

Abstract
We studied inherent variation in final leaf size among four Poa spp. that live at different elevations. The average final length of leaf 7 of the main stem of the smallest species (Poa alpina) was only one-half that of the largest species (Poa trivialis); it was correlated with leaf elongation rate, but not with the duration of leaf elongation. A faster rate of leaf elongation rate was associated with (a) larger size of the zone of cell expansion, and (b) faster rates of cell production (per cell file) in the meristem, which in turn were due to greater numbers of dividing cells, whereas average cell division rates were very similar for all species (except Poa annua). Also we found that the proliferative fraction equaled 1 throughout the meristem in all species. It was remarkable that rates of cell expansion tended to be somewhat higher in the species with slower growing leaves. We discuss the results by comparing the spatial and material viewpoints, which lead to different interpretations of the role of cell division. Although the presented data do not strictly prove it, they strongly suggest a regulatory role for cell division in determining differences in growth rate among the present four Poa spp.
The size of mature leaves varies widely among plant species and is the result of rate and duration of leaf expansion. In grass leaves, growth predominantly occurs along the longitudinal axis and the steady-state leaf elongation rate is commonly calculated from a linear increase of leaf length with time (Bultynck et al., 1999, and refs. therein). In forage grasses, leaf elongation rate has been positively associated with yield per tiller and rate of canopy development (Horst et al., 1978). In a previous paper we presented a framework for the analysis of the relationship between leaf elongation rate of individual leaves and shoot relative growth rate using contrasting Poa and Aegilops spp. as models (Bultynck et al., 1999). However, little is known about the cellular basis of intra- and interspecific variation that determines leaf elongation rate differences under favorable growth conditions. The relationship between cell division and expansion, and whole organ growth rate in general can be fruitfully studied in organs predominantly growing with a one-dimensional axis, such as roots and grass leaves. Such organs have a consistent spatial and temporal organization of the two activities, which leads to a typical cell length distribution (Green, 1976). The growth zone of monocotyledonous leaves is composed of a “division” zone, and an “elongation-only” zone. In the basal (intercalary) meristem, cells divide and expand simultaneously. The cells are, therefore, displaced to a more distal location by the ongoing formation of new cells in the basal portion of the meristem. The entrance in the elongation-only zone is characterized by the loss of the cells' ability to divide; in this region cells only expand, eventually reaching their mature size. It follows that a developmental gradient is present along the growing axis and that the spatial axis along the organ constitutes an age distribution (Silk et al., 1989). Outside the growing region, tissue maturation takes place.
There are two alternative ways of conceptually looking at how the organ growth rates are determined (for review, see Silk and Erickson, 1979; Silk, 1984). First, there is the spatial (Eulerian) view, which describes the distribution of expansion and division rates along the axis of the elongating organ. From this perspective, organ growth rates are a function of the integrated relative elongation (or strain) rates over the whole of the growth zone. This view strongly suggests that position and size of the zone where cells expand are determined by positional control mechanisms, specifically acting on the process of cell expansion. Cell division in this view is merely a process that accompanies cell expansion in a limited region of the growth zone. In this view it is unclear how meristematic activity could exert an effect on organ growth rate. Second is the material (Lagrangian) view. It focuses on the development of individual cells as they move through the growth zone and proceed through their developmental program. Following this program, cells first replicate themselves a number of times (division and expansion) and subsequently expand to a certain mature size in the absence of division. In this view, the observed spatial distribution of strain rates is a consequence of the developmental program of component cells. In this perspective, the role of meristematic activity is fundamentally different. In case the expansion of individual cells in the elongation zone follows the same program (duration and rate), the rate of cell production in the meristem determines how many cells are elongating at any time, and thereby effectively controls the spatial distribution of strain rates along a growing organ. This proposition does not suggest that there can be growth by cell division (Green, 1976), and we explicitly refer to “meristematic activity” and “cell production,” which encompasses both cell division and expansion.
Different lines of evidence support a role of meristematic activity in controlling organ growth rates. Differences in elongation rates of leaves due to nitrogen supply (Volenec and Nelson, 1983; MacAdam et al., 1989), temperature (Ben-Haj-Salah and Tardieu, 1995), and between high- and low-yielding Festuca arundinacea genotypes (Volenec and Nelson, 1981) were associated with limited or no changes in mature cell size, suggesting a correlation between leaf elongation rate and cell production rate. Beemster and Baskin (1998) recently showed that the developmental acceleration of root growth of Arabidopsis was associated with a nearly proportional increase in the rate of cell production, which was associated with a progressively larger number of cells dividing at an approximately constant rate through time. Moreover, in the elongation zone cells expanded at approximately the same rates and for approximately the same duration in these roots. In contrast, high soil mechanical impedance decreased mature cell length only, or both cell production and mature cell length depending on leaf position on the main stem (Beemster et al., 1996), showing that there are multiple possibilities of altering organ growth rate even in a given plant species and in response to the same treatment.
In the present work we investigated if the observed inherent variation in leaf elongation rate among four species of the genus Poa that grow at different elevations could be due to differences in meristematic activity. The analysis of epidermal cell dynamics in the growth zone of leaf 7 of the main stem showed that according to the spatial view the extent of the growth zone, in particular that of the elongation zone, is the major determinant of leaf elongation rate. However, when adopting a material view, differences in cell production could account for variations in the size of the elongation zone and thereby for variation in leaf growth rate among these species.
RESULTS
Variation in Mature Leaf Length, Leaf Elongation Rate, and Duration of Leaf Elongation
The average final length of leaf 7 differed more than 2-fold between the species with the longest (Poa trivialis) and the species with the shortest (Poa alpina) leaves (Table I). Poa annua and Poa compressa showed an intermediate final leaf size. Final leaf length can be viewed as a function of leaf elongation rate and the duration of leaf elongation. For all the species, the increase in leaf length with time was virtually steady for at least 3 d, from d 2 to 5 from leaf appearance (Fig. 1). The approximately steady leaf elongation rate during most of the duration of leaf expansion is in accordance with earlier studies (Gallagher, 1979; Schnyder et al., 1990). Presuming the same holds for Poa spp., these features allow the calculation of leaf elongation rate as the slope of a regression line within the period of steady growth (see “Materials and Methods”) for each analyzed leaf. Differences in leaf size were not correlated with the duration of leaf elongation, but were closely matched by differences in leaf elongation rate (Table I). Leaf elongation rates of this experiment were nearly identical to those measured in two earlier experiments under the same environmental conditions of this study (a subset of these data was described by Bultynck et al. [1999]). Moreover, in a field analysis proportional differences in leaf elongation rates were found among P. annua, P. trivialis, and P. alpina (Körner and Woodward, 1987). In these earlier experiments, we also showed that the variation in leaf elongation rate among these four Poa spp. was very similar in magnitude for each leaf position ranging from leaves 3 to 7 of the main stem (Bultynck et al., 1999). Hence, the following analysis of the cellular basis for differences in leaf elongation rate of leaf 7 was probably representative for other leaf positions.
Table I.
Final leaf length, leaf elongation rate, and duration of leaf elongation of leaf 7 of the main stem for four Poa spp.
| Species | Final Leaf Length | Leaf Elongation Rate | Duration of Leaf Elongation |
|---|---|---|---|
| mm | mm d−1 | d | |
| Poa alpina | 41 ± 2a | 6.5 ± 1.1a | 5.9 ± 0.2ab |
| Poa compressa | 81 ± 4bc | 12.0 ± 1.4b | 6.9 ± 0.3b |
| Poa annua | 69 ± 4b | 13.4 ± 1.6bc | 5.3 ± 0.2a |
| Poa trivialis | 98 ± 5c | 16.1 ± 2.3c | 5.8 ± 0.2ab |
Results are the average of 15 plants ± se. The letters represent statistically homogenous subgroups (Tukey b post hoc test at α = 0.01 significance level). The parameters were calculated as described in “Materials and Methods.”
Figure 1.

Increase in length of the blade of main stem leaf 7 as a function of time from leaf appearance for four Poa spp. Symbols indicate means ± se in case this is larger than the symbols (n = 15). In the inset, leaf elongation rate (LER) during d 2 to 5 from leaf appearance is shown. Symbols are the same as in the main panel.
Spatial Characterization of the Growth Zone: Cell Length and Strain Rate Profiles
In Lolium perenne leaves, epidermal cell length profiles were constant during the entire period of approximately linear growth (Schnyder et al., 1990). It is generally presumed this characteristic holds true for other grasses and this facilitates the use of cell length profiles as a basis for the kinematic analysis. To investigate the cellular basis of differences in leaf elongation rate, we first analyzed the relationship between leaf elongation rate and spatial distribution of cell division and expansion activity. To accomplish this, we determined, in addition to leaf elongation rate, the length of epidermal cells in files adjacent to the stomatal files as a function of position along the leaf axis (Fig. 2A). In all the species, the most basal cells in the division zone showed a highly conserved average length of about 15 μm. This means that the comparison of the profiles at further locations along the growth zone was not biased by a different initial size of meristematic cells. Moving distally from the leaf base, cells increased their length more sharply in the two subalpine species (P. alpina and P. compressa) compared with the two lowland ones (Fig. 2A). Mature cell length was very similar in P. annua (184 μm), P. compressa (199 μm), and P. trivialis (182 μm), whereas in P. alpina it was significantly lower (152 μm; lsd = 18 μm at α = 0.01). Therefore, with the exception of P. alpina, differences in cell expansion over the growth zone as a whole appear to be nearly completely mirrored by differences in cell division activity.
Figure 2.

Profiles of cell length (A) and strain rate (B) of files adjacent to stomatal files in the adaxial epidermis of main stem leaf 7 of four Poa spp. Data are averages ± se of five leaves per species. For each leaf, all cells in the basal 10 or 20 mm (depending on the species) of five files adjacent to stomatal files were measured and the data were subsequently smoothed and interpolated (see Fig. 5). Strain rate was calculated on the basis of cell length profiles as described in “Materials and Methods” and each curve is plotted outside the meristematic region, according to the estimated length of the meristem. The average maximal strain rates were 8.8%, 10.5%, 7.3%, and 7.6% h−1 for P. alpina, P. compressa, P. annua, and P. trivialis, respectively (lsd = 2% h−1 at α = 0.01).
The strain rate profiles, representing local rates of cell expansion, could be derived from the cell length profile and leaf elongation rate (see Eqs. 6 and 11). They showed a roughly bell shaped distribution with the maximum in the middle (Fig. 2B). It is interesting that the length of the growth zone reflected the differences in leaf elongation rate (Fig. 3, A and B). In contrast, the two species with the lowest leaf elongation rate had the highest maximal strain rates, which partly compensated for the shorter extent of the growth zone. It follows that no relationship was found between maximal strain rate and leaf elongation rate. Particularly the comparison between P. compressa and P. annua is striking, the first species having a roughly 30% shorter growth zone, which was almost entirely compensated for by a nearly equal difference in height of the strain rate profile (see leaf elongation rate in Table I). It seems that the length of the growth zone was the primary determinant of variation in leaf elongation rate between these species. The inference that cell division and cell expansion covary in these species was reflected in the portion of the entire growth zone where cells divide, which ranged between 15% and 18%. P. annua constituted an exception in that the meristem spanned only approximately 10% of the growth zone (Fig. 3A).
Figure 3.

Characterization of the growth zone of main stem leaf 7 of four Poa spp. The stacked bars represent length (A), total number of cells (B), and cell residence time (C) in the growth zone. The species are arranged according to ascending average leaf elongation rate and the values are shown at the top of A. In all panels, the white bars refer to the division zone and the black bars to the elongation-only zone. Data are averages ± se of five leaves per species and of five equivalent cell files adjacent to stomatal files for each leaf (adaxial epidermis). Details of the calculations are given in “Materials and Methods.” In all panels, the vertical black lines to the left represent lsd (α = 0.01) for the division zone (lsddiv), for the elongation-only zone (lsdez), and for the growth zone as a whole (lsdgz). LER, Leaf elongation rate.
Material Characterization of the Growth Zone
The main question to be addressed in this contribution is if differences in leaf elongation rate could be due to differences in meristematic activity. To address this question, the material perspective is particularly appropriate, as it integrates the development of individual cells that are produced by meristematic activity.
We first determined the number of cells in the meristem and in the elongation-only zone (Fig. 3B). Due to the fact that meristematic cells were much smaller than cells in the elongation-only zone, meristematic cells represented approximately one-half of the cells that were present in the entire growth zone, despite the fact that they occupied only approximately 15% of its total length (Fig. 3, compare A and B). An exception is P. annua, where the relatively short meristem contained only about one-third of the total number of growing cells. Despite this finding, there was a clear and positive correlation between leaf elongation rate and the number of cells in both the meristem and elongation-only zone (Fig. 3B).
The residence time in the meristem accounted for 70% to 90% of the total time cells grow (residence time in the growth zone; Fig. 3C). Although less pronounced than the length of the growth zone and the number of cells in the growth zone, the residence time in the growth zone was also positively correlated to leaf elongation rate, due to variations in the duration of the meristematic phase (Fig. 3C). In fact, the residence time in the elongation-only zone was generally very similar in all the species (Fig. 3C). An exception was formed by P. annua, whose cells divided for a significantly shorter time compared with the other three species, which was partly compensated for by a longer residence time in the elongation-only zone.
Considering all the species, differences in leaf elongation rate were not correlated with the expansion of individual cells in the elongation-only zone. Instead, the larger size of the elongation-only zone in faster growing species was closely correlated with the number of cells in it (Fig. 3, A and B). An exception was P. alpina, due to the smaller cell size in this species (Fig. 2). Therefore, the elongation zone was much shorter than that in P. compressa, even though it contained approximately the same number of cells. Given that each cell expanded for approximately the same time (Fig. 3), variations in the number of elongating cells must be due to differences in the rate at which cells were produced by the meristem. Thus, it seems plausible that differences in meristematic cell production rate determined the number of elongating cells, which in turn determined the size of the elongation zone.
Cell Production and Cell Division Rates in the Meristem
In accord with a role of meristematic cell production in determining leaf elongation rate, the number of cells produced per unit of time was strongly correlated with leaf elongation rate (Fig. 4A). Differences in meristematic cell production were not due to differences in average rate of cell division, which was not significantly different among the species, P. annua excepted (Fig. 4B). Instead, differences in cell production among P. alpina, P. compressa, and P. trivialis were associated with variations in the number of cells in the division zone, which was reflected both in a larger length of the division zone and in a longer cell residence time in this region (Fig. 3). In contrast to this general trend, P. annua showed a relatively high cell production due to a significantly shorter cell cycle time of 14 h compared with 18, 20, and 21 h for P. alpina, P. compressa, and P. trivialis, respectively (lsd = 3 h at α = 0.01). Accordingly, the faster rate of cell division of P. annua was offset by a smaller number of dividing cells which remained in the division zone for a shorter time (Fig. 3C). This explains the relatively small meristem size observed in this species (Fig. 3A).
Figure 4.

Cell production (A) and average rate of cell division (B) of main stem leaf 7 of four Poa spp. The species are arranged according to ascending average leaf elongation rate and the values are shown at the top of A. Data are averages ± se of five leaves per species and of five equivalent cell files adjacent to stomatal files for each leaf (adaxial epidermis). The vertical black lines to the left represent lsd for the two parameters (α = 0.01). LER, Leaf elongation rate.
Clowes (1976) and, more recently, Francis (1998) suggested that differences in cell production could occur because a varying fraction of the total population of meristematic cells divide continuously before being displaced to the elongation-only zone (proliferative fraction). We analyzed our cell length data to determine if the proliferative fraction could be smaller than 1 in any of these species. If a subpopulation of cells stops dividing early it will continue to expand due to the inability of cells to slide relative to each other. This will increase the size range of the total population with increasing distance from the base of the meristem (Green and Bauer, 1977; Webster and MacLeod, 1980). For each species, we calculated the proportion of cells lying outside a minimum 2-fold size range (cells divide in half) located in 100-μm intervals at 25%, 50%, and 75% of the length of the meristem, and we compared them with the proportions obtained for cells lying in 1-mm intervals in the mature region. The length of the different intervals was chosen to obtain distributions including approximately the same number of cells. For all of the species, the cell length distribution range was the same throughout the meristem and in the mature region (Table II). This result indicates that, when this specific cell type is considered, no cells stopped dividing early (proliferative fraction equaled 1 throughout the meristem in all species).
Table II.
Comparison of cell length distribution in the meristem and in the mature region (leaf 7) of four Poa spp.
| Species | Cells with Length Outside a 2-Fold Size Range
|
|||
|---|---|---|---|---|
| 25% Meristem length | 50% Meristem length | 75% Meristem length | Mature region | |
| % | ||||
| P. alpina | 17 ± 3 | 24 ± 3 | 21 ± 4 | 20 ± 5 |
| P. compressa | 17 ± 5 | 18 ± 4 | 17 ± 3 | 16 ± 3 |
| P. annua | 15 ± 2 | 16 ± 3 | 20 ± 3 | 12 ± 3 |
| P. trivialis | 15 ± 3 | 11 ± 2 | 13 ± 2 | 10 ± 2 |
Cell length distributions were analyzed for each leaf (five combined epidermal cell files) in 100-μm intervals centered at 25%, 50%, and 75% of the estimated length of the meristem and in 1-mm intervals in the mature region. Data were averaged between leaves (n = 5) and mean ± se are presented. The values for each interval in the meristem were compared with those of the mature region with paired t test. In all of the cases the two-tailed Student's t test probability was not significant (P > 0.05). The proportion of cells with length lying outside a 2-fold range was calculated expressing the data as percentiles and repeatedly eliminating equal percentiles starting from the lowest and highest limit of the distribution, until the desired range was reached. The total number of cells in the intervals at 25%, 50%, and 75% of the length of the meristem and in the mature region were, respectively, 151, 129, 112, and 167 (P. alpina); 113, 88, 95, and 102 (P. compressa); 126, 105, 103, and 107 (P. annua); and 117, 113, 94, and 111 (P. trivialis).
DISCUSSION
Final Leaf Size, Leaf Elongation Rate, and Duration of Leaf Elongation
In this study we show that variation in final leaf size among the four Poa spp. is mainly due to differences in leaf elongation rate and not to differences in duration of leaf expansion. A similar conclusion was drawn from field observations on populations of P. annua, P. trivialis, and P. alpina that live at different elevations (Körner and Woodward, 1987).
The duration of leaf elongation calculated in this work refers to the visible period of leaf extension, from emergence of the tip from the leaf sheath of the previous leaf to the end of the growth period (apparent duration). As a consequence, architectural characteristics of the examined species could affect the estimation of this parameter. In particular, differences in the proportion of leaf length that is encircled by the sheath of the previous leaf could result in leaves of different developmental stages being studied. However, we find that the portion of leaf 7 that is enclosed by leaf 6 is approximately similar for these four species (14%–17%) at maturity. Note also that in this study no distinction between sheath and blade growth has been made. Again, the portion of total leaf length that constitutes the blade of leaf 7 is approximately the same in these species, ranging from 15% to 18%. Moreover, the work of Schnyder et al. (1990) shows that except for the position of the ligule that separates the blade and the sheath domains, the cell length profile in grasses is not affected by the switch between leaf blade and sheath growth and can therefore be analyzed as a continuum.
Although the period of concealed growth from leaf initiation to leaf emergence accounts for only a minimal fraction of the final leaf size, it possibly plays an important role in setting up differences in leaf growth zone characteristics (Beemster and Masle, 1996). Therefore, developmental differences occurring during this phase and their relationship to parameters such as length and width of the meristem during the period of linear growth after tip emergence need to be addressed in future research.
Can Cell Division Determine Leaf Elongation Rate?
Here we analyzed the cellular basis of differences in leaf elongation rate among four Poa spp. From a spatial viewpoint, leaf elongation rate is determined solely by the integration of strain rate (cell expansion) over the length of the growth zone. Our data for the Poa spp. show that leaf elongation rate was primarily determined by the size of the growth zone. Cell division, according to this viewpoint, is merely an accompanying process with no direct relationship to growth, i.e. to a given increase in volume. However, when analyzed from a material viewpoint, we showed that differences in the size of the growth zone were correlated directly to the number of cells produced in the meristem per unit of time. Cell production in turn varied mainly due to differences in the number of dividing cells rather than to differences in division rate. These results are analogous to the comparison of Arabidopsis roots of different age (Beemster and Baskin, 1998) and confirm the hypothesis that meristematic activity could determine leaf elongation rate.
This hypothesis is formally presented in the model proposed by Ivanov and Dubrovsky (1997) that directly links whole organ growth rate to the number of dividing cells and average cell cycle duration:
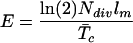 |
1 |
Note that this model is based on entirely different parameters as the spatial model represented by the following equation, which views organ elongation rate as a function of the strain rate (r[x]) only:
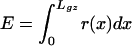 |
2 |
Specifically, E (representing organ elongation rate) in Equation 1 is related to average cell cycle duration and the number of dividing cells, which are cell division parameters. The suggestion that cell division could determine organ growth rates contrasts with the spatial model, where cell division only accompanies cell expansion and its role is limited to ascertain appropriate partitioning of the created volume and where the correlation between leaf elongation rate and division activity is a consequence rather than a cause. Based on the current study, it is impossible to prove which view accurately represents the underlying regulatory mechanism. To address this matter, various laboratories are currently studying the effect on cell and organ development of a selective inhibition or stimulation of the process of cell division in Arabidopsis roots. This is accomplished by analyzing the growth of plants in which the expression level of specific cyclins and cell cycle regulatory genes is altered (for review, see Jacobs, 1995; Mironov et al., 1999). Provisional evidence on the overexpression of the mitotic cyclin1At under the promotor of cdc2aAt, which resulted in increased growth rates of Arabidopsis seedlings not accompanied by increased mature cell length (Doerner et al., 1996), supports the material view and a key role for cell division in determining organ growth rates.
In addition to what we discussed above, the exception represented by the annual species P. annua (short meristem, but high cell division rate, which results in a high cell production) shows that a given organ growth rate can be obtained in various ways. In fact, we observed that P. annua and P. compressa were characterized by a similar leaf elongation rate and length of mature cells and by a similar cell production of leaf 7. However, the same mature cell length was realized in P. compressa during a short residence time in the elongation-only zone and a high maximal strain rate and in P. annua during a long residence time in the elongation-only zone and a low maximal strain rate. Likewise, the two species achieved the same rate of cell production by different means, either a high cell division rate and a small number of dividing cells (P. annua) or a large number of dividing cells and a slow cell division rate (P. compressa). The contrast between P. compressa and P. annua in this study additionally implies that different sorts of coordination between cell division and cell expansion may exist in different species. However, based on the comparison of all four Poa spp., we conclude that faster growing leaves are generally characterized by a higher cell production (per each equivalent epidermal cell file), principally caused by a larger size of the meristem.
Inherent Variation in Leaf Elongation Rate and Rate of Cell Division
A relatively high rate of leaf growth in this study was not related to the average rate of cell division or to the corresponding cell cycle time. On these results, two distinct comments can be made.
First, Francis (1998) recently suggested that modeling a population of meristematic cells could obscure the (putative) functional role of specific subpopulations of cells that stop dividing early within the meristems. The analysis and the comparison of cell length distribution at different locations of the leaves' growth zone region in the present study (see “Results”; Table II) reveal that cells of epidermal files continue to divide throughout the whole length of the meristem; the proliferative fraction was consequently equal to 1 throughout the meristem in all four species. Moreover, throughout the meristem we found rather low percentages of cells lying outside a 2-fold size range, indicating a fairly tight regulation of cell size at cytokinesis. To our knowledge, such comparative results, derived from cell length measurements of a specific file type, have not been presented before for leaf meristems of different species. It is interesting that our results are strikingly similar to those found by Beemster and Baskin (1998) for epidermal cells of the primary root of Arabidopsis, suggesting that a tight regulation of the size at which cells divide is common for most organs and for a wide range of species.
Second, P. annua had a 35% shorter cell cycle time compared with the other three species. A factor that may account for this difference is that P. annua is the only annual species in this comparison (Hegi and Conert, 1998). The question arises whether there is a relationship between rate of cell division and generation time, genetic (genome size, ploidy level), cytological (cell size), and nuclear features (nuclear volume). The present four species have a different ploidy level: P. annua is a tetraploid and P. trivialis is a diploid, whereas P. alpina and P. compressa are characterized by a mixture of ploidy levels ranging from diploid to tetraploid depending on the population (Hegi and Conert, 1998). The amounts of nDNA in the unreplicated haploid genome of a gamete (i.e. DNA-C value) of P. annua and P. trivialis are nearly identical, being 4C = 11.50 pg and 4C = 11.30 pg, respectively (Bennett and Smith, 1976). Based on these data, genome size is probably not the cause of the observed differences in rate of cell division in these two species. We can also remark that mature cell length is nearly identical in these two species and, therefore, mirrors DNA-C values. For P. alpina and P. compressa no data are available in current angiosperm DNA-C value databases (Bennett et al., 1998), and therefore no further conclusion is possible on this point. However, the potential importance of DNA-C value for determining cell size and growth rate was shown by Mowforth and Grime (1989). They reported a variation of about 80% in DNA-C value among seed progeny of families derived from a single pasture population of P. annua, which was positively correlated with cell size and seedling growth rate. Finally, a positive correlation has been found among minimum mitotic cell cycle time, duration of meiosis, and DNA-C value (Van't Hof and Sparrow, 1963; Bennett, 1977). The difference in cell cycle duration between P. annua and P. trivialis we observed, combined with their identical DNA-C values, seems to contradict this proposition. However, based on such considerations and with the aid of kinematic methods and cytometric analyses to estimate DNA-C values (Doležel et al., 1989), more detailed research can be done to clarify this relationship by means of intra- and interspecific comparisons of appropriate ranges of species (e.g. genotypes differing in ploidy level and genome size). Because of their relative abundance and variety in life form, Poa spp. are good objects for this kind of research.
From Individual Leaf Growth to Whole Shoot Growth
A final consideration pertains to the relevance of our findings in the perspective of whole plant growth. We investigated the cellular basis of genotypic variation in leaf size and leaf elongation rate at one developmental stage. Is the size of the meristem also a key factor that explains variation in leaf size and leaf elongation rate of grasses through a longer period of vegetative development, i.e. for subsequent leaves of the main stem and for subsequent tillers? Beemster et al. (1996) found a positive correlation between leaf elongation rate of subsequent leaves of the main stem of wheat (Triticum aestivum) and length of the division zone. The development of tillers can reasonably be assumed to be the result of modular growth, i.e. the repetition in space and time of the same fundamental unit constituted by a meristem and elongating internodes, and therefore results obtained for the main stem might also apply to tillers. However, when expanding our perspective to the whole shoot level, no obvious relationship is expected between leaf elongation rate and shoot relative growth rate, as described by Bultynck et al. (1999). In fact, steady-state leaf elongation rate of individual leaves (a linear variable) cannot quantitatively explain variation in relative growth rate (an exponential variable). Cell division and cell expansion are intrinsically exponential processes (Lockhart, 1965; Green, 1976), but they can only theoretically account for variation in relative growth rate when they are not viewed on a per organ basis. Therefore, the links between cell division, cell expansion, cell production rates, and meristem size on one hand and whole shoot and entire plant growth rate on the other still largely remain to be explored.
MATERIALS AND METHODS
Plant Material and Growing Conditions
Seeds of the lowland species Poa annua and Poa trivialis were purchased commercially (Kieft, Blokker, The Netherlands). Seeds of the subalpine Poa compressa were harvested from a natural population at 1,600 m above sea level and were kindly provided by Orto Botanico di Genova (Italy). Seeds of Poa alpina (1,800 m above sea level) were kindly provided by Dr. O.K. Atkin (Department of Biology, University of York, UK) and were obtained as described previously by Atkin et al. (1996). To avoid the comparison of species exhibiting a different leaf morphotype (flat-leafed versus rolled-up leaves, the latter being more frequent in subalpine and alpine species), only flat-leafed species were selected (Atkin et al., 1996; Van Arendonk et al., 1997). However, P. annua is an annual species, whereas the other three are perennials. Seeds of all of the species were germinated in the dark at 20°C ± 1°C for 2 to 3 d in Petri dishes on a double layer of filter paper wetted with demineralized water, until the primary root had protruded and the coleoptile had emerged. After this period, the seedlings were placed in growth chambers at 20°C ± 1°C with 70% relative humidity, photosynthetic photon flux density of 450 ± 20 μmol m−2 s−1, and a 14/10 h day/night cycle. When the primary root was at least 2 cm long, the seedlings were placed in sand for 1 week and watered daily with a one-tenth-strength modified Hoagland solution (Poorter and Remkes, 1990) to allow the further development of roots and the expansion of leaf 2. The roots were quickly rinsed with demineralized water, and plants were then transferred to an aerated full-strength modified Hoagland solution in 33-L containers (Poorter and Remkes, 1990). In each container, 24 plants of a given species were grown, and there were a total of three containers per species. The pH was adjusted to 5.8 every 2nd d and the solution was renewed weekly.
Measurement of Leaf Elongation
For all the species, the increase in length of leaf 7 with time was measured twice a day, after the start of the light period and before the beginning of the dark period, using a ruler and taking the ligule of the preceding leaf as a reference point. Since there is no significant growth of the sheath of leaf 6 after the appearance of leaf 7, this procedure allowed an accurate estimate of leaf elongation rate. The measurements were carried out on 15 plants per species throughout the whole period of blade elongation, from leaf emergence to its complete extension.
The increase of leaf length with time was linear for at least 3 d in all the species. Leaf elongation rate of individual leaves was calculated as the slope of the regression line fitted through the data points in the interval 15% to 95% of the final leaf length. Before this interval (1st d after leaf appearance) the increase in leaf length with time was not steady, and after this interval it declined sharply, marking the end of the elongation period. Data were averaged between leaves and mean leaf elongation rate was used for subsequent calculations, as described below. Duration of leaf elongation was determined for each individual leaf as the time interval between leaf appearance and the time point at which leaves reached 95% of their final length. Mean duration of leaf elongation was obtained by averaging data between leaves.
Measurement of Cell Length
During the 2nd d of steady growth of leaf 7, five plants per species were harvested from the same batch of plants that was used for determination of leaf elongation rate, mature leaf size, and duration of leaf elongation, and dissected under a binocular microscope to expose the growth zone of leaf 7. Samples of leaf 7 were immediately placed in boiling methanol for 15 to 20 min, for chlorophyll removal and fixation. Finally, they were placed in 90% (w/v) lactic acid (Merck, Darmstadt, Germany) for clearing and storage. The length of the leaves was measured before and after boiling and no significant tissue shrinkage was observed.
Moving distally from the leaf base, cell length was measured along five equivalent cell files adjacent to stomatal rows in the adaxial epidermis of each leaf. The measurements were made using image analysis on bright field light microscopical images. The setup consisted of a light microscope (fitted with Plan 6.3× and Neofluar 10× objectives, Kontron/Zeiss, Eching, Germany) equipped with a black and white CCD camera type WC-CD50 (frame size, 768 × 512 pixels, 256 gray levels, Panasonic, Tokyo). The image was acquired by a computer and displayed on screen. The image contours were enhanced to highlight cell walls, and the lengths of individual cells were measured by tracing straight lines between successive transverse cell walls with a pointer device. The length of the individual segments and cumulative length were stored in a data file. This procedure was repeated for partially overlapping image fields covering the extent of the growth zone and part of the mature region. This measurement routine was implemented as a customized program developed by Dr. M. Terlou (Image Processing and Design, Faculty of Biology, Utrecht University, The Netherlands). Cell length distribution along the leaf axis was obtained by plotting the length of each cell as a function of the position of its midpoint relative to the leaf base. The data for all files were combined for each leaf and then smoothed and interpolated into 25-μm equally spaced points by using a kernel smoothing method described earlier (Beemster and Baskin, 1998). This enabled averaging among leaves of each species to obtain the cell length profiles. The interpolation algorithm was implemented as a macro for Microsoft Excel using Microsoft Visual Basic 5 for Applications (Microsoft, Redmond, WA). A representative fit is shown in Figure 5. This method, despite the lack of obvious biological meaning of parameters derived by each local polynomial fit, has the advantage of making the least assumptions about the shape of the curve when compared, for instance, to appropriate logistic functions. For a discussion about the use of logistic models for the description of leaf growth and cell length data, see Richards (1959) and Venus and Causton (1979).
Figure 5.

Cell length distribution along the blade of a representative leaf of P. alpina (adaxial epidermis; leaf 7). The solid line represents the smoothed and interpolated data at intervals of 25 μm using the kernel smoothing algorithm described by Beemster and Baskin (1998). Cell length data were obtained from five equivalent cell files adjacent to stomatal rows and plotted against the position of their midpoint.
Observing the smoothed cell length profile of each leaf, the lengths of mature cells were calculated by averaging all data points distal to the position where the increase of cell length between successive points was ≤0 μm. The length of the growth zone (Lgz) was estimated as the distance from the leaf base to the position at which cells reached 95% of their mature length. These values were then averaged between leaves and used for subsequent calculations.
Estimation of the Length of the Meristem
An additional subset of five plants per species was used to estimate the length of the division zone. Plants were harvested and dissected to expose the growth zone of leaf 7 during the 2nd d of linear growth. Leaf tissue was fixed and chlorophyll was removed in 3:1 (v/v) absolute ethanol:glacial acetic acid for at least 24 h at 4°C. The samples were then hydrolyzed in 5 m HCl for 20 to 30 min and rinsed with a solution of 0.1 m Na2S2O5 dissolved in 0.15 m HCl. Nuclei were visualized by staining leaves for at least 2 h with a Feulgen dye prepared with basic fuchsin (pararosaniline; Janssen Chimica, Beerse, Belgium). Tissue was subsequently mounted on an object slide in the same Na2S2O5 solution and covered with a coverslip. The same image analysis system utilized for cell length measurements was used for observing mitotic figures starting from the leaf base and proceeding distally along adaxial epidermal cell files adjacent to stomatal rows. In these cell files only symmetrical (“proliferative”) divisions took place (Larkin et al., 1997; Croxdale, 1998), yielding daughter cells of approximately equal size. Similarly to the approach used by Barlow et al. (1991) for tomato (Lycopersicon esculentum) roots, the length of the division zone (Ldiv) was estimated for this cell type by recording the distance from the leaf base to the most distal mitosis. For each leaf, at least 10 cell files were observed and the most distal mitosis in any of these files was taken as the distal margin of the meristem. Coincidentally, we noticed that in the adjacent stomatal files a shift from proliferative divisions to asymmetrical (“formative”) divisions, yielding stomatal initial cells, occurred at the same distance from the leaf base.
Numerical Analysis
Based on the measurements of leaf elongation rate and cell length distribution, we performed a kinematic analysis of leaf elongation, founded predominantly on the theoretical works of Erickson and Sax (1956), Green (1976), Gandar (1980) and Silk and Erickson (1979). The position of each interpolated cell length data point as a function of the distance from the leaf base was defined as X = n × Δx, where n = 1,2,3,… and Δx is a step of 25 μm, and was obtained from the kernel smoothing procedure as described above. The number of cells per file in the growth zone (Ngz) and in the division zone (Ndiv) was calculated from local cell density, ρ(x), which is the reciprocal of local cell length, l(x), with the following formula:
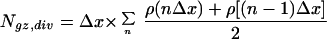 |
3 |
where the summation was stopped at the distal margin of the growth zone and of the division zone, respectively.
The length (Lez) and number (Nez) of cells of the elongation-only zone were determined as:
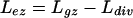 |
4 |
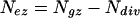 |
5 |
respectively, where Lgz and Ldiv were obtained as described above.
Cells that divide in the division zone are displaced to a more distal location by ongoing cell production in the more basal part of the meristem. Per cell file, the rate of cell production in the meristem at the base of the elongation-only zone (P, in cells per hour) was defined as:
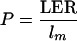 |
6 |
where LER is leaf elongation rate and lm is the length of mature cells (Silk et al., 1989). Under steady-state growth conditions, the number of cells entering at the base of the elongation-only zone per unit time is equal to the number of cells leaving the growth zone at its distal margin where cells reach their mature length. Knowing the rate of cell production and the number of cells in the division zone, the average rate of cell division (D̄, in cells per cell per hour) for the whole meristem can be calculated as (Beemster and Baskin, 1998):
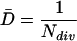 |
7 |
In a population of asynchronous cells, the duration of the cell cycle can be considered as the time necessary for the population to double in size or the time taken for an “average” cell to complete the cycle (Green and Bauer, 1977; Granier and Tardieu, 1998). On these grounds, taking into account the exponential nature of the cell division process, the average duration of the cell cycle (T̄c, in hours) was calculated (Green, 1976; Ivanov and Dubrovsky, 1997; Dubrovsky et al., 1998):
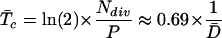 |
8 |
It is evident that, for each individual cell, the residence time in the meristem will be equal to the duration of the cell cycle, i.e. any one cell in the meristem exists only from its formation until it undergoes a subsequent cytokinesis forming two daughter cells. If a nearly constant rate of cell division over time is assumed and the formation of a new cell wall caused by division of the most basal cell in a given cell file is considered, then the residence time in the division zone for such a wall (Tdiv, in hours) can be estimated as (Beemster and Baskin, 1998):
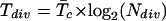 |
9 |
The residence time in the elongation-only zone (Tel, in hours) is proportional to the number of cells per file in this region (Nel) and to the time interval taken for a new cell to be added to the basal margin of the zone (cellochron). Since P is the reciprocal of the cellochron, Tel was estimated as (Beemster and Baskin, 1998):
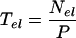 |
10 |
Cells displaced past the distal margin of the division zone cease to divide and they elongate to their mature size in the elongation-only zone. Interpolated cell length profiles (see above) and cell production of individual equivalent cell files formed the basis of strain rate (r, per hour) calculation. According to Silk et al. (1989), strain rate at any given location of the elongation-only zone is proportional to cell production and to the position-derivative of cell length and was calculated as:
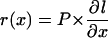 |
11 |
The corresponding strain rate profile describes the relative rate of cell elongation as a function of cell position along the leaf axis.
Statistical Analysis
The measured and calculated parameters have been analyzed for statistically significant differences with a one-way ANOVA. Homogenous subsets were defined on the basis of a post hoc Tukey b test at α = 0.01 significance level with SPSS 8.0 for Windows statistical software (SPSS, Chicago). lsd was calculated from the results of the one-way ANOVA according to Sokal and Rohlf (1995).
ACKNOWLEDGMENTS
The authors wish to thank Rens Voesenek for providing valuable comments on a previous version of this manuscript and Maarten Terlou for developing the software for measurements of cell length and observation of mitotic figures. F.F. is indebted to Frank Millenaar for suggestions about a previous version of this manuscript, to Hendrik Poorter for enlightening discussions about curve fitting, and to Andrea Damascelli for cheerful conversations concerning kinematics.
Footnotes
This work was funded by a PhD scholarship granted by Utrecht University (to F.F.) and by a Marie Curie Training and Mobility Research postdoctoral fellowship (grant no. ERBFMI–CT97–2754 to G.T.S.B.).
LITERATURE CITED
- Atkin OK, Botman B, Lambers H. The causes of inherently slow growth in alpine plants: an analysis based on the underlying carbon economies of alpine and lowland Poa species. Funct Ecol. 1996;10:698–707. [Google Scholar]
- Barlow PW, Brain P, Parker JS. Cellular growth in roots of a gibberellin-deficient mutant of tomato (Lycopersicon esculentum Mill.) and its wild-type. J Exp Bot. 1991;42:339–351. [Google Scholar]
- Beemster GTS, Baskin TI. Analysis of cell division and elongation underlying the developmental acceleration of root growth in Arabidopsis thaliana. Plant Physiol. 1998;116:1515–1526. doi: 10.1104/pp.116.4.1515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beemster GTS, Masle J. The role of apical development around the time of leaf initiation in determining leaf width at maturity in wheat seedlings (Triticum aestivum L.) with impeded roots. J Exp Bot. 1996;47:1679–1688. [Google Scholar]
- Beemster GTS, Masle J, Williamson RE, Farquhar GD. Effects of soil resistance to root penetration on leaf expansion in wheat (Triticum aestivum L.): kinematic analysis of leaf elongation. J Exp Bot. 1996;47:1663–1678. [Google Scholar]
- Ben-Haj-Salah H, Tardieu F. Temperature affects expansion rate of maize leaves without change in spatial distribution of cell length. Plant Physiol. 1995;109:861–870. doi: 10.1104/pp.109.3.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett MD. The time and duration of meiosis. Philos Trans R Soc Lond B. 1977;277:201–226. doi: 10.1098/rstb.1977.0012. [DOI] [PubMed] [Google Scholar]
- Bennett MD, Cox AV, Leitch IJ (1998) Angiosperm DNA C-values database. http://www.rbgkew.org.uk/cval/database1.html
- Bennett MD, Smith JB. Nuclear DNA amounts in angiosperms. Philos Trans R Soc Lond B. 1976;274:227–274. doi: 10.1098/rstb.1976.0044. [DOI] [PubMed] [Google Scholar]
- Bultynck L, Fiorani F, Lambers H. Control of leaf growth and its role in determining variation in plant growth rate from an ecological perspective. Plant Biol. 1999;1:13–18. [Google Scholar]
- Clowes FAL. Estimation of growth fractions in meristems of Zea mays L. Ann Bot. 1976;40:933–938. [Google Scholar]
- Croxdale J. Stomatal patterning in monocotyledons: Tradescantia as a model system. J Exp Bot. 1998;49:279–292. [Google Scholar]
- Doerner P, Jorgensen J, You R, Stepphun J, Lamb C. Control of root growth and development by cyclin expression. Nature. 1996;380:520–523. doi: 10.1038/380520a0. [DOI] [PubMed] [Google Scholar]
- Doležel J, Binarová P, Lucretti S. Analysis of nuclear DNA content in plant cells by flow cytometry. Biol Plant. 1989;31:113–120. [Google Scholar]
- Dubrovsky JG, Contreras-Burciaga L, Ivanov VB. Cell cycle duration in the root meristem of Sonoran desert Cactaceae as estimated by cell flow and the rate of cell production methods. Ann Bot. 1998;81:619–624. [Google Scholar]
- Erickson RO, Sax KB. Rates of cell division and cell elongation in the growth of the primary root of Zea mays. Proc Am Philos Soc. 1956;100:499–514. [Google Scholar]
- Francis D. The cell cycle and plant growth. In: Lambers H, Poorter H, Van Vuuren MMI, editors. Inherent Variation in Plant Growth: Physiological Mechanisms and Ecological Consequences. Leiden, The Netherlands: Backhuys Publishers; 1998. pp. 5–20. [Google Scholar]
- Gallagher JN. Field studies of cereal leaf growth: I. Initiation and expansion in relation to temperature and ontogeny. J Exp Bot. 1979;30:625–636. [Google Scholar]
- Gandar PW. The analysis of growth and cell production in root apices. Bot Gaz. 1980;141:131–138. [Google Scholar]
- Granier C, Tardieu F. Spatial and temporal analyses of expansion and cell cycle in sunflower leaves. Plant Physiol. 1998;116:991–1001. doi: 10.1104/pp.116.3.991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green PB. Growth and cell pattern formation on an axis: critique of concepts, terminology, and mode of study. Bot Gaz. 1976;137:187–202. [Google Scholar]
- Green PB, Bauer K. Analyzing the changing cell cycle. J Theor Biol. 1977;68:299–315. doi: 10.1016/0022-5193(77)90167-9. [DOI] [PubMed] [Google Scholar]
- Hegi G, Conert HJ. Illustrierte Flora von Mitteleuropa. In: Conert HJ, Hegi G, editors. Band 1, Teil 3. Spermatophyta: Angiospermae: Monocotyledons 1(2). Poaceae. Berlin: Parey Buchverlag; 1998. [Google Scholar]
- Horst GL, Nelson CJ, Asay KH. Relationship of leaf elongation to forage yield of tall fescue genotypes. Crop Sci. 1978;18:715–719. [Google Scholar]
- Ivanov VB, Dubrovsky JG. Estimation of the cell cycle duration in the root apical meristem: a model of linkage between cell cycle duration, rate of cell production and rate of root growth. Int J Plant Sci. 1997;158:757–763. [Google Scholar]
- Jacobs TW. Cell cycle control. Annu Rev Plant Physiol. 1995;46:317–339. [Google Scholar]
- Körner C, Woodward FI. The dynamics of leaf extension in plants with diverse altitudinal ranges: II. Field studies in Poa species between 600 and 3200 m altitude. Oecologia. 1987;72:279–283. doi: 10.1007/BF00379279. [DOI] [PubMed] [Google Scholar]
- Larkin JC, Marks MD, Nadeau J, Sack F. Epidermal cell fate and patterning in leaves. Plant Cell. 1997;9:1109–1120. doi: 10.1105/tpc.9.7.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lockhart JA. An analysis of irreversible plant cell elongation. J Theor Biol. 1965;8:264–275. doi: 10.1016/0022-5193(65)90077-9. [DOI] [PubMed] [Google Scholar]
- MacAdam JW, Volenec JJ, Nelson CJ. Effects of nitrogen supply on mesophyll cell division and epidermal cell elongation in tall fescue leaf blades. Plant Physiol. 1989;89:549–556. doi: 10.1104/pp.89.2.549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mironov V, De Veylder L, Van Montagu M, Inze D. Cyclin-dependent kinases and cell division in plants. Plant Cell. 1999;11:509–521. doi: 10.1105/tpc.11.4.509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mowforth MA, Grime JP. Intra-population variation in nuclear DNA amount, cell size and growth rate in Poa annua L. Funct Ecol. 1989;3:289–295. [Google Scholar]
- Poorter H, Remkes C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia. 1990;83:553–559. doi: 10.1007/BF00317209. [DOI] [PubMed] [Google Scholar]
- Richards FJ. A flexible growth function for empirical use. J Exp Bot. 1959;29:290–300. [Google Scholar]
- Schnyder H, Seo S, Rademacher IF, Kühbauch W. Spatial distribution of growth rates and of epidermal cell lengths in the elongation zone during leaf development in Lolium perenne L. Planta. 1990;181:423–431. doi: 10.1007/BF00195897. [DOI] [PubMed] [Google Scholar]
- Silk WK. Quantitative description of plant development. Annu Rev Plant Physiol. 1984;35:479–518. [Google Scholar]
- Silk WK, Erickson RO. Kinematics of plant growth. J Theor Biol. 1979;76:481–501. doi: 10.1016/0022-5193(79)90014-6. [DOI] [PubMed] [Google Scholar]
- Silk WK, Lord EM, Eckard KJ. Growth patterns inferred from anatomical recordings. Plant Physiol. 1989;90:708–714. doi: 10.1104/pp.90.2.708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sokal R, Rohlf FJ. Biometry: The Principles and Practice of Statistics in Biological Research. W.H. New York: Freeman; 1995. [Google Scholar]
- Van Arendonk J, Niemann GJ, Boon JJ, Lambers H. Effects of N-supply on anatomy and chemical composition of leaves of four grass species, belonging to the genus Poa, as determined by image-processing and pyrolysis-mass spectrometry. Plant Cell Environ. 1997;20:881–897. [Google Scholar]
- Van 't Hof J, Sparrow AH. A relationship between DNA content, nuclear volume and minimum cell cycle time. Proc Natl Acad Sci USA. 1963;49:897–902. doi: 10.1073/pnas.49.6.897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Venus JC, Causton DR. Plant growth analysis: the use of the Richards function as an alternative to polynomial exponentials. Ann Bot. 1979;43:623–632. [Google Scholar]
- Volenec JJ, Nelson CJ. Cell dynamics in leaf meristem of contrasting tall fescue genotypes. Crop Sci. 1981;21:381–385. [Google Scholar]
- Volenec JJ, Nelson CJ. Response of tall fescue leaf meristem to N fertilization and harvest frequency. Crop Sci. 1983;23:720–724. [Google Scholar]
- Webster PL, MacLeod RD. Characteristics of root apical meristem cell population kinetics: a review analyses and concepts. Environ Exp Bot. 1980;20:335–358. [Google Scholar]