

Abstract
Ca2+-dependent protein kinases (CDPKs) containing a calmodulin-like domain integrated in their primary sequence are present primarily in plants. A member of this family was characterized from the groundnut (Arachis hypogea) plant and called GnCDPK (M. DasGupta [1994] Plant Physiol 104: 961–969). GnCDPK specifically uses the myosin light chain synthetic peptide (MLCpep), which is the phosphate-accepting domain of smooth muscle myosin light chains (KKRPQRATSNVFS), as an exogenous substrate under in vitro experimental conditions. In this report we show that GnCDPK undergoes intramolecular autophosphorylation. This self-phosphorylation occurs in threonine residues in a Ca2+-dependent (K0.5 = 0.5 μm) and calmodulin-independent manner. The kinase activity toward MLCpep and its sensitivity to Ca2+ were unaffected by prior autophosphorylation when measured under saturating ATP concentrations. The role of autophosphorylation in the exogenous substrate MLCpep phosphorylation reaction was reinvestigated at low ATP concentrations. A pronounced lag time of 1 to 2 min, followed by a linear increase of activity for 7.5 min, was seen in the initial rate of MLCpep phosphorylation under such suboptimal conditions. Prior autophosphorylation completely abolished this lag phase, and a sharp rise of exogenous substrate phosphorylation was seen from the 1st min. Our results suggest that autophosphorylation is a prerequisite for the activation of GnCDPK.
CDPKs act as sensors for intracellular Ca2+ fluxes and translate them into physiological responses by reversibly phosphorylating Ser and/or Thr residues of relevant essential enzymes (Roberts and Harmon, 1992). Plants were shown to contain a novel group of CDPKs that are regulated by direct binding of Ca2+ and are independent of CaM or phospholipids (Harmon et al., 1987; Battey and Venis, 1988). Sequence analysis revealed that the unique biochemistry of these kinases results from a novel structural arrangement within a single polypeptide, wherein the catalytic domain is joined to a CaM-like regulatory domain containing four Ca2+-binding EF hands through a junction domain (Harper et al., 1991). Almost 40 different CDPK-like sequences are now available in the database, and some have significantly degenerate sequences in the CaM-like domain. Some of these CDPK-related protein kinases do not require Ca2+ as an activator (Furumoto et al., 1996). These kinases, possibly with variable Ca2+-binding properties, are thus capable of translating quantitative Ca2+ signals to qualitative messages. Because Ca2+ is involved in many of the cellular functions of higher plants (Hepler and Wayne, 1985; Trewavas, 1986), it is reasonable to believe that novel Ca2+-mediated regulatory pathways operate in plant signal transduction through these kinases. The upstream or downstream events of the CDPKs in such pathways and the details of their mechanisms of regulation are not yet known.
Autophosphorylation, a common regulatory property of protein kinases, leads to changes in their activity and/or changes in dependence on their activators. The kinases become independent of their respective effectors by autophosphorylation of a pseudosubstrate sequence, whereby the inhibitory interaction of the domain is prevented (Schworer et al., 1988; DasGupta and Blumenthal, 1995). Two independent studies (Harmon et al., 1994; Harper et al., 1994) also demonstrated pseudosubstrate-type autoinhibitory properties of the junction domain of CDPKs. Because the autophosphorylation properties of CDPKs are not known in detail, studies in the direction of understanding the relationship between pseudosubstrate-type autoinhibition and autophosphorylation of CDPK-type kinases are yet to be undertaken.
The CDPK from groundnut (Arachis hypogea), a unique member of the CDPK family, was identified and characterized from dry groundnut seeds and was named GnCDPK (DasGupta, 1994). This enzyme specifically phosphorylates the phosphate-accepting domain of chicken smooth-muscle myosin light chains. GnCDPK has been purified to homogeneity and is composed of a single subunit with a native Mr of 53,000. As with several other CDPKs (Roberts and Harmon, 1992), GnCDPK was found to autophosphorylate in a Ca2+-dependent manner. The purpose of this investigation was to study the autophosphorylation phenomenon of GnCDPK in detail and to determine the role (if any) of autophosphorylation in modulating the activity of GnCDPK toward its exogenous substrate, MLCpep. Our results show that prior autophosphorylation of GnCDPK completely abolishes the characteristic lag observed during its exogenous substrate phosphorylation reaction. We believe that autophosphorylation is an activation step for GnCDPK.
MATERIALS AND METHODS
Materials
Groundnut (Arachis hypogea var JL24) seeds were obtained from the National Research Centre for Groundnuts (Gujrat, India). [γ-32P]ATP (specific activity 3000 Ci/mmol) was obtained from the Bhaba Atomic Research Centre (Bombay, India). Q Sepharose and Sephacryl 300 were purchased from Pharmacia. Blue Sepharose was prepared according to the method of Botime et al. (1972). W7, W5, calmidazolium, and cellulose plates were obtained from Sigma. All other reagents of analytical grade were obtained from Sisco Research Laboratories (Bombay, India).
Protein Purification
GnCDPK was purified as described previously (DasGupta, 1994). The final enzyme preparation was further clarified through a preparative polyacrylamide gel (Laemmli, 1970) in a slab-gel apparatus in the absence of SDS (nondenaturing conditions). Two-millimeter sections were cut from the gel, crushed, and left in 1 mL of 20 mm Tris-HCl, pH 7.4, at 4°C overnight. The centrifuged supernatants were concentrated using an ultrafiltration unit (Centricon 30, Amicon, Beverly, MA). The highest active fraction showed a single band in 10% SDS-PAGE and was used for all of the experiments described. Protein estimation was according to the method of Bradford (1976).
Kinase Assays
GnCDPK activity was determined by phosphate incorporation into MLCpep using the P81 filter-binding method as described by DasGupta et al. (1989) with the modifications described by DasGupta (1994). The final reaction mixture (25 μL) contained 50 mm Tris-HCl, pH 7.4, 150 μm substrate peptide, 10 mm MgOAc, 200 μm CaCl2, 250 μm [γ32-P]ATP (300 cpm/pmol), and 0.25 μg of enzyme. The reaction was carried out for 10 min at 25°C. Autophosphorylation was performed under the same conditions except that the [γ32-P]ATP concentration was 1 μm (30,000 cpm/pmol) and MLCpep was omitted. Reactions were terminated at the indicated times by the addition of SDS-PAGE sample buffer. Samples were subjected to 10% or 17.5% SDS-PAGE, and the gels were stained for 30 min and destained for 1 h before autoradiography. The phosphorylated protein was excised and the incorporated 32P was determined by liquid scintillation counting. For Ca2+-dependence curves, free Ca2+ levels were set using Ca2+/EGTA buffers with the stability constants described by Martell and Smith (1974).
The Effect of Autophosphorylation of GnCDPK on MLCpep Phosphorylation
The effect of autophosphorylation on MLCpep phosphorylation activity was measured by a two-step assay. In the first step (preincubation) the autophosphorylation reaction was carried out with GnCDPK (0.25 μg/assay volume) in a 15-μL reaction mixture as described above except that nonradioactive 1 μm ATP was used. After 10 min the entire reaction mixture was added to a cocktail for MLCpep phosphorylation, so that the final reaction mixture (25 μL) contained 50 mm Tris-HCl, pH 7.4, 150 μm substrate peptide, 10 mm MgOAc, 200 μm CaCl2, and 250 μm or 5 μm [γ32-P]ATP (300 or 30,000 cpm/pmol). The reaction was monitored for 7.5 min at 25°C. In control experiments GnCDPK was preincubated under identical conditions but without 1 μm nonradioactive ATP.
Phosphoamino Acid Analysis
Purified GnCDPK was autophosphorylated under standard conditions. Free ATP was removed by gel filtration through Sephadex G25 (Pharmacia). The eluted protein was extensively dialyzed, and the filtrate was evaporated in a freeze drier (Speed-Vac, Savant Instruments, Holbrook, NY) and subjected to hydrolysis in 6 n HCl for 2 h at 110°C. Hydrolysates were mixed with standard phosphoserine, phosphothreonine, and phosphotyrosine and analyzed by thin-layer electrophoresis on cellulose plates at pH 3.5 (acetic acid:pyridine:water, 50:5:945 [v,v]) at 800 V for 45 min (Cooper et al., 1983). Radiolabeled amino acids were identified by autoradiography, and the standards were identified by ninhydrin staining.
RESULTS
Autophosphorylation of GnCDPK
Pure GnCDPK was found to autophosphorylate its own sequence (Fig. 1A, lane a). This self-catalyzed phosphate incorporation was inhibited in the presence of 1 mm EGTA (Fig. 1A, lane b), indicating that the reaction occurs in a Ca2+-dependent manner. The concentration of Ca2+ needed for half-maximal autophosphorylation of GnCDPK was 0.5 μm (Fig. 1B), which is similar to the Ca2+ requirement for exogenous substrate phosphorylation by GnCDPK (Table I). The Mg2+ ion requirement of 5 to 10 mm was the same for both catalytic activities of GnCDPK (Table I). However, a wide difference was noted in the affinity of GnCDPK toward ATP, depending on whether it used its own structure or an exogenous substrate as its target. There was about a 100-fold difference in the enzyme's Km for ATP in autophosphorylation (100 nm) in the absence of substrate compared with the Km for ATP in the exogenous substrate phosphorylation (10 μm) reaction (Table I). Such differences were also noted in phosphorylase kinase, in which the Km value of ATP for autophosphorylation was found to be 27 μm compared with 240 μm for kinase-dependent phosphorylase phosphorylation (Hallenbeck and Walsh, 1983).
Figure 1.

Ca2+-dependent autophosphorylation of GnCDPK. A, Purified GnCDPK was autophosphorylated as described in Methods. The standard reaction mixture (25 μL) contained 50 mm Tris-HCl, pH 7.4, 10 mm MgOAc, 200 μm CaCl2, 1 μm [γ32-P]ATP (30,000 cpm/pmol), and 0.25 μg of enzyme. Reactions were carried out for 10 min at 25°C and terminated by the addition of SDS-PAGE sample buffer. Samples were analyzed in 10% polyacrylamide gels and autoradiographed. Lane a, Standard reaction mixture; lane b, plus 1 mm EGTA. B, Autophosphorylation of GnCDPK was monitored in different concentrations of free Ca2+ by using Ca2+/EGTA buffers in otherwise standard reaction mixtures. Catalytic rates are expressed relative to the reaction rate under standard assay conditions.
Table I.
Autophosphorylation and MLCpep phosphorylation by GnCDPK
| Parameter | Autophosphorylation | MLCpep Phosphorylation |
|---|---|---|
| K0.5 for Ca2+ | 0.5 μm | 0.5 μm |
| Mg2+ for maximum activity | 5–10 mm | 5–10 mm |
| Km for ATP | 100 nm | 10 μm |
Assays were according to procedures described in Methods. Values represent the means of triplicate assays.
Effect of CaM and CaM Antagonists on GnCDPK Activity
The nature of the Ca2+/CaM dependence in the autophosphorylation reaction and in the exogenous substrate phosphorylation reaction of GnCDPK was found to be the same. In a previous report (DasGupta, 1994) it was shown that exogenous CaM did not have an effect on GnCDPK activity, but that CaM-binding drugs such as W7 and R2457 (calmidazolium) inhibited GnCDPK activity with an IC50 of 30 and 10 μm, respectively. Here we show that the addition of exogenous CaM (10 μm) did not have any effect on autophosphorylation or on GnCDPK-catalyzed MLCpep phosphorylation (Fig. 2, lane b). However, in the presence of 50 μm R2457 (calmidazolium) or W7, both autophosphorylation and exogenous substrate phosphorylation by GnCDPK were completely inhibited (Fig. 2, lanes c–d). Under the same conditions W5, a much less potent analog of W7, had no inhibitory effect on GnCDPK activity (Fig. 2, lane e). Considering the proper order of potency of these drugs and the identical sensitivity of both of the catalytic functions of GnCDPK toward these drugs, the mechanism of Ca2+ dependence of both autophosphorylation and MLCpep phosphorylation reactions catalyzed by GnCDPK appears to be mediated through a CaM-like sequence integrated into its primary structure.
Figure 2.

Effect of CaM and CaM antagonists on autophosphorylation of GnCDPK and phosphorylation of MLCpep. Purified GnCDPK was incubated in standard reaction mixtures as described in Methods. The final reaction mixture (25 μL) contained 50 mm Tris-HCl, pH 7.4, 150 μm substrate peptide, 10 mm MgOAc, 200 μm CaCl2, 5 μm [γ32-P]ATP (30,000 cpm/pmol), 0.25 μg of enzyme, and the indicated effectors. Reactions were carried out for 10 min at 25°C and terminated by the addition of SDS-PAGE sample buffer. Samples were analyzed in 17.5% polyacrylamide gels and autoradiographed. Lane a, Standard reaction mixture; lane b, plus 10 μm CaM; lane c, plus 50 μm R2457; lane d, plus 50 μm W7; and lane e, plus 50 μm W5.
Mechanism of Autophosphorylation
Autocatalysis of GnCDPK could occur by two potential reaction mechanisms, intermolecular and intramoleular. To determine by which mechanism autophosphorylation occurs, the dependency of the rate of autophosphorylation was examined as a function of the GnCDPK concentration. The reaction rate was linearly dependent on the GnCDPK concentration over a 20-fold (25–500 nm) range of GnCDPK concentrations, which is diagnostic of an intramolecular reaction (Fig. 3A, lanes a–d). Support for this model was also provided by analysis of the relationship between the reaction rate and the enzyme concentration using Van't Hoff's plot, which had a slope of 1.042 (Fig. 3B).
Figure 3.

Effect of enzyme concentration on autophosphorylation. A, Purified GnCDPK was successively diluted to the indicated concentrations and autophosphorylated as described in Methods for 1 min. The standard reaction mixture (25 μL) contained 50 mm Tris-HCl, pH 7.4, 10 mm MgOAc, 200 μm CaCl2, 1 μm [γ32-P]ATP (30,000 cpm/pmol), and enzyme in varying amounts. Lane a, 25 nm (0.03 μg/25 μL); lane b, 50 nm (0.06 μg/25 μL); lane c, 250 nm (0.3 μg/25 μL); and lane d, 500 nm (0.6 μg/25 μL). Reactions were terminated by the addition of SDS-PAGE sample buffer, and samples were analyzed in 10% polyacrylamide gels and autoradiographed. The radioactivity associated with GnCDPK bands was quantitated by scintillation counting and the specific activity was plotted against the enzyme concentration. B, Van't Hoff plot of log velocity versus log enzyme concentration using the data in A.
The time course of the autophosphorylation reaction is shown in Figure 4. Saturation of this self-catalyzed phosphate incorporation was attained within 2 min under our experimental conditions. The level of phosphorylation remained the same thereafter (Fig. 4, inset). This level of phosphorylation was found not to change even after 24 h (data not shown). To determine the stoichiometry of phosphate incorporation, the gel bands shown in the inset of Figure 4 were counted for radioactivity, as described in Methods. The stoichiometry of phosphate incorporation in GnCDPK was 0.2 mol/mol, which remained unchanged even after prolonged incubation with ATP (Fig. 4). Therefore, under the prescribed experimental conditions, at most only about 20% of the kinase could have been phosphorylated to the extent of 1 mol/mol at the point of saturation.
Figure 4.

Time course and stoichiometry of autophosphorylation of GnCDPK. Purified GnCDPK (0.25 μg) was autophosphorylated as described in Methods for the indicated time periods. The standard reaction mixture (25 μL) contained 50 mm Tris-HCl, pH 7.4, 10 mm MgOAc, 200 μm CaCl2, and 1 μm [γ32-P]ATP (30,000 cpm/pmol). Samples were analyzed in 10% SDS PAGE. The phosphorylated enzyme was visualized by autoradiography and the radioactivity was quantitated by scintillation counting. Inset, Autoradiogram of the analyzed samples.
The stoichiometry of phosphate incorporation into the autophosphorylation reaction was determined in four of our enzyme preparations and was consistently found to be in low order, indicating that low phosphate incorporation is not due to inactivation of the enzyme during purification. In another well-characterized CDPK from soybean, the stoichiometry of phosphate incorporation in the autophosphorylation reaction was also found to be approximately 0.2 mol/mol (Roberts and Harmon, 1992). The low stoichiometry of autophosphorylation in in vitro conditions could have been due to the absence of unknown factors that may determine optimum autophosphorylation in vivo. For example, in the case of phosphorylase kinase phosphorylation by cAMP-dependent protein kinase, the stoichiometry of phosphorylation was found to increase 28- to 36-fold by increasing the Mg2+ concentration (Hallenbeck and Walsh, 1983), indicating that reaction conditions have a profound influence on phosphorylation reactions.
An important difference was noted in the choice of the amino acid as the site of self-catalyzed phosphorylation compared with exogenous substrate phosphorylation. GnCDPK was found to be predominantly autophosphorylated in Thr residues, as shown in Figure 5. This is in contrast to what was observed in its exogenous substrate, MLCpep, in which the site of phosphorylation was preferably Ser (DasGupta, 1994).
Figure 5.
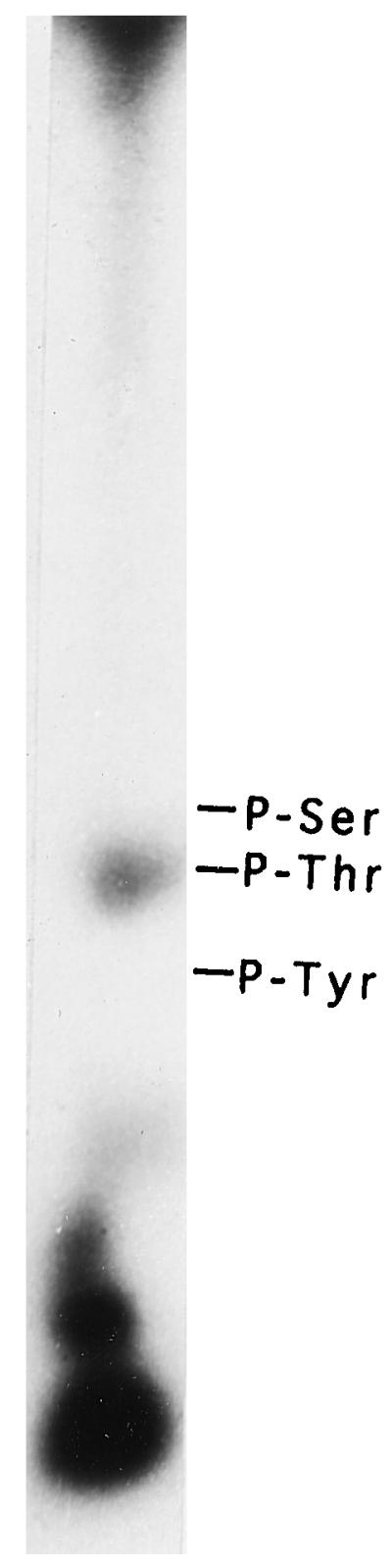
Identification of the phosphoamino acid residue in autophosphorylated GnCDPK. Autophosphorylation of GnCDPK was carried out as described in Methods. Phosphorylated GnCDPK was hydrolyzed in 6 n HCl, and the hydrolysate was dried, dissolved in water, and subjected to thin-layer electrophoresis at pH 3.5. The positions of the standards visualized by ninhydrin staining are indicated.
Effect of Autophosphorylation on GnCDPK Activity
For prior autophosphorylation, GnCDPK was preincubated in the presence of 1 μm ATP for 10 min, and this autophosphorylated enzyme was then used for MLCpep phosphorylation. Under saturating concentrations of ATP (250 μm) in a MLCpep phosphorylation reaction, autophosphorylation of GnCDPK was found to have no effect on the rate of its exogenous substrate phosphorylation activity. As shown in Figure 6A, the progress in MLCpep phosphorylation was identical in previously autophosphorylated and nonautophosphorylated GnCDPK. However, for Ca2+-dependent protein kinases, autophosphorylation is known to affect not only the enzymatic activity but also the response of the kinases to the metal ion. Such changes in the Ca2+ dependence of CaMKII are reported to range from a decrease in affinity for the metal ion to a formation of a Ca2+-independent form (Lai et al., 1986; Schworer et al., 1986). Autophosphorylation of GnCDPK was found to have no such effect on the Ca2+ dependence of the enzyme. As shown in Figure 6A, phosphorylation of MLCpep by both autophosphorylated GnCDPK and nonautophosphorylated GnCDPK was inhibited in the presence of EGTA, indicating that both reactions are Ca2+ dependent.
Figure 6.

Effect of autophosphorylation on MLCpep phosphorylation by GnCDPK. GnCDPK (0.25 μg/assay volume) was autophosphorylated in the presence of 50 mm Tris-HCl, pH 7.4, 10 mm MgOAc, 200 μm CaCl2, and 1 μm nonradioactive ATP for 10 min at 25°C as described in Methods. Control experiments were performed in which GnCDPK was preincubated under identical conditions except that water was substituted for ATP. GnCDPK-dependent MLCpep phosphorylation was then tested according to the procedure described in detail in Methods. The specific reaction conditions are indicated below. Maximal activity is defined as total counts per minute incorporated in the substrate under the indicated experimental conditions of assay at the 7.5-min time point. A, MLCpep phosphorylation assayed in the presence of 250 μm [γ32-P]ATP (300 cpm/pmol) for the indicated time periods with nonautophosphorylated GnCDPK (□), nonautophosphorylated GnCDPK and 1 mm EGTA (▵), autophosphorylated GnCDPK (▪), and autophosphorylated GnCDPK and 1 mm EGTA (▴). Values represent the means of triplicate assays. B, MLCpep phosphorylation assayed in the presence of 5 μm [γ32-P]ATP (300 cpm/pmol) in otherwise standard conditions for the indicated time periods with nonautophosphorylated GnCDPK (▪) or with autophosphorylated GnCDPK (□). Values represent the means of triplicate assays. C and D, MLCpep phosphorylation assayed in the presence of 5 μm [γ32-P]ATP (30,000 cpm/pmol) in otherwise standard conditions for 15 s (lanes a), 30 s (lanes b), 45 s (lanes c), 1 min (lanes d), 2.5 min (lanes e), 5 min (lanes f), or 7.5 min (lanes g) with nonautophosphorylated (C) or autophosphorylated (D) GnCDPK. The reaction mixtures were analyzed in 17.5% SDS PAGE and autoradiographed. The positions of GnCDPK and MLCpep are indicated.
These experiments for studying the effect of autophosphorylation of GnCDPK were then repeated in the presence of low ATP concentration (5 μm) during substrate phosphorylation to slow the initial rate of reaction. The reaction temperature (25°C) was kept unchanged. Interestingly, under such suboptimal conditions, without prior autophosphorylation of GnCDPK, there was a significant lag in the progress curve of MLCpep phosphorylation (Fig. 6B). When GnCDPK was incubated with 1 μm ATP for 10 min, this lag in MLCpep phosphorylation was completely abolished and there was a sharp increase in the reaction rate in the 1st min. The time of preincubation with ATP for autophosphorylation of GnCDPK in this experiment could be reduced to 2 min without significant loss of the stimulatory effect on the initial rates of MLCpep phosphorylation (data not shown).
The reaction mixtures described in Figure 6B were also analyzed in SDS-PAGE to monitor the specific labeling kinetics of the enzyme autophosphorylation and MLCpep phosphorylation, both with nonautophosphorylated enzyme (Fig. 6C) and with autophosphorylated enzyme (Fig. 6D). In the nonautophosphorylated enzyme the lag in MLCpep phosphorylation by GnCDPK could be seen clearly (Fig. 6C). Autophosphorylation under these conditions had no lag and progressed linearly until saturation was obtained after 2 min (Fig. 6C). The observed lag in MLCpep phosphorylation was completely abolished by prior autophosphorylation of the enzyme, as shown in Figure 6D. In this case, however, radiolabeled autophosphorylated GnCDPK bands were absent because of saturation of the autophosphorylation reaction in the preincubation period (Fig. 6D). This also indicates that the autophosphorylation reaction reaches a plateau by 2 min and therefore fulfills the structural change required to show optimum kinase activity. Preincubation of GnCDPK with a nonhydrolyzable ATP analog failed to show a similar effect on GnCDPK activity, indicating that it is not an ATP-dependent conformational change but autophosphorylation that is responsible for the modulation of the activity of GnCDPK toward its exogenous substrate (data not shown).
The next question concerns which aspect of GnCDPK that is changed by autophosphorylation leads to an increase in its activity toward MLCpep, as demonstrated under limiting conditions and shown in Figure 6B. For this answer we examined the affinities of pre-autophosphorylated and nonautophosphorylated GnCDPK for Ca2+ and MLCpep under the same limiting assay conditions (5 μm ATP), combined with short assay times (2 min). As shown in Table II, the K0.5 values for Ca2+ were the same in both pre-autophosphorylated and nonautophosphorylated GnCDPK, indicating that autophosphorylation does not change the sensitivity of the enzyme toward this divalent ion. Experiments performed under saturating conditions also indicated that autophosphorylation does not affect the Ca2+ dependence of the GnCDPK enzyme (Fig. 6A).
Table II.
MLCpep phosphorylation by preautophosphorylated and nonautophosphorylated GnCDPK
Under the same set of limiting conditions, the pre-autophosphorylated and the nonautophosphorylated GnCDPK differed widely in their affinities toward MLCpep, as reflected in the estimated Km values of 50 and 250 μm, respectively (Table II). This observation indicates that a conformational change in GnCDPK after autophosphorylation enables the enzyme to phosphorylate its exogenous substrate by increasing its affinity toward the exogenous substrate. We also examined the ATP dependence of GnCDPK using short assay times for both its autophosphorylated and nonautophosphorylated forms. The estimated Km values under such conditions were found to be 10 μm in both cases, indicating that autophosphorylation does not change the affinity of the enzyme for the nucleotide.
DISCUSSION
Autophosphorylation of several protein kinases was found to lead to changes in activity and/or changes in the dependence of the enzymes on their activators. For example, autophosphorylation of phosphorylase kinase caused an increase in enzymatic activity (King et al., 1983), whereas autophosphorylation of CaMKII rendered it autonomous of Ca2+ and CaM (Lai et al., 1986). However, autophosphorylation is not always necessarily linked to a modulation of kinase activity. It also regulates the subcellular distribution of protein kinases, promoting rapid and preferential modulation of specific targets within a defined microenvironment in response to diffusible second messengers. For example, CaMKII was released from the cytoskeleton after autophosphorylation (Saitoh and Schwartz, 1985). The phenomenon of autophosphorylation was demonstrated in many CDPKs (Roberts and Harmon, 1992) and shown to have an up- or down-regulatory effect on the associated kinase activity (Bogre et al., 1988; Saha and Singh, 1995). For wingbean CDPK, the autophosphorylation reaction was found to be independent of Ca2+, although the exogenous substrate phosphorylation was strictly dependent on Ca2+ (Saha and Singh, 1995). The present studies were undertaken to investigate autophosphorylation and its regulatory effect on GnCDPK.
Autophosphorylation-dependent up-regulation of CDPK activity was previously demonstrated in a heterogeneous preparation of CDPK from alfalfa, in which it was shown that incubation with unlabeled ATP before the addition of labeled ATP and exogenous substrate resulted in a 4-fold increase in rate but no difference in Ca2+ sensitivity (Bogre et al., 1988). However, for another well-characterized, pure CDPK from soybean, no such regulatory effect of autophosphorylation could be detected under comparable conditions (Roberts and Harmon, 1992). Similar experiments with pure GnCDPK also indicated that the rate of phosphorylation of MLCpep by GnCDPK, as well as its Ca2+ sensitivity, remained unchanged by prior autophosphorylation of the pure enzyme (Fig. 6A).
In a similar set of experiments in which the MLCpep phosphorylation reaction by GnCDPK was performed under suboptimal concentrations of ATP, interesting deviations were observed. With nonautophosphorylated GnCDPK under such conditions, a significant lag in MLCpep phosphorylation was noted (Fig. 6B), suggesting that autophosphorylation and the substrate phosphorylation of GnCDPK may not be a concomitant phenomenon. It is possible that autophosphorylation precedes substrate phosphorylation and is a prerequisite needed by GnCDPK to show its exogenous substrate phosphorylation. This proposition is supported by the experiments described in Figure 6, B and D, where prior autophosphorylation completely abolished the lag in MLCpep phosphorylation by GnCDPK.
As shown in Figure 6, B and D, there was a sharp increase in the initial rate of MLCpep phosphorylation with autophosphorylated GnCDPK. The lag period in the nonautophosphorylated GnCDPK is due to the autophosphorylation reaction and the subsequent change of conformation that enables the enzyme to interact with its exogenous substrate, MLCpep, with increasing affinity (Table II) and to phosphorylate it. However, the maximal rate of MLCpep phosphorylation attained by the nonautophosphorylated enzyme after overcoming the lag period was only one-fourth of the maximal rate of MLCpep phosphorylation by the preautophosphorylated enzyme (Fig. 6B), even though the autophosphorylation reaction in the presence of MLCpep is saturated within 2.5 min (Fig. 6C). A possible explanation may be that the prior presence of MLCpep in the vicinity of the enzyme is inhibitory toward the requisite structural change that follows in the enzyme after it is autophosphorylated.
In an earlier report on CDPK from alfalfa (Bogre et al., 1988), the difference in the labeling kinetics of the enzyme protein and its substrate, histone, was discussed without any presented data. Autophosphorylation of this alfalfa CDPK was reported to be saturated within 30 s, with an increased rate of histone phosphorylation thereafter. This is worth mentioning because it is similar to what was observed with GnCDPK. Another possibility that arises from these data is that autophosphorylation may be a manifestation of the formation of an enzyme-bound covalent intermediate during turnover of the phosphate to MLCpep. The large difference between the catalytic constants of the autophosphorylation reaction (0.9 nmol min−1 mg−1) and the exogenous substrate phosphorylation reaction (600 nmol min−1 mg−1) is a strong argument against such a proposition, however.
We propose a working model based on our observations, shown in Figure 7, of GnCDPK-dependent phosphorylation that occurs in two steps. In step I, the enzyme is autophosphorylated in a Thr residue in the presence of Ca2+ and undergoes a structural change. This structural change enables the enzyme to recognize and phosphorylate MLCpep in step II. Identity of the phosphorylation site associated with the transition of GnCDPK from step I to step II is not yet known. The present set of data cannot explain how the low extent of autophosphorylation of GnCDPK (0.2 mol/mol) could be responsible for the observed effects.
Figure 7.

Possible model for the role of autophosphorylation in the activation of GnCDPK. The shaded structure represents GnCDPK and the line structure represents MLCpep. Details of the model are discussed in the text.
One explanation, based on our preliminary observations (S. Chaudhuri and M. DasGupta, unpublished data), can be suggested. In all of our efforts to purify GnCDPK, 10% to 30% GnCDPK activity was found to be eluted in high-Mr fractions, which do not autophosphorylate. It is possible that a low extent of autophosphorylation of GnCDPK initiates cooperative interaction between the phosphorylated and nonphosphorylated enzyme, resulting in the formation of a multimeric active enzyme complex that is resistant to further autophosphorylation. This explanation accounts for and explains how 0.2 mol/mol self-catalyzed phosphate incorporation can be responsible for the activation of GnCDPK. Such a cooperative model for phosphorylation and activation was suggested to take place in CaMKs by Kwiatkowski et al. (1988). Autophosphorylation of a recently characterized atypical Ca2+/CaM-dependent protein kinase was found to occur in a stoichiometry of 0.339 mol phosphate mol−1 kinase (Patil et al., 1995; Takezawa et al., 1996). The low level of autophosphorylation of this kinase also leads to stimulation of its kinase activity. Why certain kinases tend to have a very low level of self-catalyzed phosphorylation and how such low levels of phosphorylation enable them to effect their activity toward exogenous substrates needs more investigation.
ACKNOWLEDGMENT
We thank Dr. D.K. Blumenthal for generously providing the MLCpep.
Abbreviations:
- CaM
calmodulin
- CaMKII
type II CaM-dependent protein kinase
- CDPK
Ca2+-dependent protein kinase
- MLCpep
myosin light-chain synthetic peptide
- R2457
(1-[bis-(4-chlorophenyl)methyl]-3-[2-(2,4-dichlorophenyl)-2-[(2,4-dichloro-phenyl)methoxy]-ethyl]-1H-imidazolium chloride)
- W5
N-(6-aminohexyl)-1-naphthalene sulfonamide
- W7
N-(6-aminohexyl)-5-chloro-1-naphthalene sulfonamide
Footnotes
This work was supported by the Council of Scientific and Industrial Research (grant no. 38-0894-95-EMR II), Government of India.
LITERATURE CITED
- Battey NH, Venis MA. Calcium-dependent protein kinase from apple fruit membranes is calmodulin-independent but has calmodulin-like properties. Planta. 1988;176:91–97. doi: 10.1007/BF00392484. [DOI] [PubMed] [Google Scholar]
- Bogre L, Olah Z, Dudits D. Calcium dependent protein kinase from alfalfa (Medicago varia): partial purification and autophosphorylation. Plant Sci. 1988;58:135–144. [Google Scholar]
- Botime HJ, Kopperschlager G, Schulz J. Affinity chromatography of phosphofructokinase using Cibacron blue F3G-A. J Chromatogr. 1972;69:209–214. doi: 10.1016/s0021-9673(00)83103-9. [DOI] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for quantitation of microgram quantities of proteins utilising the principle of protein dye binding. Anal Biochem. 1976;170:116–122. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Cooper JA, Bartholomew MS, Hunter T. Detection and quantification of phosphotyrosine in proteins. Methods Enzymol. 1983;99:387–405. doi: 10.1016/0076-6879(83)99075-4. [DOI] [PubMed] [Google Scholar]
- DasGupta M. Characterization of a calcium dependent protein kinase from groundnut (Arachis hypogea) seeds. Plant Physiol. 1994;104:961–969. doi: 10.1104/pp.104.3.961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DasGupta M, Blumenthal DK. Characterisation of the regulatory domain of the γ-subunit of phosphorylase kinase. J Biol Chem. 1995;270:22283–22289. doi: 10.1074/jbc.270.38.22283. [DOI] [PubMed] [Google Scholar]
- DasGupta M, Honeycutt T, Blumenthal DK. The γ-subunit of skeletal muscle phosphorylase kinase contains two non-contiguous domains that act in concert to bind calmodulin. J Biol Chem. 1989;264:17156–17163. [PubMed] [Google Scholar]
- Furumoto T, Ogawa N, Hata S, Izui K. Plant calcium dependent protein kinase-related kinases (CRKs) do not require calcium for their activities. FEBS Lett. 1996;396:147–151. doi: 10.1016/0014-5793(96)01090-3. [DOI] [PubMed] [Google Scholar]
- Hallenbeck PC, Walsh DA. Autophosphorylation of phosphorylase kinase: divalent metal ion and nucleotide dependency. J Biol Chem. 1983;258:13493–13501. [PubMed] [Google Scholar]
- Harmon AC, Putnam-Evans C, Cormier MJ. A calcium dependent but calmodulin-independent protein kinase from soybean. Plant Physiol. 1987;83:830–837. doi: 10.1104/pp.83.4.830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harmon AC, Yoo BC, McCaffery C. Pseudosubstrate inhibition of CDPK, a protein kinase with a calmodulin-like domain. Biochemistry. 1994;33:7278–7283. doi: 10.1021/bi00189a032. [DOI] [PubMed] [Google Scholar]
- Harper JF, Hung JF, Lloyd SJ. Genetic identification of an autoinhibitor in CDPK, a protein kinase with a calmodulin-like domain. Biochemistry. 1994;33:7267–7277. doi: 10.1021/bi00189a031. [DOI] [PubMed] [Google Scholar]
- Harper JF, Sussman MR, Scheller EG, Putnam-Evans C, Charbonneau H, Harmon AC. A calcium dependent protein kinase with a regulatory domain similar to calmodulin. Science. 1991;252:951–954. doi: 10.1126/science.1852075. [DOI] [PubMed] [Google Scholar]
- Hepler PK, Wayne RO. Calcium and plant development. Annu Rev Plant Physiol. 1985;36:397–439. [Google Scholar]
- King MM, Fitzgerald TJ, Carlson GM. Characterisation of initial autophosphorylation events in rabbit skeletal muscle phosphorylase kinase. J Biol Chem. 1983;258:9925–9930. [PubMed] [Google Scholar]
- Kwiatkowski AP, Shell DJ, King MM. The role of autophosphorylation in activation of the type II calmodulin-dependent protein kinase. J Biol Chem. 1988;263:6484–6486. [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;277:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Lai Y, Nairn AC, Greengard P. Autophosphorylation reversibly regulates the Ca2+/calmodulin dependence of Ca2+/calmodulin-dependent protein kinase II. Proc Natl Acad Sci USA. 1986;83:4253–4257. doi: 10.1073/pnas.83.12.4253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martell AE, Smith RM (1974) Critical Stability Constants. Plenum Press, New York, pp 269–271
- Patil S, Takezawa D, Poovaiah BW. A chimeric Ca2+/calmodulin-dependent protein kinase (CCaMK) gene characterized by a catalytic domain, a calmodulin-binding domain, and a neural visinin-like Ca2+-binding domain was recently cloned from plants. Proc Natl Acad Sci USA. 1995;92:4797–4801. doi: 10.1073/pnas.92.11.4897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts DM, Harmon AC. Calcium modulated proteins: targets of intracellular calcium signals in higher plants. Annu Rev Plant Physiol Plant Mol Biol. 1992;43:375–414. [Google Scholar]
- Saha P, Singh M. Characterisation of a winged bean (Psophocarpus tetragonolobus) protein kinase with a calmodulin-like domain: regulation by autophosphorylation. Biochem J. 1995;305:205–210. doi: 10.1042/bj3050205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saitoh T, Schwartz JH. Phosphorylation-dependent subcellular translocation of a Ca2+/calmodulin-dependent protein kinase produces an autonomous enzyme in Aplysia neurons. J Cell Biol. 1985;100:835–842. doi: 10.1083/jcb.100.3.835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schworer CM, Colbran RJ, Keefer JR, Soderling TR. Ca2+(calmodulin)-dependent protein kinase II: identification of a regulatory autophosphorylation site adjacent to the inhibitory and calmodulin-binding domains. J Biol Chem. 1988;263:13486–13489. [PubMed] [Google Scholar]
- Schworer CM, Colbran RJ, Soderling TR. Reversible generation of a Ca2+-independent form of Ca2+(calmodulin)-dependent protein kinase II by an autophosphorylation mechanism. J Biol Chem. 1986;261:8581–8584. [PubMed] [Google Scholar]
- Takezawa D, Ramachadiran S, Paranjape V, Poovaiah BW. Dual regulation of a chimeric plant serine/threonine kinase by calcium and calcium/calmodulin. J Biol Chem. 1996;271:8126–8132. doi: 10.1074/jbc.271.14.8126. [DOI] [PubMed] [Google Scholar]
- Trewavas AJ, ed (1986) Molecular and Cellular Aspects of Calcium in Plant Growth and Development. Plenum Press, New York