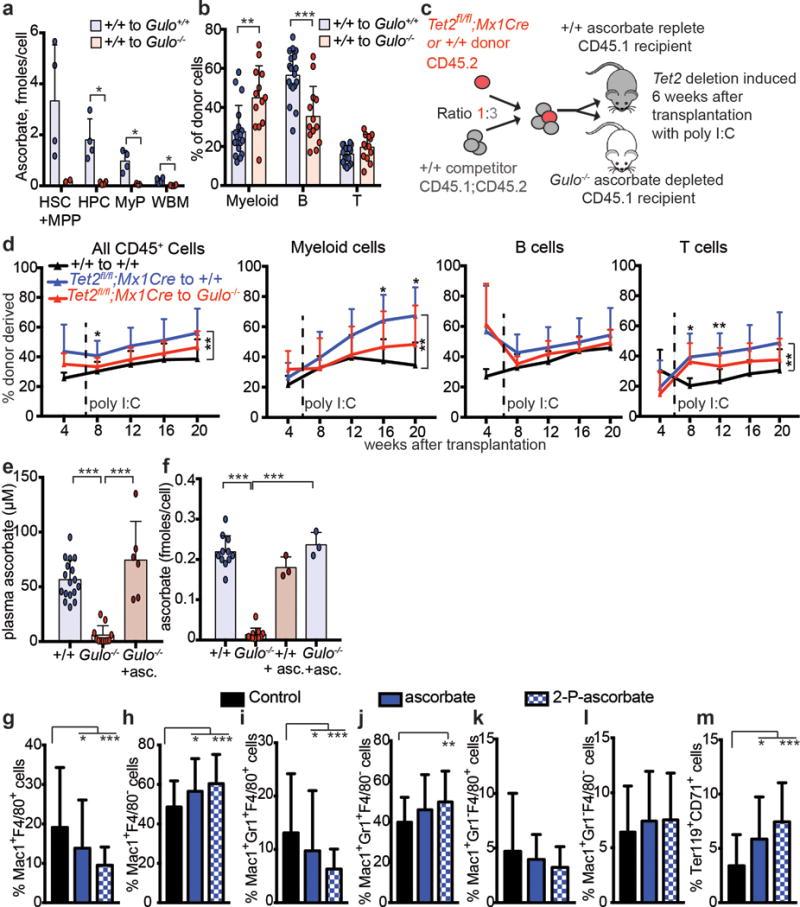
a, Ascorbate levels in sorted cell populations from transplanted recipient mice (a total of n=2-4 mice per genotype from 2 independent experiments). b, Percentage of donor cells that are myeloid, B, or T cells in wild type or Gulo−/− recipients 16 weeks after transplantation (a total of n=13-18 mice per condition analysed in 4 independent experiments). c-d, Competitive transplantation of 500,000 donor bone marrow cells from Tet2fl/fl;Mx1Cre mice or littermate controls along with 1,500,000 competitor wild-type cells into irradiated wild-type (ascorbate replete) or Gulo−/− (ascorbate depleted) mice (a total of 2 donor mice and 5–10 recipient mice per treatment in 2 independent experiments). e, Plasma ascorbate levels in wild type mice, Gulo−/− mice, or Gulo−/− mice fed an ascorbate supplemented diet (a total of n=6-17 mice per condition analysed in 4 independent experiments). f, Ascorbate content in bone marrow cells from wild type and Gulo−/− mice fed normal mouse chow or an ascorbate supplemented diet (a total of n=3-11 mice per condition analysed in 3 independent experiments). g-m, Myeloid and erythroid differentiation were assessed 8 days after culturing HSCs in the presence or absence of ascorbate or its more stable derivative, 2-phospho-ascorbate (a total of n=48 wells for myeloid and 24 wells for erythroid differentiation in 2 independent experiments). All data represent mean±SD. Statistical significance was assessed with Welch’s tests (a-b), Kruskal-Wallis tests (e-m), or a non-parametric mixed model followed by a Kruskal-Wallis test for individual time points (d). We corrected for multiple comparisons by controlling the false discovery rate (*p<0.05, **p<0.01, ***p<0.001).