
FIGURE 6.
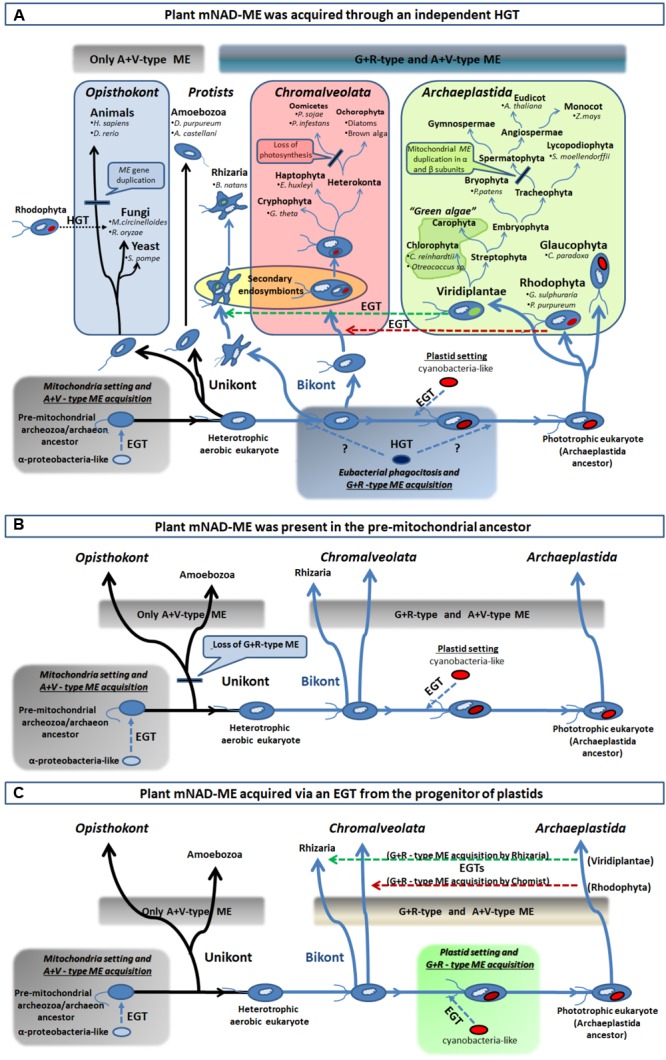
Different scenarios of acquisition of the R+G clade of Class I-ME in eukaryotes. (A) From bottom left, an extensive EGT from an engulfed α-proteobacteria-like ancestor (mitochondria precursor) to the nucleus of a unikont (one flagellum) host took place. This resulted in the establishment of eubacterial ME version in early eukaryotes and the foundation of the A+V clade of ME. After unikont and bikont (two flagella) splitting and prior – or post – plastid setting (indicated by question marks), an HGT event from one eubacteria (possibly a γ-proteobacteria-like) into the Archaeplastida ancestor established the G+R clade of Class I-MEs in eukaryotes. This allowed spreading of G+R isoforms in all primary photosynthetic organisms, as well as, in the secondary phototrophs (Chromalveolata and Rhizaria) via an EGT (green and red horizontal broken line arrows) and in some fungi by an HGT event (black horizontal dotted line arrow). (B) The G+R ME could be the ancestral isoform that was present in the pre-mitochondrial, and founder of the eukaryotic lineage, ancestor (the “Archezoa,” or an autotrophic methanogenic archaeon, depending on the model for the evolution of eukaryotes). The establishment of mitochondria in this ancestor allowed the acquisition of an additional isoform (the type A+V) and this chimeric structure for the MEF was maintained by the bikont lineage. However, only the A+V-type ME was retained by the unikont ancestor, as it lost the more ancestral isoform in what could have been a genetic replacement. (C) The G+R ME acquisition by photosynthetic organisms (containing the A+V ME derived from the mitochondria setting) may have taken place in with the plastid genesis, through the extensive EGT from the engulfed cyanobacteria to the host genome. Hence, the presence of this isoform in secondary phototrophs organisms results from secondary endosymbiosis processes, as is explained in A. For clarity, the major taxa of eukaryotic organisms were omitted in B and C.