

Abstract
Recent structure-function analysis of heterologously expressed K+-selective inward-rectifying channels (KIRCs) from plants has revealed that external protons can have opposite effects on different members of the same gene family. An important question is how the diverse response of KIRCs to apoplastic pH is reflected at the tissue level. Activation of KIRCs by acid external pH is well documented for guard cells, but no other tissue has yet been studied. In this paper we present, for the first time to our knowledge, in planta characterization of the effects of apoplastic pH on KIRCs in roots. Patch-clamp experiments on protoplasts derived from barley (Hordeum vulgare) roots showed that a decrease in external pH shifted the half-activation potential to more positive voltages and increased the limit conductance. The resulting enhancement of the KIRC current, together with the characteristic voltage dependence, strongly relates the KIRC of barley root cells to AKT1-type as opposed to AKT3-type channels. Measurements of cell wall pH in barley roots with fluorescent dye revealed a bulk apoplastic pH close to the pK values of KIRC activation and significant acidification of the apoplast after the addition of fusicoccin. These results indicate that channel-mediated K+ uptake may be linked to development, growth, and stress responses of root cells via the activity of H+-translocating systems.
Changes in apoplastic pH are involved in many physiological processes in plants, such as development, growth, leaf movement, gas exchange, and pathogen defense (Grignon and Sentenac, 1991; Kutschera, 1994; Palmgren, 1998, and refs. therein). Stimuli for these changes are diverse and include light, plant hormones, mechanical stress, osmotic potential, and nutrient availability. Changes of apoplastic pH can be achieved via modulation of the activities of H+-extruding ATPase or H+-coupled nutrient transporters in the plasma membrane, as well as by export of acid metabolites and CO2 evolution. The role of apoplastic pH in H+-coupled uptake of nutrients such as high-affinity uptake of K+ (Maathuis and Sanders, 1994) is evident: apoplastic protons not only act as a substrate for the transport system but also affect the electrical driving force for this charged transport process through modulation of the membrane potential. Although low-affinity uptake of K+ is not physically coupled to that of H+, charge compensation for K+ uptake is achieved by H+ extrusion (Behl and Raschke, 1987; Kochian and Lucas, 1988).
Low-affinity K+ uptake by plants is mediated by KIRCs that activate at negative membrane potentials when K+ concentrations are in the high micromolar or millimolar range, ensuring that K+ movement is directed inward (Maathuis et al., 1997; Hirsch et al., 1998). In strong contrast to KIRCs in animal cells, which are blocked by extracellular protons (Coulter et al., 1995; Sabirov et al., 1997), KIRCs in the plasma membrane of plant guard cells have been shown to be activated by acidification of the external medium, both in planta and after heterologous expression (Blatt, 1992; Hedrich et al., 1995; Müller-Röber et al., 1995; Véry et al., 1995; Ilan et al., 1996; Hoth et al., 1997). It was further demonstrated that H+ activation of a KIRC cloned from potato guard cells (KST1) is mediated by a His residue in the outer-pore region of this channel (Hoth et al., 1997). Sequence alignment of many cloned animal and plant KIRCs revealed that the relevant His residue is well conserved among plant channels but is not present in their animal counterparts. The alignment included cDNAs of AKT1-type plant KIRCs, which have been shown to be expressed in root tissue (Basset et al., 1995; Lagarde et al., 1996) and are likely to play a significant role in K+ uptake (Hirsch et al., 1998). The authors therefore concluded that proton activation from the extracellular side of the membrane is a typical feature of all plant KIRCs. This hypothesis was strengthened by a study of SKT1, a KIRC cloned from potato (Zimmermann et al., 1998), the mRNA of which was detected in root tissue. This KIRC was activated by external protons when heterologously expressed in baculovirus-infected insect cells.
In contrast to the findings on H+-induced activation of IKIRC, work on suspension-cultured cells from Arabidopsis demonstrated that currents through the KIRCs of these cells decreased upon acidification of the external medium (Giromini et al., 1997). This observation coincides with a report indicating that currents through the Arabidopsis KIRC AKT3 are blocked by H+ when expressed in Xenopus laevis oocytes (Marten et al., 1998). In neither case is the tissue localization of the respective channels known.
We studied the effect of apoplastic pH on the IKIRC of root cells and show activation of this channel type by external protons (as was previously described for guard cells) and a partial dependence of the pH effect on the external KCl concentration. Our study also included measurements of the apoplastic pH in root tissue, which enabled us to put channel activity into a physiological context.
MATERIALS AND METHODS
Growth of Plant Material
Seeds of barley (Hordeum vulgare L. cv Puffin) were sown in washed grit-sand and grown at 22°C/18°C day/night temperatures with a 14-h photoperiod and 200 to 350 μmol m−2 s−1 PAR. RH was maintained at 60% to 80%. After 5 d young plants were transferred to hydroponic conditions (20 plants per 2 L of growth medium, changed twice a week). The growth medium contained 9 mm NaNO3 or KNO3, 0.5 mm NaH2PO4, 0.5 mm KH2PO4, 1 mm CaCl2, 1 mm Ca(NO3)2, 1.5 mm MgSO4, 0.1 mm FeNaEDTA, and 0.1 mm NaCl, plus trace elements (23 μm H3BO3, 10 μm MnSO4, 0.7 μm ZnSO4, 0.25 μm CuSO4, and 0.65 nm [NH4]6MO7O2). In patch-clamp experiments 4- to 6-week-old plants were used, whereas determination of apoplastic pH was performed on younger plants (10–14 d old).
Preparation of Protoplasts
For protoplast preparation 3- to 4-cm distal segments of primary roots (excluding about 1 cm at the root tip) were chopped and incubated with 1.5% (w/v) cellulase (“Onozuka” Yakult Honsha Co., Tokyo, Japan), 0.15% (w/v) pectolyase (Sigma), and 0.1% (w/v) BSA in solution A for 2 h at room temperature. Protoplasts were released from the tissue by gentle squeezing in solution B, washed, and collected. Solutions A and B contained 10 mm KCl, 2 mm MgCl2, 2 mm CaCl2, and 2 mm Mes at pH 5.7 (Tris) and were adjusted to 600 mOsm (solution A) and 400 mOsm (solution B) with sorbitol. Alternatively, cortical tissue was stripped off and discarded before enzyme incubation. Protoplasts obtained from whole-root and stelar preparations were very similar in size, appearance, and features, and we therefore assumed that both preparations produced stelar protoplasts only, probably originating from xylem parenchyma (Wegner and Raschke, 1994).
Patch-Clamp Setup and Data Acquisition and Analysis
Standard patch-clamp techniques (Hamill et al., 1981) were applied. Details for pipette preparation, voltage-clamp, and recording equipment were as as described by Amtmann et al. (1997). Protoplasts of similar size (approximately 30 μm in diameter) were chosen for the experiments. Whole-cell inward currents were elicited by 3-s bipolar voltage pulses from holding potentials more positive than EK (equilibrium potential for K+) filtered at 300 Hz, and recorded with a sample frequency of 1.5 kHz. The access resistance was measured with the amplifier but not corrected for; measurements were discarded when errors in the clamp voltage larger than 5% arose from high access resistance. Conductance of the instantaneous current component was ohmic over the whole voltage range analyzed. Therefore, leak subtraction was used as the general method for obtaining time-dependent IKIRC. Steady-state IKIRC were measured 3 s after onset of the voltage pulse and averaged over three identical voltage protocols. Values are means ± se. Curve fitting was carried out using the program FigP (Biosoft, Cambridge, UK).
Experimental Solutions
Patch pipettes contained 100 mm KCl, 1 mm MgCl2, 1 mm CaCl2, 1.4 mm EGTA (4 mm KOH), 1.5 mm MgATP, and 2 mm Hepes or Tes buffered to pH 7.4 with Tris. The free Ca2+ concentration was 176 nm, as calculated with the CALCIUM program (Föhr et al., 1993). Bath solutions contained 10 or 1 mm KCl, 1 mm MgCl2, and 1 mm CaCl2, at various pH values (2–5 mm Mes, Tes, or Hepes and Tris or NaOH). Liquid junction potentials were measured and corrected when larger than 3 mV according to the method of Amtmann and Sanders (1997). All solutions were adjusted to 500 mOsm with sorbitol and sterile-filtered before use.
Determination of Apoplastic pH
Calibration and Dye Loading
Stock solutions (10 mm) of the pH-sensitive dye NERF-DM (Molecular Probes, Leiden, Netherlands) were prepared in 20 mm Mes buffer, pH 6.3, and stored at 4°C. For calibration measurements the dye was diluted to 10 μm in 100 mm phosphate buffer at an appropriate pH. Calibration points were fitted with Equation 3 (see Results). pH sensitivity (ratiomax − ratiomin = 0.54) and affinity (pK = 4.9) of the in vitro calibration were satisfactory (compare with Whitaker et al., 1992). For in situ calibration, 1-cm-long fragments of barley roots (2 cm above the tip) were incubated for 2 h in 10 μm NERF-DM. Imaging of root fragments was performed immediately after they were rinsed in dye-free buffer. We found that root fragments had to be exposed to buffer concentrations greater than 200 mm to influence apoplastic pH. However, when 1 mm KCN, 1 mm salicylhydroxamic acid, and 1 μm carbonyl cyanide p-trifluoromethoxyphenylhydrazone (all from Sigma) were added during dye incubation, in situ calibration was achieved in 100 mm phosphate buffers. Compared with the in vitro calibration, ratio values were shifted to higher values in situ (by 0.24 unit on average) and the sensitivity of the dye was slightly reduced (ratiomax − ratiomin = 0.49), whereas the affinity for H+ was only slightly affected (pK 5.1). The in situ calibration curve is shown in Figure 6. For in vivo measurements root fragments were incubated for 1 h in 100 μm Mes-Tris, pH 5.9, and 10 μm NERF-DM with and without 5 μm fusicoccin. The ethanol concentration in both solutions was 0.5% (v/v).
Figure 6.

In situ calibration of the fluorescent dye DM-NERF and measurements of apoplastic pH in barley roots. In situ calibration (○, means ± se; n = 9) was obtained by ratio-imaging NERF-DM fluorescence from barley root cells incubated in 100 mm phosphate buffers and respiratory inhibitors (see Methods). The curve was fitted with Equation 3 (in which X was replaced by the fluorescence ratio), and the 95% confidence band is indicated with dashed lines. Solid symbols show mean ratio/pH values (±se; n = 4) measured in barley root segments without (♦) and with (▴) 5 μm fusicoccin.
Dye localization in the cell wall was confirmed by comparison of NERF-DM 70-kD dextran-loaded cells and NERF-DM-loaded cells. No difference in dye localization was apparent. Furthermore, barley root protoplasts incubated for 2 h in 200 μm NERF-DM did not show any fluorescence, even at an increased laser intensity (data not shown).
Imaging
Confocal fluorescence imaging (model MRC 1000, Bio-Rad) was controlled by CoMOS and TCSM software (Bio-Rad). The confocal system was interfaced with an upright microscope (Optiphot 300, Nikon). Imaging was performed using a ×60 (1.4 numerical aperture) oil-immersion objective (PlanApo, Nikon) and a ×3.5 electronic zoom. Fluorescence images were acquired using the 514- and 488-nm excitation lines of an argon laser at intensities between 1% and 10%, depending on dye loading into the cell walls. Emission was recorded using the integral emission filter (540 DF 30, Nikon) of the confocal microscope. The pinhole size (4) and gain setting (900) were chosen to give optimal fluorescence signals at all pH values in the calibration curve and were identical for all experiments. Fluorescence ratio values (514/488 nm) were calculated pixel by pixel using TCSM software. Average ratio values for NERF-DM-loaded cell walls were extracted using the Histogram command in CoMOS. Care was taken not to include areas close to the edge of the cell wall, since low fluorescence intensity caused artifacts in this region.
RESULTS
Typical IKIRC in the Plasma Membrane of Barley Root Cells
Hyperpolarization of the plasma membrane to voltages more negative than −80 mV evoked large inward currents with the typical features of KIRCs, such as time-dependent double-exponential activation, single-exponential deactivation, high selectivity for K+ over anions and other cations, and blockage by tetraethylammonium ions. The IKIRC has been described for many plants and in particular for barley root xylem parenchyma cells (Wegner and Raschke, 1994; Wegner et al., 1994).
Response of IKIRC to Varying External pH
Figure 1 shows a typical response of IKIRC to changes in external pH. A shift of pH from 7.5 to 5.5 increased IKIRC considerably. Furthermore, the voltage needed for half-activation of IKIRC was more positive at pH 5.5 than at pH 7.5. The effects of external pH on IKIRC were fully reversible. The reversal potentials (Erev) of the time-dependent currents as determined in tail-current experiments (resolution ± 5 mV) were not significantly different at different pH values: for 10 mm external KCl (n = 5 protoplasts), Erev = −54 ± 2 mV at pH 5.5 and −58 ± 3 mV at pH 7.5; for 1 mm external KCl (n = 4 protoplasts), Erev = −101 ± 4 mV at pH 5.5 and −98 ± 2 mV at pH 7.5.
Figure 1.

Typical inward currents in barley root protoplasts at pH 5.5 and 7.5. Currents were elicited by clamping the membrane potential with negative square voltage pulses of 3 s. The holding potential was −20 mV, and the voltage increment was −20 mV. The external medium contained 10 mm KCl, 1 mm CaCl2, and 1 mm MgCl2 and was buffered to pH 7.5 (5 mm Mes/Tris) or pH 5.5 (5 mm Tes/Tris). No leak subtraction was performed. Fitted time constants (τ1, τ2) for the time-dependent current activation at −180 mV were 50 ms, 250 ms (top), 37 ms, 320 ms (middle), and 37 ms, 240 ms (bottom).
Fitting the IKIRC-Voltage Relations
Figure 2 depicts typical current-voltage (I-V) relationships of the KIRC at pH 5.5 and 7.5 (same protoplast as in Fig. 1) recorded with 10 or 1 mm KCl in the bathing medium. All I-V relationships were well fitted by a simple function in which the Goldman-Hodgkin-Katz equation for the open-channel current is multiplied by the Boltzman distribution of the open probability, the maximal open probability, and the total number of channels in the protoplast. Thus,
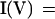 |
1 |
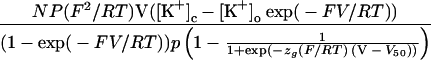 |
where I is the current, V is the membrane voltage, N is the total number of channels, P is the permeability of the open channel for K+, [K+]c and [K+]o are the cytosolic and external K+ activities, respectively, p is the maximal open probability, zg is the gating charge, and R, T, and F are the gas constant, absolute temperature, and Faraday's constant, respectively. The product NPp was treated as one adjustable parameter. Glim was calculated from:
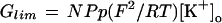 |
2 |
Glim describes the constant conductance at voltages far more negative than EK with maximal number of channels open. From whole-cell recordings it cannot be decided whether a change in Glim is due to a change in P or in p. Fitted parameters for zg were very similar at all external pH values tested, and the fits were not significantly worse if zg was fixed to a value of 1.8. Both V50 and Glim, however, differed significantly at varying external pH values and the following analysis of the effect of pH on IKIRC concentrates on these two parameters.
Figure 2.

Typical current-voltage relations of KIRC with 1 mm (○) or 10 mm (▵) external K+ at pH 5.5 and 7.5 (same protoplast as in Fig. 1; protoplast diameter, 27 μm). Means ± se of leak-subtracted currents obtained in three identical voltage protocols are shown. Curves were obtained from Equation 1. Fitted parameters: V50 = −138 mV, Glim = 215 pS, NPp = 5.7 × 10−17 m3 s−1 for 1 mm [K+]o at pH 5.5; V50 = −171 mV, Glim = 101 pS, NPp = 2.7 × 10−17 m3 s−1 for 1 mm [K+]o at pH 7.5; V50 = −139 mV, Glim = 2000 pS, NPp = 5.9 × 10−17 m3 s−1 for 10 mm [K+]o at pH 5.5; V50 = −157 mV, Glim = 1051 pS, NPp = 3.1 × 10−17 m3 s−1 for 10 mm [K+]o at pH 7.5.
V50 and Glim at pH 5.5 and 7.5
Figure 3 displays the results of a statistical evaluation of V50 and Glim measured at external pH values of 5.5 and 7.5 and external KCl concentrations of 1 and 10 mm. For both external KCl concentrations, mean absolute magnitudes of V50 (Fig. 3A) were significantly more positive at pH 5.5 than at pH 7.5. The difference in absolute magnitudes of Glim determined at pH 5.5 and 7.5 was not significant because variation of Glim among different protoplasts was high, probably because of varying channel densities in the membrane (for means, see the legend of Fig. 3A). This problem was overcome by analyzing changes of V50 and Glim in response to individual medium-exchange events in each protoplast, where the total number of channels could be assumed to be constant (Fig. 3B). When the external pH changed from 5.5 to 7.5, V50 shifted to a more negative value by an average of 28 ± 3 mV in 1 mm external KCl (n = 5) and 16 ± 2 mV in 10 mm external KCl (n = 6). The Glim at pH 7.5 was, on average, 63% ± 7% (1 mm KCl, n = 5) or 61% ± 5% (10 mm KCl, n = 6) of the Glim at pH 5.5.
Figure 3.

Effect of external pH on V50 and Glim. Each value shown is the mean ± se of 6 to 10 determinations from five or six different protoplasts (one or two determinations per protoplast). A, Absolute values of V50 at pH 5.5 and 7.5 in 1 or 10 mm external KCl. Absolute values of Glim (not shown) varied between 334 ± 101 pS (n = 5) at pH 7.5 and 488 ± 137 pS (n = 5) at pH 5.5 in 1 mm external KCl and 1341 ± 511 pS (n = 6) at pH 7.5 and 2183 ± 682 pS (n = 6) at pH 5.5 in 10 mm external KCl. B, Changes in V50 and Glim within each protoplast caused by changes in external pH.
The effect of pH on V50 was clearly dependent on the concentration of KCl in the bath, whereas no such dependence was observed for the pH effect on Glim. Also, there was no correlation between the size of ΔV50 and the Glim ratio: in the same protoplast pH could have a relatively strong effect on V50 and a relatively weak effect on Glim or vice versa: the correlation coefficient was 0.7 (10 mm external KCl, n = 6) and 0.6 (1 mm KCl, n = 5), respectively, and in both P > 0.1. Therefore, external pH affects KIRC activity via two independent modulation mechanisms: the first one shifts the Boltzman distribution of the open probability along the voltage axis (effect on V50), and the second changes the asymptote of the Boltzman distribution by affecting the maximal open probability or the conductance of the open channel (effect on Glim). Both effects of external pH were independent of the type of pH buffer used (Mes or HCl for pH 5.5, Hepes or Tes for pH 7.5, Tris or NaOH) and the buffer concentration (between 2 and 5 mm) used.
To obtain a titration curve for the effect of external pH on IKIRC, V50, and Glim, one protoplast was exposed to steps of 0.5 pH unit over a wide range of external pH values (4.3–7.8). The pH range was sampled twice, going from basic to acidic values and vice versa, and a slight hysteresis was observed. IKIRC elicited by hyperpolarizing voltage pulses were recorded and the corresponding I-V relationships were determined and fitted with Equation 1. The mean IKIRC at a given voltage (Fig. 4A) was then plotted against external pH. At all voltages, IKIRC(pH) followed a simple function expected from a single titratable binding site of H+:
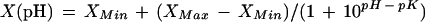 |
3 |
where X is the IKIRC and XMin and XMax are the minimal and maximal IKIRC at very high and very low external pH, respectively. pK values of IKIRC were very similar for all voltages (5.9 and 6.0). However, the proportion of IKIRC affected by external pH was voltage dependent. The relative XMin (XMin as a percentage of the XMax) decreased with positive-going voltages from 46% at −200 mV to 44% at −180 mV, 40% at −160 mV, 34% at −140 mV, 19% at −120 mV, and 0% at −100 mV. Equation 3 (with X now representing V50 or Glim) was also sufficient to fit V50 and Glim derived from the I-V relationships of this protoplast at different pH values (Fig. 4, B and C). pK values were 6.5 and 5.8 for V50 and Glim, respectively. Glim Min was 48% of Glim Max.
Figure 4.

Titration of the effect of external pH on IKIRC, V50, and Glim at 10 mm external KCl. A, I-pH relations of a single protoplast at different voltages. Data points are means of two current values (acid-alkaline or alkaline-acid bath exchange, pH 6.7, one only), each of which is the mean of currents obtained in three identical voltage protocols. Error bars indicate the position of the two individual values. Curves were obtained by fitting Equation 3 to the data. Fitted parameters for −100 (−120, −140, −160, −180, and −200) mV: IMin (pA) = 0, (−20, −61, −102, −140, −170) pA, IMax = −53 (−108, −179, −254, −317, −371) pA, pK = 5.9 (6.0, 5.9, 5.9, 5.9, 5.9). B and C, Titration of the pH effect on V50 (B) and Glim (C) derived from the I-V relationships of this protoplast (solid symbols). Solid lines are the fits with Equation 3; X represents V50 or Glim. The protoplast used for this experiment was representative of relative values (ΔV50 and relative Glim) but resided at the periphery of the data set for all protoplasts with respect to absolute values (compare with Fig. 3). For comparison, mean absolute data for V50 and Glim at pH 5.5 and 7.5 are shown (open symbols). Dashed line in B is the solid line shifted vertically by −16 mV.
Apoplastic pH in Barley Roots
To assess the physiological relevance of low-pH-induced activation of IKIRC, we measured apoplastic pH in barley root tissue using the pH-dependent fluorescent dye DM-NERF. Confocal microscopy confirmed that the dye accumulated selectively in the cell walls of barley roots (Fig. 5). The mean fluorescence ratio measured in root fragments was 0.93 ± 0.02 (n = 4 plants, three fragments per plant). The fluorescence ratio was significantly lower (0.82 ± 0.01) in root fragments of the same four plants that had been incubated for 1 h with 5 μm fusicoccin. The apoplastic pH was determined as 5.9 ± 0.1 for control root tissue and as 5.3 ± 0.05 for fusicoccin-treated tissue from a comparison with the in situ calibration performed in root fragments exposed to 100 mm phosphate buffer and respiratory blockers added (Fig. 6).
Figure 5.

Pair of fluorescence images of barley root cortical cell wall loaded with NERF-DM. Emission detected at 540 nm. A, Excitation at 514 nm. B, Excitation at 488 nm. Bar represents 10 μm.
DISCUSSION
Effects of Apoplastic pH on KIRCs in Different Cell Types
Effects of apoplastic pH on IKIRC were described previously for intact guard cells from fava bean (Blatt, 1992), for guard cell protoplasts from fava bean (Ilan et al., 1996) and potato (Hoth et al., 1997), and after heterologous expression of guard cell KIRC mRNA in X. laevis oocytes (KAT1: Hedrich et al., 1995; Véry et al., 1995; KST1: Müller-Röber et al., 1995; Hoth et al., 1997). Although reports regarding guard cells are unanimous in describing an activating effect of external protons on KIRCs, they differ in the number and type of parameters reported as affected (V50, Glim, and the time constants of activation and deactivation, τact and τdeact: Blatt, 1992, and Ilan et al., 1996; Glim: Véry et al., 1995; V50, τact, and τdeact: Hedrich et al., 1995; τact: Müller-Röber et al., 1995; and V50: Hoth et al., 1997). The only report relating to pH effects on KIRC activity in a cell type other than guard cells was from cultured Arabidopsis cells (Giromini et al., 1997) and described the opposite effect: IKIRC decreased upon acidification of the external medium because of a reduction of Glim, whereas V50 was not affected. Our description of modulation of IKIRC by apoplastic pH is the first to our knowledge for root cells and confirms speculations about the extracellular proton activation of the KIRC in root cells that arose from studies of the potato KIRC SKT1 (Zimmermann et al., 1998).
Mechanisms of KIRC Activation by External Protons
The effect of external pH on V50 reflects proton-dependent modulation of channel gating. Ilan et al. (1996) suggested that external protons affect V50 by decreasing the negative surface-charge density in the vicinity of the channel gates. Our observation that the effect of external protons on V50 was attenuated by increasing external KCl concentrations points to the same conclusion. One possibility is that protons act on V50 by screening the surface potential of the entire membrane, which would result in a different proportion of clamp voltage being sensed by the channel. The second possibility is that the pH effect on V50 involves protonation and charge screening of intrinsic channel sites that are exposed to the extracellular side of the membrane. Analysis of mutations performed on KST1 suggested that a His residue situated in the outer-pore region of the channel is crucial for the proton action on V50 (Hoth et al., 1997) and the pK of 6.5, which in our study was derived from the pH titration of V50 and is approximately the pK of His protonation (Creighton, 1993).
The observed effect of external pH on Glim could be due to pH dependence of either P or p (Eq. 2), and a final statement can be derived only from single-channel studies.
pH titration of currents through the KIRC at different voltages (Fig. 4A) reflects the combined effect of pH on V50 and Glim. However, the pK of IKIRC(pH) was always approximately the pK of Glim(pH) and was voltage independent. pK values for V50 and Glim were probably not different enough to cause a significant shift in the pK of IKIRC(pH) with voltage.
The fact that Glim is not reduced to 0 at high pH (Fig. 4C) is reflected in the residual IKIRC at very negative voltages. Although current activation still exhibited double-exponential kinetics in these conditions (compare with Fig. 1), we cannot disregard the possibility that the pH-independent component of IKIRC is due to a different channel type with similar gating kinetics (compare with Amtmann et al., 1997).
Contrasting Responses of Plant KIRCs to External Protons
Recent studies established that the Arabidopsis K+-channel AKT3 displays unique properties with respect to pH sensitivity (Marten et al., 1998), even though its cDNA sequence is highly similar to that of the other Arabidopsis KIRCs, KAT1 and AKT1 (Ketchum and Slayman, 1996). When expressed in X. laevis oocytes, AKT3 shows weak inward rectification and blockage by external protons. Acidification of the external medium does not affect V50 but decreases Glim because of a decrease in single-channel conductance (Marten et al., 1998). From these findings one may speculate that the KIRC of cultured Arabidopsis cells, which is inhibited by external protons (Giromini et al., 1997), is closely related to or identical to AKT3. Similarities between other characteristics of both channel currents have been highlighted previously (Colombo and Cerana, 1991; compare with Ketchum and Slayman, 1996). The expression pattern of AKT3 in the plant has not yet been established. pH dependence and gating properties clearly relate the barley root KIRC to AKT1 and SKT1 but not to AKT3-type channels.
Implications of the Effect of External pH on IKIRC for Low-Affinity K+ Uptake
KIRCs have been shown to be the major pathway for low-affinity K+ uptake by plant cells (Maathuis et al., 1997). Any change in KIRC activity caused by changes in the external pH can therefore be expected to change K+-uptake rates. However, it has to be taken into account that a change in the amount of K+ inward current may influence the membrane potential and, since the KIRC is voltage dependent, this will also affect its activity. Roelfsema and Prins (1998) calculated that apoplastic acidification would ultimately reduce K+ influx into guard cell protoplasts from Arabidopsis since activation of the KIRC caused a strong depolarization. However, experimental modification of apoplastic pH as applied by Roelfsema and Prins (1998) does not reflect the physiological conditions in which acidification is generated. In many cases apoplastic acidification is achieved via enhanced proton-pumping activity (Palmgren, 1998), which will not only acidify the apoplast but also hyperpolarize the membrane. To assess pH-linked effects on low-affinity K+ uptake, both parameters, apoplastic pH and membrane potential, would have to be measured simultaneously.
Apoplastic pH in Barley Root Tissue
To determine whether the pH effects on the KIRC in root cells could have a physiological role we measured the steady-state pH in cell walls of barley roots and found that the pH-dependent fluorescent dye DM-NERF was suitable for measurements of apoplastic pH in root tissue insofar as it failed to permeate the plasma membrane. The values of apoplastic pH determined here are in the same range as those derived for other tissues (Grignon and Sentenac, 1991), e.g. in the vicinity of guard cells (pH 6.0–7.0, Edwards et al., 1988), leaf epidermis (5.2–5.9, Mühling et al., 1995), various leaf tissues (5.5–6.5, Hoffmann and Kosegarten, 1995), and root epidermis (4.5–4.9, Taylor et al., 1996). Furthermore, we found that the apoplastic pH of barley root cells changes when fusicoccin is added. This result agrees well with recent data from maize roots, in which the addition of 2 μm fusicoccin caused apoplast acidification from pH 5.6 to 4.8, as measured with pH-sensitive microelectrodes (Felle, 1998). The conclusion that the plasma membrane H+-ATPase strongly regulates apoplastic pH is further supported by our observation that tissue calibration could be achieved only in the presence of respiratory blockers (see Methods). Accordingly, Felle (1998) reported increased apoplastic pH after the addition of KCN and salicylhydroxamic acid. Taking into account that the pH effect on the KIRC has a pk of 5.9, we can predict a tight linkage between activities of KIRC and H+-ATPase via apoplastic pH.
Putative Physiological Roles of pH Modulation of KIRC Activity
One of the few external factors of physiological relevance that has been specifically shown to influence the plasma membrane H+-ATPase is salt stress (Palmgren, 1998). Salt-induced gene expression and increased activity of the H+-ATPase have been reported for many plant species (Braun et al., 1986; Niu et al., 1993; Binzel, 1995; Ayala et al., 1996; Wu and Seliskar, 1998) and seem to be positively correlated with salt tolerance (Niu et al., 1993). Our results are in accord with the notion that activation of the H+-ATPase would acidify the apoplast and increase IKIRC. This might reverse the inhibitory effect of salt-induced membrane depolarization (Cakirlar and Bowling, 1981; Katsuhara and Tazawa, 1990; Kourie and Findlay, 1990) on the KIRCs and allow the root cells to maintain a relatively high K+/Na+-influx ratio under saline conditions (Amtmann and Sanders, 1999).
ACKNOWLEDGMENTS
We thank Alison Karley for assistance with plant maintenance and Frans Maathuis, Steve Roberts, and Richard Parton for useful discussions.
Abbreviations:
- Glim
limit conductance
- IKIRC
KIRC current(s)
- KIRC
K+-selective inward-rectifying channel
- V50
half-activation potential
Footnotes
This work was supported by the European Union and the Biotechnology and Biological Sciences Research Council of the United Kingdom.
LITERATURE CITED
- Amtmann A, Laurie S, Leigh R, Sanders D. Multiple inward channels provide flexibility in Na+/K+ discrimination at the plasma membrane of barley suspension culture cells. J Exp Bot. 1997;48:481–497. doi: 10.1093/jxb/48.Special_Issue.481. [DOI] [PubMed] [Google Scholar]
- Amtmann A, Sanders D. A unified procedure for the correction of liquid junction potentials in patch clamp experiments on endo- and plasma membranes. J Exp Bot. 1997;48:361–364. doi: 10.1093/jxb/48.Special_Issue.361. [DOI] [PubMed] [Google Scholar]
- Amtmann A, Sanders D. Mechanisms of Na+ uptake by plant cells. Adv Bot Res. 1999;29:75–112. [Google Scholar]
- Ayala F, O'Leary JW, Schumaker S. Increased vacuolar and plasma membrane H+-ATPase activities in Salicornia bigelovii Torr. in response to NaCl. J Exp Bot. 1996;47:25–32. [Google Scholar]
- Basset M, Conejero G, Lepetit M, Fourcroy P, Sentenac H. Organization and expression of the gene coding for the potassium transport system AKT1 of Arabidopsis thaliana. Plant Mol Biol. 1995;29:947–958. doi: 10.1007/BF00014968. [DOI] [PubMed] [Google Scholar]
- Behl R, Raschke K. Close coupling between extrusion of H+ and uptake of K+ by barley roots. Planta. 1987;172:531–538. doi: 10.1007/BF00393871. [DOI] [PubMed] [Google Scholar]
- Binzel M. NaCl-induced accumulation of tonoplast and plasma membrane H+-ATPase message in tomato. Physiol Plant. 1995;94:722–728. [Google Scholar]
- Blatt MR. K+ channels of stomatal guard cells. Characterization of the inward rectifier and its control by pH. J Gen Physiol. 1992;99:615–644. doi: 10.1085/jgp.99.4.615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braun Y, Hassidim M, Lerner HR, Reinhold L. Studies on H+ translocating ATPases in plants of varying resistance to salinity. I. Salinity during growth modulates the proton pump in the halophyte Atriplex nummularia. Plant Physiol. 1986;87:104–108. doi: 10.1104/pp.81.4.1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cakirlar H, Bowling DJF. The effect of salinity on the membrane potential of sunflower roots. J Exp Bot. 1981;32:479–485. [Google Scholar]
- Colombo R, Cerana R. Plant Physiol. 1991;97:1130–1135. doi: 10.1104/pp.97.3.1130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coulter KL, Perier F, Radeke CM, Vandenberg CA. Identification and molecular localization of a pH-sensing domain for the inward rectifier potassium channel HIR. Neuron. 1995;15:1157–1168. doi: 10.1016/0896-6273(95)90103-5. [DOI] [PubMed] [Google Scholar]
- Creighton TE (1993) Proteins: Structures and Molecular Properties. WH Freeman, New York
- Edwards MC, Smith GN, Bowling DJF. Guard cells extrude protons prior to stomatal opening: a study using fluorescence microscopy and pH micro-electrodes. J Exp Bot. 1988;39:1541–1547. [Google Scholar]
- Felle HH. The apoplastic pH of the Zea mays root cortex as measured with pH-sensitive microelectrodes: aspects of regulation. J Exp Bot. 1998;49:987–995. [Google Scholar]
- Föhr KJ, Wardiol W, Gratzl M. Calculation and control of free divalent cations in solutions used for membrane fusion studies. Methods Enzymol. 1993;221:149–157. doi: 10.1016/0076-6879(93)21014-y. [DOI] [PubMed] [Google Scholar]
- Giromini L, Camattari G, Cerana R, Colombo R. Effect of apoplastic and cytoplasmic pH on inward and outward rectifying plasma membrane K+ channels of Arabidopsis thaliana cultured cells. J Plant Physiol. 1997;151:682–688. [Google Scholar]
- Grignon C, Sentenac H. pH and ionic conditions in the apoplast. Annu Rev Plant Physiol. 1991;42:103–128. [Google Scholar]
- Hamill OP, Marty A, Neher E, Sakman B, Sigworth FJ. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflügers Arch. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- Hedrich R, Moran O, Conti F, Busch H, Becker D, Gambale F, Dreyer I, Küch A, Neuwinger K, Palme K. Inward rectifier potassium channels in plants differ from their animal counterparts in response to voltage and channel modulators. Eur Biophys J. 1995;24:107–115. doi: 10.1007/BF00211406. [DOI] [PubMed] [Google Scholar]
- Hirsch RE, Lewis BD, Spalding EP, Sussman MR. A role for the AKT1 potassium channel in plant nutrition. Science. 1998;280:918–921. doi: 10.1126/science.280.5365.918. [DOI] [PubMed] [Google Scholar]
- Hoffmann B, Kosegarten H. FITC-dextran for measuring apoplast pH and apoplastic pH gradients between various cell types in sunflower leaves. Physiol Plant. 1995;95:327–335. [Google Scholar]
- Hoth S, Dreyer I, Dietrich P, Becker D, Müller-Röber B, Hedrich R. Molecular basis of plant-specific acid activation of K+ uptake channels. Proc Natl Acad Sci USA. 1997;94:4806–4810. doi: 10.1073/pnas.94.9.4806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ilan N, Schwartz A, Moran N. External protons enhance the activity of the hyperpolarization-activated K channel in guard cell protoplasts of Vicia faba. J Membr Biol. 1996;154:169–181. doi: 10.1007/s002329900142. [DOI] [PubMed] [Google Scholar]
- Katsuhara M, Tazawa M. Mechanism of calcium-dependent salt tolerance in cells of Nitellopsis obtusa: role of intracellular adenine nucleotides. Plant Cell Environ. 1990;13:179–184. [Google Scholar]
- Ketchum KA, Slayman CW. Isolation of a ion channel gene from Arabidopsis thaliana using the H5 signature sequence from voltage-dependent K+ channels. FEBS Lett. 1996;378:19–26. doi: 10.1016/0014-5793(95)01417-9. [DOI] [PubMed] [Google Scholar]
- Kochian LV, Lucas WJ. Potassium-transport in roots. Adv Bot Res. 1988;15:93–178. [Google Scholar]
- Kourie JI, Findlay GP. Ionic currents across the plasmalemma of Chara inflata cells. II. Effects of external Na+, Ca2+ and Cl− on K+ and Cl− currents. J Exp Bot. 1990;41:151–163. [Google Scholar]
- Kutschera U. The current status of the acid-growth hypothesis. New Phytol. 1994;126:549–569. [Google Scholar]
- Lagarde D, Basset M, Lepetit M, Conejero G, Gaymard F, Astruc S, Grignon C. Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J. 1996;9:195–203. doi: 10.1046/j.1365-313x.1996.09020195.x. [DOI] [PubMed] [Google Scholar]
- Maathuis FJM, Ichida AM, Sanders D, Schroeder JI. Roles of higher plant K+ channels. Plant Physiol. 1997;114:1141–1149. doi: 10.1104/pp.114.4.1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maathuis FJM, Sanders D. Mechanism of high-affinity potassium uptake in roots of Arabidopsis thaliana. Proc Natl Acad Sci USA. 1994;91:9272–9276. doi: 10.1073/pnas.91.20.9272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marten I, Hoth S, Ketchum K, Hedrich R, Hoshi T (1998) The Arabidopsis K+ transporter AKT3 displays unique channel properties. Presented at the 11th International Workshop on Plant Membrane Biology, 1998, Cambridge, UK
- Mühling KH, Plieth C, Hansen UP, Sattelmacher B. Apoplastic pH of intact leaves of Vicia faba as influenced by light. J Exp Bot. 1995;46:377–382. [Google Scholar]
- Müller-Röber B, Ellenberg J, Provart N, Willmitzer L, Busch H, Becker D, Dietrich P, Hoth S, Hedrich R. Cloning and electrophysiological analysis of KST1, an inward rectifying K+ channel expressed in potato guard cells. EMBO J. 1995;14:2409–2416. doi: 10.1002/j.1460-2075.1995.tb07238.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu X, Narasimhan ML, Salzman RA, Bressan RA, Hasegawa PM. NaCl regulation of plasma membrane H+-ATPase gene expression in a glycophyte and a halophyte. Plant Physiol. 1993;103:713–718. doi: 10.1104/pp.103.3.713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmgren MG. Proton gradients and plant growth: role of the plasma membrane H+-ATPase. Adv Bot Res. 1998;28:1–70. [Google Scholar]
- Roelfsema MRG, Prins HBA. The membrane potential of Arabidopsis thaliana guard cells; depolarizations induced by apoplastic acidification. Planta. 1998;205:100–112. doi: 10.1007/s004250050301. [DOI] [PubMed] [Google Scholar]
- Sabirov RZ, Okada Y, Oiki S. Two-sided action of protons on an inward rectifier K+ channel (IRK1) Pflügers Arch. 1997;433:428–434. doi: 10.1007/s004240050296. [DOI] [PubMed] [Google Scholar]
- Taylor DP, Slattery J, Leopold AC. Apoplastic pH in corn root gravitropism: a laser scanning confocal microscopy measurement. Physiol Plant. 1996;97:35–38. [PubMed] [Google Scholar]
- Véry A-A, Gaymard F, Bosseux C, Sentenac H, Thibaud J-B. Expression of a cloned plant K+ channel in Xenopus oocytes: analysis of macroscopic currents. Plant J. 1995;7:321–332. doi: 10.1046/j.1365-313x.1995.7020321.x. [DOI] [PubMed] [Google Scholar]
- Wegner LH, De Boer AH, Raschke K. Properties of the K+ inward rectifier in the plasma membrane of xylem parenchyma cells from barley roots: effects of TEA+, Ca2+, Ba2+ and La3+ J Membr Biol. 1994;142:363–379. doi: 10.1007/BF00233442. [DOI] [PubMed] [Google Scholar]
- Wegner LH, Raschke K. Ion channels in the xylem parenchyma of barley roots. A procedure to isolate protoplasts from this tissue and a patch-clamp exploration of salt passageways into xylem vessels. Plant Physiol. 1994;105:799–813. doi: 10.1104/pp.105.3.799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitaker JE, Haugland RP, Ryan D, Hewitt PC, Prendergast FG. Fluorescent rhodol derivatives: versatile, photostable labels and tracers. Anal Biochem. 1992;207:267–279. doi: 10.1016/0003-2697(92)90011-u. [DOI] [PubMed] [Google Scholar]
- Wu J, Seliskar DM. Salinity adaptation of plasma membrane H+-ATPase in the salt marsh plant Spartina patens: ATP hydrolysis and enzyme kinetics. J Exp Bot. 1998;49:1005–1013. [Google Scholar]
- Zimmermann S, Talke I, Ehrhardt T, Nast G, Müller-Röber B. Characterization of SKT1, an inwardly rectifying potassium channel from potato, by heterologous expression in insect cells. Plant Physiol. 1998;116:879–890. doi: 10.1104/pp.116.3.879. [DOI] [PMC free article] [PubMed] [Google Scholar]