

Abstract
Background
Invasive pulmonary aspergillosis (IPA) is a life-threatening disease in immunosuppressed patients. Voriconazole is commonly used to prevent and treat IPA in the clinic, but the optimal prophylactic antifungal regimen is unknown. The objective of this study was to clarify the mechanism underlying how voriconazole prevents IPA based on a target cellular pharmacokinetics/pharmacodynamics model, with the aim of identifying a way to design an optimal prophylactic antifungal regimen.
Methods
A nystatin assay was used to establish a target-cells model for A. fumigatus infection. An inhibitory effect sigmoid Emax model was developed to explore the cellular PK/PD breakpoint, and Monte Carlo simulation was used to design the prophylactic antifungal regimen.
Results
The intracellular activity of voriconazole in the target cells varied with its concentration, with the minimum inhibitory concentration (MIC) being an important determinant. For A. fumigatus strains AF293 and AF26, voriconazole decreased the intracellular inoculum by 0.79 and 0.84 lg cfu, respectively. The inhibitory effect sigmoid Emax model showed that 84.01% of the intracellular inoculum was suppressed by voriconazole within 24 h, and that a PK/PD value of 35.53 for the extracellular voriconazole concentration divided by MIC was associated with a 50% suppression of intracellular A. fumigatus. The Monte Carlo simulation results showed that the oral administration of at least 200 mg of voriconazole twice daily was yielded estimated the cumulative fraction of response value of 91.48%. Concentration of voriconazole in the pulmonary epithelial lining fluid and the plasma of > 17.77 and > 1.55 mg/L, respectively, would ensure the PK/PD > 35.53 for voriconazole against most isolates of A. fumigatus and may will be benefit to prevent IPA in clinical applications.
Conclusions
This study used a target cellular pharmacokinetics/pharmacodynamics model to reveal a potential mechanism underlying how voriconazole prevents IPA and has provided a method for designing voriconazole prophylactic antifungal regimen in immunosuppressed patients.
Keywords: Voriconazole, Invasive pulmonary aspergillosis, Prophylactic antifungal regimen, Monte Carlo simulation, Cellular pharmacodynamic/pharmacodynamic
Background
Invasive pulmonary aspergillosis (IPA) is mainly caused by A. fumigatus and has a high morbidity and mortality in high-risk immunosuppressed patients [1]. The incidence of IPA varies according to the underlying disease, being up to 24% in patients with acute leukemia, up to 10% in patients with allogeneic hematopoietic stem cell transplantation, and up to 7% in patients with lymphoid malignancies [2]. Additionally, the mortality rate of IPA is reportedly as high as 36–75% in patients with hematological malignancies despite the application of antifungal agents such as triazoles and echinocandins [3–5]. Thus, antifungal prophylaxis is currently recommended in high-risk immunosuppressed patients [6, 7].
Currently, no reported study has investigated how to develop an optimal antifungal prophylactic dosage regimen, and so the most common practice in clinical is to apply the recommended dosage regimen for prophylaxis. Theoretically, an understanding of the pathogenesis of A. fumigatus could provide clues for designing prophylactic antifungal regimen that are effective at preventing IPA. Inhalation is the primary route for acquiring Aspergillus spores. Pulmonary epithelial cells are the first cells encountered by the inhaled spores, which then germinate and grow in these cells, followed by the development of serious IPA disease in immunosuppressed patients [8]. Therefore, pulmonary epithelial cells can be viewed as the target of prophylaxis in IPA.
Voriconazole is a triazole exhibiting broad-spectrum antifungal activity against Aspergillus species [9], and is approved as the first-line therapy for invasive aspergillosis [6, 10]. The Infectious Diseases Society of America recommends using voriconazole for prophylaxis against invasive aspergillosis in high-risk patients, such as patients with prolonged neutropenia, patients with graft-versus-host disease, and lung transplant patients (strong recommendation and moderate-quality evidence) [6]. In addition, voriconazole is safe and effective for the secondary prophylaxis of systemic fungal infection in patients receiving allogeneic stem cell transplantation [11]. It has been reported that the concentration of voriconazole in the pulmonary epithelial lining fluid (ELF) crucially influences the prevention of IPA [12]. However, the ability of voriconazole penetrate the ELF varies widely in patients, with concentrations ranging from 0 to 83 mg/L being found in transplant patients receiving the recommended oral dosage regimen [13, 14]. While the ELF is on the surface of the pulmonary epithelial cells [15], the activity of voriconazole against A. fumigatus in these cells is unclear, which hinders the ability to design an optimal prophylactic dosage regimen for voriconazole in immunosuppressed patients.
We established a model of infected human pulmonary epithelial cells caused by A. fumigatus conidia with the following aims: (1) to determine the cellular pharmacokinetics/pharmacodynamics (PK/PD) characteristics of voriconazole in pulmonary epithelial cells, and (2) to identify a strategy for designing prophylactic regimen of voriconazole that are effective at preventing IPA.
Methods
Cell line, strains, and drug preparation
Human pulmonary epithelial cells (A549) were obtained from Shanghai Institute of Cell Biology in the Chinese Academy of Sciences (Shanghai, China). A549 cells were grown in vitro using 1640 medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin, respectively. A. fumigatus strain AF293 (from FGSC, Fungal Genetics Stock Center, School of Biological Sciences, University of Missouri, Kansas City, Missouri, USA) and clinical isolate strain (AF26, identified by department of laboratory, The First Affiliated Hospital of Xi’an Jiaotong University) were used for our study and grown on Sabouraud Dextrose Agar (SDA) medium at 35 °C for 5 days. For these two strains, conidia were harvested using phosphate-buffered saline plus 0.1% Tween 80. Voriconazole and nystatin were purchased from the national institute for Food and Drug control (Beijing, China) and dissolved in dimethyl sulfoxide.
Antifungal susceptibility testing
The E-test was performed according to the manufacturer’s instructions. The E-test strips (Biomeriux, USA) contained voriconazole at concentrations ranging from 0.002 to 32 mg/L. E-test strips were placed on SDA and the plates were incubated at 35 °C, and the minimum inhibitory concentration (MIC) was determined after an incubation time of 48 h. The MIC determined by the E-test was the lowest drug concentrations at which the border of the elliptical inhibition intercepted the scale on the antifungal strip.
Cellular PK/PD properties of voriconazole
Infected model of A549 cells
A549 cells were seeded at 5 × 105 cells/well in six-well plates and grown for 24 h. Following cell growth and washing by PBS, A549 cells and conidia were combined at a 1:5 ratio in six-well plates and incubated for 3 h at 37 °C. After incubation, the cells were washed three times with PBS and then treated with nystatin-supplemented media for a further 3 h to kill the conidia that adhered to the cells [16, 17]. Our previous study showed that only 1.81% of the viable initial A. fumigatus conidia were internalized by A549 cells, and that the infected-cells model could be used to evaluate the intracellular activity of voriconazole against A. fumigatus [17].
Biomarker of A549 cells and A. fumigatus
The viability of infected A549 cells in the presence of voriconazole was evaluated by measuring the cytosolic enzyme lactate dehydrogenase (LDH) released into the culture medium after 24 h of incubation using an LDH cytotoxicity assay kit (Beyotime Institute of Biotechnology, Haimen, Jiangsu, China). The growth of A. fumigatus was quantified by collecting culture supernatants and measuring the levels of galactomannan using the Platelia® Aspergillus assay (Bio-Rad, Marnes la Coquette, France) according to the manufacturer’s instructions.
Cellular pharmacodynamics of voriconazole
The intracellular activity was examined for extracellular voriconazole concentrations (Ce) ranging from 0.0155 to 64 mg/L, which is similar to the range of voriconazole ELF concentrations measured in transplant patients. After 24 h of incubation, the A549 cells were lysed with 1 mL of 0.5% Triton X-100, and the released conidia were diluted and then plated onto SDA to count the colony-forming units (cfu). The number of viable A. fumigatus conidia in each lysate sample was determined in duplicate incubations on SDA and using the standard plate-count method. The change in the number of cfu from that in the initial intracellular inoculum (at 0 h) was taken as the response to voriconazole and plotted as a function of the extracellular concentration (in logarithmic units). The measured data were fitted with a sigmoid model having a slope of − 1. The curve fittings were performed using GraphPad Prism software (version 5.0, GraphPad Software, San Diego, CA, USA). This sigmoid model allowed us to calculate the static concentration (Cs) corresponding to the extracellular concentration of voriconazole causing no apparent changes in cfu relative to the initial intracellular inoculum.
Inhibitory effect sigmoid Emax model
An inhibitory effect sigmoid Emax model (formula 1) was used to construct the relationship between Ce divided by MIC (the Ce/MIC ratio) and the intracellular antifungal effect at 24 h:
| 1 |
where E is the intracellular antifungal effect of voriconazole, expressed as a proportion of the initial intracellular inoculum, Emin is the effect for an infinitely low Ce/MIC value (this value is > 100%, and corresponds to the increase in cfu compared to the initial intracellular inoculum for an infinity low value of Ce/MIC). Emax is the maximum effect for an infinitely large Ce/MIC value (this value is < 100% and corresponds to the decrease in cfu compared to the initial intracellular inoculum for an infinitely high value of Ce/MIC), E50 is the extracellular concentration of voriconazole at which E is halfway between Emin and Emax, and H is the slope function. The intracellular pharmacodynamics data were modeled using WinNonlin software (version 4.1, Pharsight, Mountain View, CA, USA).
Monte Carlo simulation
To further explore the therapeutic implications of these experimental results, Monte Carlo simulation was used to evaluate the probability of different oral dosage regimens achieving the target value of Ce/MIC to design the appropriate prophylactic regimen of voriconazole in immunosuppressed patients. The MCS was performed by combining experimental results for the infected A549 cells with the data from a previous population pharmacokinetics study. The Monte Carlo simulation was based on
| 2 |
where the Ce/MIC break point of voriconazole comes from the inhibitory effect sigmoid Emax model; RELF/plasma is the ELF-to-plasma ratio of voriconazole concentration, and Voriconazole can penetrate well into the pulmonary ELF in patients, with a RELF/plasma of 11.5 ± 6.9 (mean ± SD) from published literatures [13, 14]; Voriconazole plasma concentrations and the variability of different dosage regimens in patients were simulated based on the previous population pharmacokinetics model using NONMEM software and twice daily oral dosage regimens of 100, 200, and 300 mg (under the maintenance dose) were simulated [18, 19]. The voriconazole ELF concentration was then calculated as the simulated voriconazole plasma concentration multiplied by RELF/plasma; the MIC distribution of A. fumigatus was obtained from the EUCAST (European Committee on Antimicrobial Susceptibility Testing) website.
The results of the Monte Carlo simulation were expressed as the probability of target attainment and the cumulative fraction of response [20]. A cumulative fraction of response value of ≥ 90% was considered to be an appropriate prophylactic regimen [21].
Statistical analysis
In the present study, numbers of cfu were converted to logarithms, base 10, for statistical analysis. All experimental values were expressed as the mean ± standard deviations of three independent experiment.
Results
Minimum inhibitory concentrations
The voriconazole MICs for A. fumigatus strains AF293 and AF26 were 0.19 and 0.064 mg/L, respectively.
Biomarker
We examined the viability of infected A549 cells for Ce values ranging from 0.0155 to 64 mg/L by measuring the LDH released into the medium. The amount released remained similar to the control values and lower than 11.5% over the whole range of concentrations investigated, which was therefore these levels considered when performing further experiments. Additionally, we found that the kinetics profile of galactomannan was correlated with Ce and that voriconazole induced a concentration-dependent decrease in the galactomannan index for both AF293 and AF26 (Fig. 1).
Fig. 1.
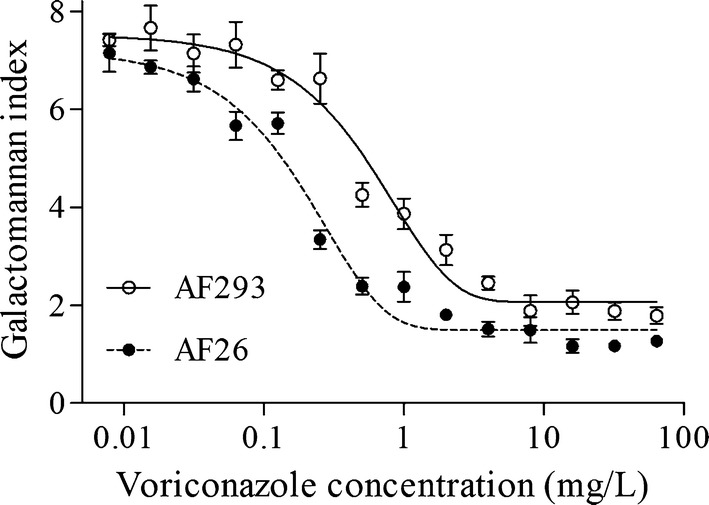
Concentration—galactomannan index relationships for voriconazole against two strains of A. fumigatus with differing MIC values in infected A549 model. The solid and dashed lines were fitted by the sigmoid function
Cellular pharmacodynamics of voriconazole against AF293 and AF26
The initial intracellular inoculum (at 0 h) was 1–2 × 105 cfu/mL per plate for the two strains investigated in this study. The voriconazole activity developed in a concentration-dependent manner, with a sigmoid function providing a satisfactory fit to the data for the two strains (Fig. 2a). The Emin values were similar for the two strains (approximate increases of 0.2 lg cfu), and reduced the intracellular inoculum by 0.79–0.84 lg cfu compared to the initial value at a high Ce (Emax). The number of intracellular A. fumigatus conidia was unchanged for the two strains compared to the initial intracellular inoculum at a Cs of 1.86 mg/L for AF293 and 0.52 mg/L for AF26, and E50 was 7.05 mg/L for AF293 and 2.11 mg/L for AF26 (Table 1). A particularly interestingly observation was that when Ce was expressed in multiples of the corresponding MIC (i.e., Ce/MIC ratio), the concentration–response curves of voriconazole against AF293 and AF26 became extraordinary close on the coordinate axis, which demonstrates the importance of MIC values to intracellular activity of voriconazole (Fig. 2b).
Fig. 2.

Goodness of fit fitted by sigmoidal function: extracellular concentration-dependent activity of voriconazole against intracellular A. fumigatus. The ordinate shows the change in the value of cfu (∆lg cfu) per mL of cell lysate after 24 h incubation compare with the initial intracellular conidia. a Data were plotted against the extracellular concentration (mg/L) of voriconazole. b Data were plotted against the ratio of extracellular concentration divide MIC value
Table 1.
The regression parameters and statistical analysis of the extracellular concentrations-activity curves illustrated in Fig. 2
| Strain | Emax (95%CI)a | Emin (95%CI)b | EC50 (95%CI)c | Cds | R2e |
|---|---|---|---|---|---|
| AF293 | − 0.79 (− 0.86 ~ − 0.72) lg cfu | 0.21 (0.18 ~ 0.24) lg cfu | 7.05 mg/L (5.36 ~ 9.27) 36.71 × MIC (27.91 ~ 48.30) |
1.86 mg/L (9.69 × MIC) |
0.9813 |
| AF26 | − 0.84 (− 0.89 ~ − 0.78) lg cfu | 0.20 (0.16 ~ 0.24) lg cfu | 2.11 mg/L (1.63 ~ 2.73) 32.93 × MIC (25.39 ~ 42.70) |
0.52 mg/L (8.32 × MIC) |
0.9828 |
acfu decrease (in lg units) at 24 h from the corresponding original cellular conidia, as extrapolated for infinitely high concentrations of voriconazole (note that a larger maximal relative activity corresponds to a more negative value of Emax)
bcfu increase (in lg units) at 24 h from the corresponding original cellular conidia, as extrapolated for infinitely low concentrations of voriconazole
cExposure (in extracellular concentration and multiple of corresponding MIC) causing a reduction halfway between the Emin and Emax values, as obtained from the hill equation by using a slope factor of − 1
dExposure (in extracellular concentration and multiple of corresponding MIC) resulting in no apparent pathogen growth (the number of cfu was identical to that of the original cellular conidia), as determined by graphical interpolation
eR is coefficient of correlation
Cellular PK/PD of voriconazole
All of the data could be fitted well by the inhibitory effect sigmoid Emax model, with a correlation coefficient of 0.97 and a slope of 1.10 (Fig. 3a). The model showed that progressively higher Ce/MIC values resulted in a progressive decrease in the number of A. fumigatus conidia in cells. Compared to the initial intracellular inoculum, the number of intracellular A. fumigatus conidia could be suppressed by 84.01% or increased by 59.36%, respectively, within 24 h, corresponding to the asymptotic maximum and minimum Ce/MIC values. Ce/MIC values of 8.93, 14.9, 35.53, and 311.30 resulted in 100, 80, 50, and 20% of the initial intracellular number of A. fumigatus conidia surviving in the A549 cells (Fig. 3b).
Fig. 3.

Relationship between the total voriconazole Ce/MIC and the intracellular antifungal effect as measured by the proportion of initial incubation. a The solid black line is the fit of an mathematical model to data from the infected A549 model, and the calculated equation by WinNonlin software is: effect index = 159.4 − 143.4 × (Ce/MIC)1.1/(12.21.1 + (Ce/MIC)1.1), R2 = 0.9738. b The relationship between Ce/MIC breakpoint and intracellular antifungal effect
Monte Carlo simulation results
Monte Carlo simulation was used to evaluate the probability of different dosage regimens achieving the target value of Ce/MIC = 35.53. Table 2 lists the probability of target attainments for specific MIC values ranging from 0.0625 to 16 mg/L for each dosage regimen. For the MIC of 0.125 mg/L, the values of probability of target attainments for voriconazole dosage regimens of 100, 200, and 300 mg administered twice daily via the oral route were 85.27, 92.94 and 98.03%, respectively. For A. fumigatus, voriconazole oral dosages of 100, 200, and 300 mg twice daily yielded estimated cumulative fraction of response values of 85.85, 91.48 and 96.19%, respectively (Table 2).
Table 2.
The probability of target attainment values at a certain minimum inhibitory concentration and the cumulative fraction of response values for A. fumigatus at different dosage regimens
| Dosage regimens | Probability of target attainment (%) at different minimum inhibitory concentrations | CFR (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.03125 mg/L | 0.0625 mg/L | 0.125 mg/L | 0.25 mg/L | 0.5 mg/L | 1 mg/L | 2 mg/L | 4 mg/L | 8 mg/L | 16 mg/L | ||
| 100 mg/12 h, p.o. | 99.25 | 96.35 | 85.27 | 63.9 | 36.39 | 14.92 | 4.14 | 0.72 | 0.099 | 0.006 | 84.45 |
| 200 mg/12 h, p.o. | 99.65 | 98.21 | 92.94 | 80.01 | 58.78 | 34.15 | 15.28 | 4.92 | 1.15 | 0.21 | 91.48 |
| 300 mg/12 h, p.o. | 99.98 | 99.67 | 98.03 | 92.25 | 77.55 | 54.15 | 29.07 | 11.35 | 3.07 | 0.61 | 96.19 |
CFR cumulative fraction of response
Discussion
Pulmonary epithelial cells invaded by A. fumigatus conidia in human body is the first step of causing IPA, and IPA is still related to high rates of morbidity and mortality after the utilization of antifungal agents [3–5]. Therefore, how to design the optimum prophylactic dosage regimen to prevent IPA is an intractable problem in clinical practice. Voriconazole has a strong activity against and it is vital to explore the intracellular activity of voriconazole against A. fumigatus in pulmonary epithelial cells. This study built the cells model infected by A. fumigatus to investigate, firstly, the PK/PD property of voriconazole in infected cells, and then use the Monte Carlo simulation and PK/PD breakpoint to design the prophylactic dosage regimen. This research provided a potential mechanism of voriconazole preventing IPA on a cellular level and provided a novel theoretical and experimental method for the designation of voriconazole prophylactic dosage regimen and the reduction of IPA morbidity.
Previous pharmacodynamics studies have used galactomannan (a major constituent of aspergillus cell walls) [22] as a biomarker for assessing the antifungal activity against A. fumigatus [23–25]. The present study found a distinct relationship between the voriconazole concentration and the galactomannan index for the two A. fumigatus strains (Fig. 1). In contrast to previous studies, [23–25] we did not calculate the pharmacodynamics parameters based on the relationship between the voriconazole concentration and the galactomannan index. One of the limitations in semiquantitative assessment is that the content of galactomannan in the medium cannot precisely reflect the number of A. fumigatus conidia in cells. Thus, the number of A. fumigatus conidia in A549 cells was determined in this study by performing quantitative culturing in a subsequent step.
A. fumigatus conidia in A549 cells were exposed for 24 h to voriconazole over a wide range of concentrations to determine the overall pattern of intracellular activity. The data obtained in this study of voriconazole cellular pharmacodynamics properties have not been reported previously in detail. The Emax values of voriconazole for the AF293 and AF26 strains were 0.79 and 0.84 lg cfu, respectively, showing a decrease compared with the initial intracellular inoculum (Table 1), which meant that 78.20 and 85.57% of intracellular A. fumigatus conidia were suppressed within 24 h for AF293 and AF26, respectively. At the infinitely low Ce, the number of intracellular A. fumigatus conidia increased by 63.63 and 51.28% for AF293 and AF26, respectively. When the data were plotted against Ce, the Cs calculated using the sigmoid model was 0.52 mg/L for AF26 and 1.86 mg/L for AF293, respectively. A particularly interesting observation was how close the intracellular pharmacodynamics parameters of voriconazole against AF293 and AF26 became when the data were plotted against Ce/MIC, with Cs was 8.32-fold MIC for AF26 and 9.69-fold MIC for AF293, and E50 was 32.93-fold MIC for AF26 and 36.71-fold MIC for AF293 (Table 1 and Fig. 2b). This result suggests that Ce and the inherent susceptibility of A. fumigatus were the key factors influencing the voriconazole activity against A. fumigatus in the intracellular milieu. In other words, the Ce/MIC ratio might be a crucial determinant of the voriconazole intracellular activity against A. fumigatus.
Through the above analysis we were able to demonstrate that Ce/MIC might be the best predictor of the voriconazole intracellular antifungal effect. To our knowledge, the present study is the first to use infected A549 cells combined with an inhibitory effect sigmoid Emax model to study the PK/PD breakpoint. All of the data for the voriconazole intracellular activity against AF293 and AF26 are summarized in Fig. 3. The model showed that up to 84.01% of the initial intracellular inoculum was suppressed by voriconazole within 24 h (Fig. 3a). Given that 15.99% of the initial intracellular inoculum (about 4.4 lg cfu conidia) survived in the cells indicates that voriconazole could not completely suppress intracellular A. fumigatus even at higher incubation concentrations. This phenomenon may be due to the saturation of voriconazole absorption and the low voriconazole concentration in A549 cells [17, 26].
In the present study, we firstly found that the PK/PD breakpoint in the infected A549 model predicting the intracellular antifungal effect of voriconazole was Ce/MIC = 35.53, which was associated with a 50% suppression of intracellular A. fumigatus (Fig. 3b). Previous studies have shown that PK/PD parameters of antimicrobial agents could be useful for designing dosage regimens. Thus, Ce/MIC = 35.53 was utilized as the PK/PD index for the subsequent Monte Carlo simulation to design the prophylactic regimen for voriconazole. For cases of IPA that are mostly caused by airborne A. fumigatus, knowledge of the epidemiology of A. fumigatus is pivotal and could be used to underpin prophylactic strategies. The value of cumulative fraction of response is an estimate of the proportion of a population achieving the target PK/PD index. The results of Monte Carlo simulation showed that the effective suppression of intracellular A. fumigatus at least requires an oral dosage of 200 mg of voriconazole twice daily, which yielded a CFR value of 91.48% (Table 2). Actually, the Monte Carlo simulation result has been largely confirmed in clinical applications, because the dosage regimen has been widely demonstrated to be effective at preventing IPA [27–31]. For example, an oral dosage of 200 mg of voriconazole twice daily has been described as an appropriate and effective prophylactic agent in children and adults with acute myeloid leukemia or myelodysplastic syndrome and in those undergoing hematopoietic stem cell transplantation or solid-organ transplantation [32–35]. This study has also been demonstrated that the Ce/MIC breakpoint is a rational metric to use when designing dosage regimens for patients receiving voriconazole for prophylaxis.
The pulmonary ELF is on the surface of pulmonary epithelial cells and the concentration of an antifungal agent within the ELF is critical to the prophylaxis and treatment of the early stages of IPA [12, 36]. Clinically, patients typically take 200 mg of voriconazole orally twice daily—which is the recommended dosage regimen on the package insert—during prophylaxis applications. However, there was a lot of sub-therapeutic concentrations among some patients taking the oral dosage of 200 mg twice daily. For example, the concentration was adequate (< 1.0 mg/L) in 45% of allogeneic hematopoietic stem cell transplantation recipients [37]. Based on in vitro susceptibility data, the MIC90 (MIC at which 90% of the strain was inhibited) for voriconazole against A. fumigatus was 0.5 mg/L [38] and the Ce/MIC value was 35.53. We can thus calculate that the target voriconazole concentration in ELF for prophylaxis is 17.77 mg/L. However, the actual voriconazole concentration in ELF is difficult to measure, and it is much easier to measure it in plasma. According to the mean value of RELF/plasma, we calculated that the target voriconazole plasma concentration was 1.55 mg/L. A prospective, observational study showed that voriconazole trough concentration higher than 1.5 mg/L was the most effective for prophylaxis [39], which was close to the result of present study. Notably, when applying these results to clinical practice, we should characterize the local epidemiology of A. fumigatus and monitor the concentration of voriconazole in ELF or plasma, which would ensure the Ce/MIC > 35.53 for voriconazole against most isolates of A. fumigatus in patients receiving voriconazole prophylaxis. However, the relationship between Ce/MIC and clinical efficacy still needs to be verified in further clinical studies.
This study was subject to some major limitations. (1) The RELF/plasma value from the previous two studies may be somewhat unreliable due to the inclusion of only 20 samples; (2) currently the mechanism underlying how voriconazole is loaded into pulmonary epithelial cells remains unknown. The present study used extracellular voriconazole concentrations modelled on those reported in ELF fluid, whereas intracellular voriconazole may come from both ELF and plasma; (3) for the reliability of the results, additional experiments should be carried out in pulmonary epithelial cells different from A549. However, the method for building a cell model infected by A. fumigatus in this study is based on the study of Wasylnka et al. [16]., and the this model was proved to be reasonable and reliable in previous studies [17, 26, 40, 41]. Therefore, this cells model could afford a reliable result for this study.
Conclusions
Voriconazole exhibits concentration-dependent activity against intracellular A. fumigatus, and this study found that up to 84.01% of the intracellular inoculum was suppressed compared with the initial intracellular inoculum, and 50% suppression of the initial intracellular A. fumigatus was observed at a Ce/MIC breakpoint of 35.53. Additionally, this study identified the target voriconazole concentration in ELF and plasma was 17.77 and 1.55 mg/L, respectively, in patients receiving voriconazole prophylaxis. The present study has revealed the potential mechanism of voriconazole prophylaxis from the perspective of cellular PK/PD model, as well as provided a method for designing or optimizing prophylactic dosage regimens for voriconazole in high-risk immunosuppressed patients based on Monte Carlo simulation and the target voriconazole concentration. The reported findings have significant implications for reducing the morbidity of IPA in immunosuppressed patients.
Authors’ contributions
YD contributed to study design and was primarily responsible for writing the manuscript; TW, TZ, TM and JL conducted the experiments and analysis; YL, QY and LC contributed to data analysis; HD and LC revised the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank Professor Ling Lu (Jiangsu Key Laboratory for Microbes and Functional Genomics, Jiangsu Engineering and Technology Research Center for Microbiology, College of Life Sciences, Nanjing Normal University, Nanjing, China.) for supplying us with the A. fumigatus (AF293).
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets used during the current study are available from the corresponding author on reasonable request.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (Grant Numbers 81703618).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- IPA
Invasive pulmonary aspergillosis
- ELF
Epithelial lining fluid
- A549
Human pulmonary epithelial cells
- MIC
Minimum inhibitory concentration
- LDH
lactate dehydrogenase
- cfu
Colony forming units
- Ce/MIC
The ratio of extracellular voriconazole concentration divided minimum inhibitory concentration
- PK/PD
Pharmacokinetic/pharmacodynamics
- SDA
Sabouraud dextrose agar
- Cs
Static concentration
Contributor Information
Taotao Wang, Email: wangtaotao@stu.xjtu.edu.cn.
Tao Zhang, Email: hangtao_xjtu@126.com.
Ti Meng, Email: jasminemng@stu.xjtu.edu.cn.
Ying Li, Email: jiayuzhishi@stu.xjtu.edu.cn.
Lu Chen, Email: chenluc@stu.xjtu.edu.cn.
Qianting Yang, Email: yangqianting12@163.com.
Haiyan Dong, Email: dhy260113@sina.cn.
Jin’e Lei, Email: lq2640@163.com.
Limei Chen, Email: clmml@126.com.
Yalin Dong, Phone: +86-29-85323241, Email: dongyalin@mail.xjtu.edu.cn, Email: dongyalin@medmail.com.cn.
References
- 1.Schmiedel Y, Zimmerli S. Common invasive fungal diseases: an overview of invasive candidiasis, aspergillosis, cryptococcosis, and Pneumocystis pneumonia. Swiss Med Wkly. 2016;146:w14281. doi: 10.4414/smw.2016.14281. [DOI] [PubMed] [Google Scholar]
- 2.Bergeron A, Porcher R, Sulahian A, de Bazelaire C, Chagnon K, Raffoux E, Vekhoff A, Cornet M, Isnard F, Brethon B, et al. The strategy for the diagnosis of invasive pulmonary aspergillosis should depend on both the underlying condition and the leukocyte count of patients with hematologic malignancies. Blood. 2012;119(8):1831–1837. doi: 10.1182/blood-2011-04-351601. [DOI] [PubMed] [Google Scholar]
- 3.Groll AH, Silling G, Young C, Schwerdtfeger R, Ostermann H, Heinz WJ, Gerss J, Kolve H, Lanvers-Kaminsky C, Vieira Pinheiro JP, et al. Randomized comparison of safety and pharmacokinetics of caspofungin, liposomal amphotericin B, and the combination of both in allogeneic hematopoietic stem cell recipients. Antimicrob Agents Chemother. 2010;54(10):4143–4149. doi: 10.1128/AAC.00425-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pagano L, Caira M, Valentini CG, Posteraro B, Fianchi L. Current therapeutic approaches to fungal infections in immunocompromised hematological patients. Blood Rev. 2010;24(2):51–61. doi: 10.1016/j.blre.2009.11.003. [DOI] [PubMed] [Google Scholar]
- 5.Heng SC, Chen SC, Morrissey CO, Thursky K, Manser RL, De Silva HD, Halliday CL, Seymour JF, Nation RL, Kong DC, Slavin MA. Clinical utility of Aspergillus galactomannan and PCR in bronchoalveolar lavage fluid for the diagnosis of invasive pulmonary aspergillosis in patients with haematological malignancies. Diagn Microbiol Infect Dis. 2014;79(3):322–327. doi: 10.1016/j.diagmicrobio.2014.03.020. [DOI] [PubMed] [Google Scholar]
- 6.Patterson TF, Thompson GR, 3rd, Denning DW, Fishman JA, Hadley S, Herbrecht R, Kontoyiannis DP, Marr KA, Morrison VA, Nguyen MH, et al. Practice guidelines for the diagnosis and management of aspergillosis: 2016 update by the Infectious Diseases Society of America. Clin Infect Dis. 2016;63(4):e1–e60. doi: 10.1093/cid/ciw326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pappas PG, Kauffman CA, Andes DR, Clancy CJ, Marr KA, Ostrosky-Zeichner L, Reboli AC, Schuster MG, Vazquez JA, Walsh TJ, et al. Clinical practice guideline for the management of candidiasis: 2016 update by the Infectious Diseases Society of America. Clin Infect Dis. 2016;62(4):e1–e50. doi: 10.1093/cid/civ1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dagenais TR, Keller NP. Pathogenesis of Aspergillus fumigatus in invasive aspergillosis. Clin Microbiol Rev. 2009;22(3):447–465. doi: 10.1128/CMR.00055-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Theuretzbacher U, Ihle F, Derendorf H. Pharmacokinetic/pharmacodynamic profile of voriconazole. Clin Pharmacokinet. 2006;45(7):649–663. doi: 10.2165/00003088-200645070-00002. [DOI] [PubMed] [Google Scholar]
- 10.Tissot F, Agrawal S, Pagano L, Petrikkos G, Groll AH, Skiada A, Lass-Florl C, Calandra T, Viscoli C, Herbrecht R. ECIL-6 guidelines for the treatment of invasive candidiasis, aspergillosis and mucormycosis in leukemia and hematopoietic stem cell transplant patients. Haematologica. 2017;102(3):433–444. doi: 10.3324/haematol.2016.152900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cordonnier C, Rovira M, Maertens J, Olavarria E, Faucher C, Bilger K, Pigneux A, Cornely OA, Ullmann AJ, Bofarull RM, et al. Voriconazole for secondary prophylaxis of invasive fungal infections in allogeneic stem cell transplant recipients: results of the VOSIFI study. Haematologica. 2010;95(10):1762–1768. doi: 10.3324/haematol.2009.020073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hope WW, Drusano GL. Antifungal pharmacokinetics and pharmacodynamics: bridging from the bench to bedside. Clin Microbiol Infect. 2009;15(7):602–612. doi: 10.1111/j.1469-0691.2009.02913.x. [DOI] [PubMed] [Google Scholar]
- 13.Capitano B, Potoski BA, Husain S, Zhang S, Paterson DL, Studer SM, McCurry KR, Venkataramanan R. Intrapulmonary penetration of voriconazole in patients receiving an oral prophylactic regimen. Antimicrob Agents Chemother. 2006;50(5):1878–1880. doi: 10.1128/AAC.50.5.1878-1880.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Heng SC, Snell GI, Levvey B, Keating D, Westall GP, Williams TJ, Whitford H, Nation RL, Slavin MA, Morrissey O, Kong DC. Relationship between trough plasma and epithelial lining fluid concentrations of voriconazole in lung transplant recipients. Antimicrob Agents Chemother. 2013;57(9):4581–4583. doi: 10.1128/AAC.00942-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Park SJ, Mehrad B. Innate immunity to Aspergillus species. Clin Microbiol Rev. 2009;22(4):535–551. doi: 10.1128/CMR.00014-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wasylnka JA, Moore MM. Aspergillus fumigatus conidia survive and germinate in acidic organelles of A549 epithelial cells. J Cell Sci. 2003;116(Pt 8):1579–1587. doi: 10.1242/jcs.00329. [DOI] [PubMed] [Google Scholar]
- 17.Wang T, Yang Q, Chen L, Li Y, Meng T, Wang Y, Zhang T, Lei J, Xing J, Dong Y. Uptake and efflux kinetics, and intracellular activity of voriconazole against Aspergillus fumigatus in human pulmonary epithelial cells: a new application for the prophylaxis and early treatment of invasive pulmonary aspergillosis. Fundam Clin Pharmacol. 2017;31(3):311–318. doi: 10.1111/fcp.12262. [DOI] [PubMed] [Google Scholar]
- 18.Wang T, Zhu H, Sun J, Cheng X, Xie J, Dong H, Chen L, Wang X, Xing J, Dong Y. Efficacy and safety of voriconazole and CYP2C19 polymorphism for optimised dosage regimens in patients with invasive fungal infections. Int J Antimicrob Agents. 2014;44(5):436–442. doi: 10.1016/j.ijantimicag.2014.07.013. [DOI] [PubMed] [Google Scholar]
- 19.Wang T, Chen S, Sun J, Cai J, Cheng X, Dong H, Wang X, Xing J, Dong W, Yao H, Dong Y. Identification of factors influencing the pharmacokinetics of voriconazole and the optimization of dosage regimens based on Monte Carlo simulation in patients with invasive fungal infections. J Antimicrob Chemother. 2014;69(2):463–470. doi: 10.1093/jac/dkt369. [DOI] [PubMed] [Google Scholar]
- 20.Mouton JW, Dudley MN, Cars O, Derendorf H, Drusano GL. Standardization of pharmacokinetic/pharmacodynamic (PK/PD) terminology for anti-infective drugs: an update. J Antimicrob Chemother. 2005;55(5):601–607. doi: 10.1093/jac/dki079. [DOI] [PubMed] [Google Scholar]
- 21.Masterton RG, Kuti JL, Turner PJ, Nicolau DP. The OPTAMA programme: utilizing MYSTIC (2002) to predict critical pharmacodynamic target attainment against nosocomial pathogens in Europe. J Antimicrob Chemother. 2005;55(1):71–77. doi: 10.1093/jac/dkh511. [DOI] [PubMed] [Google Scholar]
- 22.Maertens J, Theunissen K, Verbeken E, Lagrou K, Verhaegen J, Boogaerts M, Eldere JV. Prospective clinical evaluation of lower cut-offs for galactomannan detection in adult neutropenic cancer patients and haematological stem cell transplant recipients. Br J Haematol. 2004;126(6):852–860. doi: 10.1111/j.1365-2141.2004.05140.x. [DOI] [PubMed] [Google Scholar]
- 23.Howard SJ, Lestner JM, Sharp A, Gregson L, Goodwin J, Slater J, Majithiya JB, Warn PA, Hope WW. Pharmacokinetics and pharmacodynamics of posaconazole for invasive pulmonary aspergillosis: clinical implications for antifungal therapy. J Infect Dis. 2011;203(9):1324–1332. doi: 10.1093/infdis/jir023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hope WW, Kruhlak MJ, Lyman CA, Petraitiene R, Petraitis V, Francesconi A, Kasai M, Mickiene D, Sein T, Peter J, et al. Pathogenesis of Aspergillus fumigatus and the kinetics of galactomannan in an in vitro model of early invasive pulmonary aspergillosis: implications for antifungal therapy. J Infect Dis. 2007;195(3):455–466. doi: 10.1086/510535. [DOI] [PubMed] [Google Scholar]
- 25.Jeans AR, Howard SJ, Al-Nakeeb Z, Goodwin J, Gregson L, Majithiya JB, Lass-Florl C, Cuenca-Estrella M, Arendrup MC, Warn PA, Hope WW. Pharmacodynamics of voriconazole in a dynamic in vitro model of invasive pulmonary aspergillosis: implications for in vitro susceptibility breakpoints. J Infect Dis. 2012;206(3):442–452. doi: 10.1093/infdis/jis372. [DOI] [PubMed] [Google Scholar]
- 26.Campoli P, Perlin DS, Kristof AS, White TC, Filler SG, Sheppard DC. Pharmacokinetics of posaconazole within epithelial cells and fungi: insights into potential mechanisms of action during treatment and prophylaxis. J Infect Dis. 2013;208(10):1717–1728. doi: 10.1093/infdis/jit358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Balogh J, Gordon Burroughs S, Boktour M, Patel S, Saharia A, Ochoa RA, McFadden R, Victor DW, Ankoma-Sey V, Galati J, et al. Efficacy and cost-effectiveness of voriconazole prophylaxis for prevention of invasive aspergillosis in high-risk liver transplant recipients. Liver Transpl. 2016;22(2):163–170. doi: 10.1002/lt.24365. [DOI] [PubMed] [Google Scholar]
- 28.Barreto JN, Beach CL, Wolf RC, Merten JA, Tosh PK, Wilson JW, Hogan WJ, Litzow MR. The incidence of invasive fungal infections in neutropenic patients with acute leukemia and myelodysplastic syndromes receiving primary antifungal prophylaxis with voriconazole. Am J Hematol. 2013;88(4):283–288. doi: 10.1002/ajh.23388. [DOI] [PubMed] [Google Scholar]
- 29.Siwek GT, Pfaller MA, Polgreen PM, Cobb S, Hoth P, Magalheas-Silverman M, Diekema DJ. Incidence of invasive aspergillosis among allogeneic hematopoietic stem cell transplant patients receiving voriconazole prophylaxis. Diagn Microbiol Infect Dis. 2006;55(3):209–212. doi: 10.1016/j.diagmicrobio.2006.01.018. [DOI] [PubMed] [Google Scholar]
- 30.Hayashi Y, Kanda Y, Nakamae H, Kanamori H, Ohashi K, Hidaka M, Yano S, Hatanaka K, Kohno A, Moriuchi Y. Voriconazole vs. itraconazole for antifungal prophylaxis in patients with GVHD: a randomized trial. Biol Blood Marrow Transpl. 2014;20(2):S91. doi: 10.1016/j.bbmt.2013.12.117. [DOI] [Google Scholar]
- 31.Hicheri Y, Cook G, Cordonnier C. Antifungal prophylaxis in haematology patients: the role of voriconazole. Clin Microbiol Infect. 2012;18(Suppl 2):1–15. doi: 10.1111/j.1469-0691.2012.03772.x. [DOI] [PubMed] [Google Scholar]
- 32.Maron GM, Hayden RT, Rodriguez A, Rubnitz JE, Flynn PM, Shenep JL, Knapp KM. Voriconazole prophylaxis in children with cancer: changing outcomes and epidemiology of fungal infections. Pediatr Infect Dis J. 2013;32(12):e451–e455. doi: 10.1097/INF.0b013e3182a74233. [DOI] [PubMed] [Google Scholar]
- 33.Sano H, Kobayashi R, Hori D, Kishimoto K, Suzuki D, Yasuda K, Kobayashi K. Prophylactic administration of voriconazole with two different doses for invasive fungal infection in children and adolescents with acute myeloid leukemia. J Microbiol Immunol Infect. 2016 doi: 10.1016/j.jmii.2016.05.002. [DOI] [PubMed] [Google Scholar]
- 34.Mattiuzzi GN, Cortes J, Alvarado G, Verstovsek S, Koller C, Pierce S, Blamble D, Faderl S, Xiao L, Hernandez M, Kantarjian H. Efficacy and safety of intravenous voriconazole and intravenous itraconazole for antifungal prophylaxis in patients with acute myelogenous leukemia or high-risk myelodysplastic syndrome. Support Care Cancer. 2011;19(1):19–26. doi: 10.1007/s00520-009-0783-3. [DOI] [PubMed] [Google Scholar]
- 35.Torres A, Serrano J, Rojas R, Martin V, Martin C, Tabares S, Molina JR, Capote M, Martinez F, Gomez P, Sanchez-Garcia J. Voriconazole as primary antifungal prophylaxis in patients with neutropenia after hematopoietic stem cell transplantation or chemotherapy for acute myeloid leukemia. Eur J Haematol. 2010;84(3):271–273. doi: 10.1111/j.1600-0609.2009.01367.x. [DOI] [PubMed] [Google Scholar]
- 36.Kiem S, Schentag JJ. Interpretation of antibiotic concentration ratios measured in epithelial lining fluid. Antimicrob Agents Chemother. 2008;52(1):24–36. doi: 10.1128/AAC.00133-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Trifilio S, Singhal S, Williams S, Frankfurt O, Gordon L, Evens A, Winter J, Tallman M, Pi J, Mehta J. Breakthrough fungal infections after allogeneic hematopoietic stem cell transplantation in patients on prophylactic voriconazole. Bone Marrow Transpl. 2007;40(5):451–456. doi: 10.1038/sj.bmt.1705754. [DOI] [PubMed] [Google Scholar]
- 38.Lamoth F, Alexander BD. Comparing Etest and broth microdilution for antifungal susceptibility testing of the most-relevant pathogenic molds. J Clin Microbiol. 2015;53(10):3176–3181. doi: 10.1128/JCM.00925-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mitsani D, Nguyen MH, Shields RK, Toyoda Y, Kwak EJ, Silveira FP, Pilewski JM, Crespo MM, Bermudez C, Bhama JK, Clancy CJ. Prospective, observational study of voriconazole therapeutic drug monitoring among lung transplant recipients receiving prophylaxis: factors impacting levels of and associations between serum troughs, efficacy, and toxicity. Antimicrob Agents Chemother. 2012;56(5):2371–2377. doi: 10.1128/AAC.05219-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Campoli P, Al Abdallah Q, Robitaille R, Solis NV, Fielhaber JA, Kristof AS, Laverdiere M, Filler SG, Sheppard DC. Concentration of antifungal agents within host cell membranes: a new paradigm governing the efficacy of prophylaxis. Antimicrob Agents Chemother. 2011;55(12):5732–5739. doi: 10.1128/AAC.00637-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Xu XY, Chen F, Sun H, Chen C, Zhao BL. Important factors mediates the adhesion of Aspergillus fumigatus to alveolar epithelial cells with E-cadherin. Am J Transl Res. 2016;8(5):2419–2425. [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used during the current study are available from the corresponding author on reasonable request.