

Abstract
Phellodendron amurense extract is a Chinese herbal remedy that has recently been studied for its antitumor, antimicrobial and other biological activities. It is previously unknown if these agents are bioavailable and effective against tumors when delivered as a dietary component. It is also unknown if the anti-tumorigenic properties of berberine, an isoquinoline alkaloid component of P. amurense, is equally effective when administered alone. There are contrasting reports on the cellular processes involved in anti-tumorigenesis by P. amurense and berberine. Here we find that berberine, when administered orally through the diet, inhibits in-vivo tumorigenesis of both p53 expressing and p53 null lung tumor xenografts equally whether administered in its pure form or as a part of P. amurense extract. We also show that berberine induces G1 cell cycle arrest, inhibits proliferative kinase signaling and arrests the growth of lung tumor cells in culture. Berberine administered in the diet was detectable by HPLC in the lungs of mice fed P. amurense or equivalent doses of berberine at concentrations of 455 and 518 ng/ml respectively and inhibited the growth of xenografted A549 cell tumors, which grew to 9.4 and 6.4 mm3 respectively, compared to 58.9 mm3 in control mice (p<0.001). Phosphorylation of Akt, CREB and MAPK was inhibited in A549 cells by P. amurense. Demonstration of oral bioavailability and anti-tumorigenic efficacy of dietary berberine, as well as further demonstration of signaling pathway modulation and cell-cycle arrest, implicate this relatively safe, natural compound as a potentially important therapeutic and chemopreventive agent for lung cancer.
Keywords: lung cancer, herbal therapy, cell-cycle, berberine
INTRODUCTION
Lung cancer is the leading cause of cancer death in both men and women in the United States [1]. Despite major advances in recent years, most lung cancers are disseminated at the time of presentation and have a mortality rate of about 90%, with a 5 year survival rate of only 16% [1]. This demonstrates an urgent need for better chemoprevention, treatment and adjuvant therapy for lung cancer.
Recently studies demonstrated that Phellodendron amurense (P. amurense) bark extract, prevented the development of prostate cancer in a murine model of prostate cancer chemoprevention [2] and inhibited prostate cancer cell proliferation [3] through inhibition of Akt/CREB mediated cyclin D1 activation. P. amurense extract inhibited CREB mediated Cox-2 activation [4], a signaling pathway involved in both prostate and lung cancer [5,6]. Berberine, an isoquinoline alkaloid present in many medicinal herbs including P. amurense extract, has pharmacological and biological activities including Cox-2 inhibition and other anti-inflammatory effects [7]. Recent reports have shown that berberine is one of the bioactive components of P. amurense extract [8]. Berberine has been studied for its inhibitory effect on proliferation and pro-apoptotic effect in several tumor cell lines, including non-small cell lung tumor cell lines [9–12], and has been shown to have tumor killing effects on lung cancer in-vivo when administered intraperitoneally [13,14]. Berberine has also been shown to inhibit prostate cancer and neuroblastoma cell growth through p53-dependent induction of apoptosis [15,16], and through p53-dependent cell cycle arrest and apoptosis in osteosarcoma cells induced by DNA damage [17].
Potential chemopreventive agents must be safe, effective, easily accessible and easily administered, preferably orally. We have previously demonstrated that the efficacy of chemopreventive/therapeutic agents can be dramatically affected by their inclusion in a complex mixture [18]. To our knowledge, no study has evaluated the efficacy of dietary berberine on tumorigenesis in an in-vivo model or compared the efficacy of berberine in-vivo to that of whole P. amurense extract. The goals of the current study were to determine the bioavailability and efficacy of berberine administered in the diet, to compare berberine alone to P. amurense extract and to determine the effect of inhibitory doses of dietary berberine on tumorigenesis related cellular phenotypes such as cell cycle arrest.
In demonstration of the efficacy of dietary berberine induced tumor growth arrest, we show that berberine, administered alone or in P. amurense extract are equally able to induce tumor cell growth arrest in association with G1 cell-cycle arrest regardless of p53 status. P. amurense extract and berberine induced G1 cell-cycle arrest in both p53 positive and null cells and dramatically inhibited the proliferation of these lung tumor cells in vitro. P. amurense extract concurrently reduced constitutive activation of Akt, CREB and MAPK indicating inhibition of an important proliferative signaling axis. Berberine was equally detectable in lung tissue following dietary administration of pure berberine or that as a part of P. amurense extract, indicating that relevant concentrations of berberine are achievable through oral administration whether given alone or in whole extract. No toxicity was observed even at the highest doses of berberine. Together, these findings show that dietary berberine and P. Amurense extract have significant potential as easily administered and safe lung cancer therapeutic agents and can inhibit proliferative kinase signaling pathways, arrest cell-cycle progression and prevent tumor growth.
MATERIALS AND METHODS
Proliferation Assays
The human lung tumor cell lines H1299 and A549 were cultured in RPMI-1640 plus 10% FBS (Life Technologies). Cells were counted using a hemacytometer and seeded onto 12-well tissue culture dishes at 500 cells per well. After attachment, cells were treated with DMSO solvent or the indicated concentrations of the P. amurense extract Nexrutine® (Solanova, Novato CA), or berberine (Sigma, St. Louis, MO) dissolved in DMSO. Final DMSO concentration was 0.025%. Mixtures were homogenized and were from a single batch. Cells were assayed for viable cell numbers in triplicate using the MTS based CellTiter 96 AQueous One Solution Cell Proliferation Assay kits (Promega). A490 was measured on a plate reader after attachment of cells (8 hours after seeding) for a baseline reading, and again throughout 5 days of growth in culture. Media and drugs were replaced every 2 days. p-values for biological triplicates were determined by a one-tailed Student’s T-test.
Tumorigenesis Models
A549 lung tumor cells were cultured, counted and resuspended in serum free media at a concentration of 5 × 106 cells/mL. A volume of 200 μL (1 × 106 cells) were injected subcutaneously into the right flank of athymic nude mice at an age of 4-6 weeks. Mice were monitored 3 times weekly and tumor sizes measured periodically until sacrifice at 4 weeks post-injection. Tumor volume was determined at 2 weeks post-injection by the formula for an ellipsoid π/6(l × w × h). P-values were determined by one-tailed Student’s t-test. Photographs were taken of tumors in live mice at 18 days post injection. At 4 weeks post-injection, mice were anesthetized by intra-peritoneal injection of a cocktail of 87mg/kg ketamine and 15mg/kg xylazine. Anesthesia was determined by absence of rear foot reflex. Blood was collected from the retro-orbital plexus into tubes containing 10μl of 0.5M EDTA. Mice were euthanized by CO2 asphyxiation. Lung tissues were collected, pooled within experimental groups of 4 mice and homogenized in DMSO using an electric tissue homogenizer. Lung and blood samples were centrifuged at 16,000 × g for 15 minutes at 4ºC. Serum and lung supernatants were collected and used for HPLC analysis as described below.
HPLC Analysis of Berberine Concentrations
The concentrations of Berberine in lung and serum samples were determined by HPLC. The HP 1100 series HPLC system consisted of an autosampler, a binary pump, a thermostat equipped column compartment, and a diode array detector (Agilent Tech, Santa Clara, CA). The HPLC column was 4.6 × 75 mm Zorbax SB-C18 3.5 μm column, and the detection wavelength was 343 nm. Stock solutions were prepared by dissolving Berberine in acetonitrile, and Nexrutine in DMSO, respectively. The mobile phase composed of A (20 mM phosphate buffer) - B(acetonitrile) was used under gradient elution of 0 min, 80:20, v/v; 6.5 min, 60:40 v/v; 10 min, 80:20 v/v. The column temperature was controlled as constant at 25 °C. The flow rate of the mobile phase was 1 ml/min during the whole gradient cycle.
Cell Cycle Analysis
After 48 hours in culture treated with the indicated concentrations of DMSO solvent, P. amurense extract, or berberine, 106 cells were washed and pelleted with PBS. Supernatant was removed and the pellet was dispersed by tapping. 500ul of cold 70% ethanol was added and incubated on ice for 20 minutes. Cell pellets were washed with PBS with 1% BSA, pelleted and resuspended in 100ul staining solution consisting of 10ug/ml propidium iodide and RNase A in PBS with 1% BSA. Cells in staining solution were diluted with 500ul PBS and analyzed on a Coulter flow cytometer for fluorescence associated with propidium iodide staining.
Western Blotting
Cells on 6-well tissue culture dishes were treated with the indicated concentrations of drugs for 48 hours at which time the cells were approximately 80% confluent. Cells were lysed with 250 μl of 1X sample buffer containing phosphatase and proteinase inhibitors, sonicated, and then boiled for 10 min. Gel loading was normalized by quantification of protein by Bradford assay. 20 μl was resolved on SDS-PAGE and subjected to immunoblot with the indicated antibodies. Band intensities on photographed film were measured using Image J software, a public domain application from NIH.
RESULTS
Phellodendron amurense extract and berberine inhibit lung tumor cell proliferation
To determine the effect of P. amurense extract and berberine on lung tumor cell proliferation, we treated p53 negative H1299 and p53 positive A549 lung tumor cell lines with 0, 2.5 or 5 μg/ml of P. amurense extract or 0, 25, or 50 μM berberine. Cells were counted, seeded at low density and monitored for proliferation using an MTS assay. Both P. amurense extract and berberine treatment caused dose dependent inhibition of proliferation of lung tumor cell lines (Fig. 1). Treatment with P. amurense extract at 2.5 and 5 μg/ml inhibited proliferation by 58 and 68% respectively in A549 cells and by 18 and 44% respectively in H1299 cells. Treatment with berberine at 25 and 50 μM concentrations inhibited proliferation by 90 and 100% respectively in A549 cells and by 76 and 100% respectively in H1299 cells. Apoptosis was undetectable at 24 or 48 hours at highest doses as measured by Caspase3/7 activity (data not shown).
Figure 1. P. amurense extract or Berberine treatment causes dose dependent inhibition of growth of lung tumor cell lines.
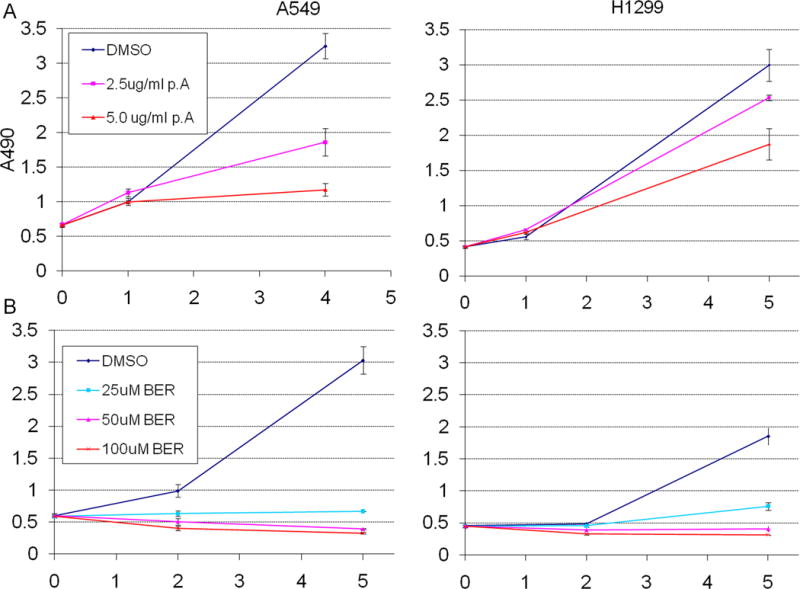
Cell proliferation over 5 days in culture as measured by MTS assay after (A) treatment with P. amurense extract at 2.5 and 5 μg/ml in A549 cells and H1299 cells. (B) treatment with berberine at 25 and 50 μM in A549 cells H1299 cells. Error bars represent standard deviations of biological triplicates. All treatment groups were significantly less proliferative than controls (p≤0.005 at day 5). The horizontal axes represent days post-seeding.
P. amurense extract and berberine induce G1 cell cycle arrest in lung tumor cells
To determine if the inhibition of proliferation observed was associated with cell cycle arrest, both H1299 and A549 cells were treated with either DMSO, 5 μg/ml P. amurense extract or 50 μM berberine for 48 hours. DNA content was determined by propidium iodide staining and flow cytometry to analyze the cell cycle distribution of treated and control cell populations. Signal within the 2N DNA content peak was quantified to identify those cells in the G0/G1 phase of the cell cycle. Approximately 45% of H1299 cells treated with DMSO alone had 2N DNA content, compared to 67% of those treated with P. amurense extract and 81% of those treated with berberine (Fig. 2A). Likewise, A549 cells with 2N DNA content were increased from 63% in control cells to 74% and 77% with P. amurense extract and berberine treatment, respectively. Cyclin B1, a cell cycle regulating cyclin that is present only during late S phase and G2 phase of the cell cycle was quantified by western blot analysis (Fig. 2B). Both P. amurense extract and berberine treatment resulted in decreased cyclin B1 content in A549 cells. These results are consistent with the concept that treatment of lung tumor cells with P. amurense extract or berberine results in cell cycle arrest at G1 phase. These cell lines exhibited the expected p53 status, with A549 expressing p53 and H1299 not expressing p53 (Fig. 2C).
Figure 2. P. amurense extract and berberine induce G1 cell cycle arrest and decrease Akt, CREB and MAPK phosphorylation in lung tumor cells.
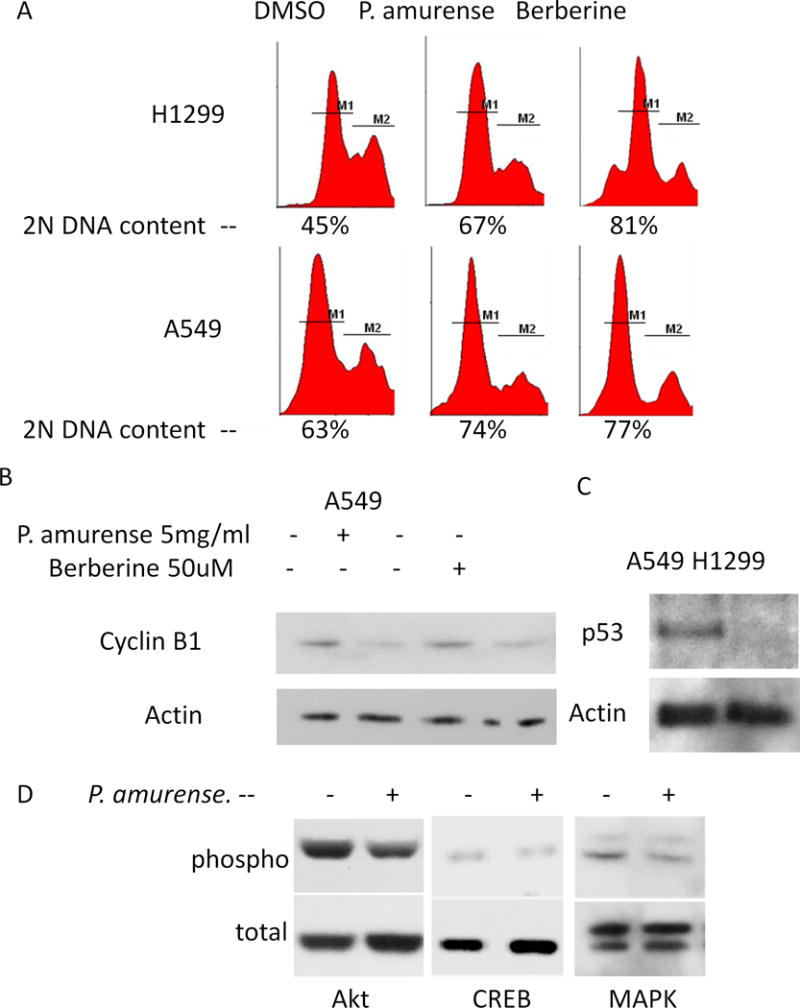
(A) DNA content analysis by PI staining and flow cytometry demonstrates accumulation of cells in G1 phase upon treatment of either P. amurense extract or berberine. (B) A representative immunoblot for cyclin B1 in cells treated for 48 hours. P. amurense extract and berberine treatment both resulted in decreased accumulation of Cyclin B1 indicating G1 arrest. (C) p53 status of A549 and H1299 cells represented by immunoblot. (D) Representative immunoblots for total and phosphorylated Akt, CREB and MAPK demonstrates decreased phosphorylation in A549 cells treated with 5μg/ml P. amurense extract for 48 hours.
P. amurense extract inhibits sustained activation of Akt, CREB and MAPK
In order to determine if the Akt/CREB signaling axis is affected by P. amurense extract in lung tumor cells, as has been observed in prostate tumor cells [3], A549 cells were treated with P. amurense extract or DMSO solvent for 48 hours and subjected to western blotting for total and phosphorylated Akt, CREB and MAPK. With P. amurense extract treatment, phospho-Akt, phosphor-CREB and phosphor-MAPK levels were decreased by 47%, 44% and 33%, respectively, when compared to DMSO control (Fig. 2D). No difference in acute activation of Akt, CREB, or MAPK was observed after starvation for 50 hours followed by serum stimulation and drug treatment for 1 hour (data not shown).
Dietary Phellodendron amurense extract and berberine are delivered equally to lung tissue and equally inhibit tumorigenesis in-vivo
To determine the berberine content of commercially available P. amurense extract, and to learn whether an amount of P. amurense extract that contains an equivalent amount of berberine has additional therapeutic effects, we determined the berberine content of P. amurense extract using HPLC analysis. Berberine was detected at 343nm. Standard curves were established using known concentrations of berberine and P. amurense extract, yielding linear regressions with slopes (concentration/signal) of 0.0757 for berberine and 0.2289 for P. amurense extract (Fig. 3). This indicates that the extract contains 33% berberine.
Figure 3. HPLC Detection of Berberine.
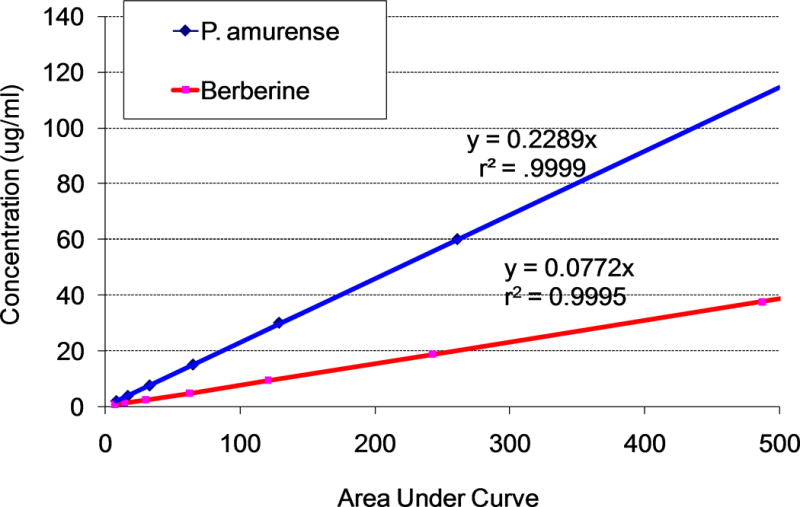
Standard curves for detection of the peak representing berberine in P. amurense extract and pure berberine, yielding linear regressions with slopes (concentration/signal) of 0.0757 for berberine and 0.2289 for P. amurense extract. The concentration of berberine in P. amurense extract is 33% of that in purified berberine. Berberine was detected at 343nm.
Based on our HPLC analysis and effective dosage in previous studies [11], we administered berberine to athymic nude mice in powdered diet at 1000 or 1800 parts per million (ppm), and P. amurense extract at 3000 or 5400 ppm to give equivalent amounts of berberine. One week after initiation of these diets or a control diet consisting of base powdered diet alone, all mice were injected sub-cutaneously in the right flank with 1 × 106 A549 lung tumor cells. Mice were monitored regularly for health, weight and tumor volume. All mice appeared healthy and gained weight appropriately throughout the study (data not shown). Both P. amurense extract and berberine inhibited tumor growth in a dose dependent manner. At 14 days post injection, the high doses of both P. amurense extract and berberine resulted in dramatically and significantly (p<0.001) decreased tumor volume at 9.4 and 6.4 mm3 respectively compared to 58.9 mm3 in control mice (Fig. 4). 3000 ppm P. amurense extract and 1000 ppm berberine also significantly decreased tumor growth at 28.8 (p<0.02) and 20.4 (p<0.05) mm3 respectively compared to control mice. Hence, treatment with P. amurense extract did not appear to offer a distinct advantage to administering pure berberine.
Figure 4. Dietary P. amurense Extract and Berberine Equally Inhibit Tumorigenesis of Lung Tumor Cells in-vivo.
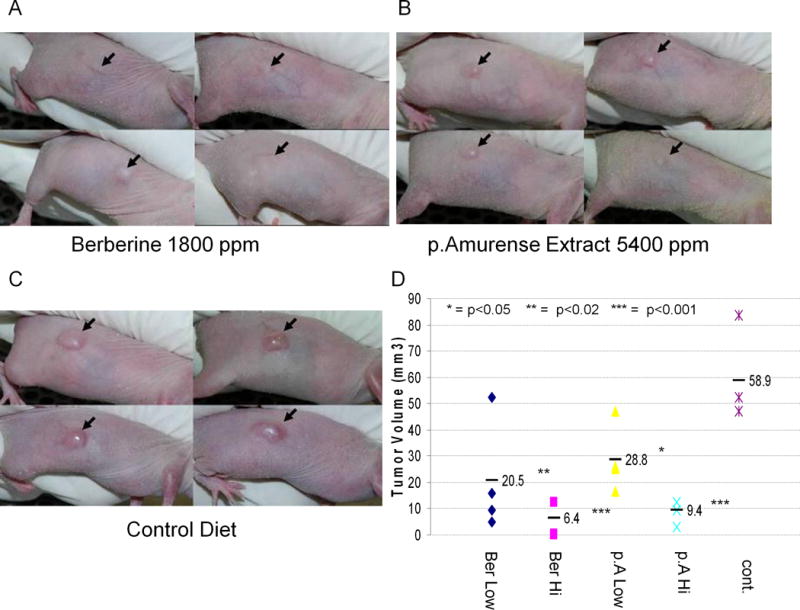
(A) Xenografted A549 tumors in athymic nude mice fed 1800 ppm berberine in powdered diet ad-libitum. (B) Tumors in mice fed 5400 ppm P. amurense Extract. (C) Tumors in mice fed control diet. (D) Tumor volumes at 14 days after injection of tumor cells, showing significant dose dependent inhibition of tumor growth by both treatments. n=4 mice per group.
To determine the physiological concentration of berberine in the nude mice used in these experiments, we performed HPLC analysis on pooled lung homogenates. Lungs were pooled in order to achieve the concentrations necessary to detect berberine by HPLC. Lung homogenates from mice fed control diet, 3000 ppm P. amurense extract and 1000 ppm berberine contained 0, 455 and 518 ng/ml berberine respectively (data not shown). Thus mice fed P. amurense extract in doses intended to replicate the amount of berberine administered in the pure berberine group, exhibited similar physiologic concentrations of berberine in the lung. This demonstrates that berberine administered as a pure compound or in the extract is equally bioavailable and detectable by HPLC.
DISCUSSION
We demonstrate the novel finding that berberine is equally effective in inhibiting the growth of lung tumors whether administered as pure berberine or P. amurense extract, and that berberine is effective when administered orally through the diet. Furthermore, we show that this effect is in association with G1 cell cycle arrest and is observed regardless of p53 status. Sub-G1 DNA content, suggesting apoptosis, was only observed at the highest concentrations of berberine. The concentration of berberine used in previous studies demonstrating apoptosis in lung tumor cells [11] was equal to the highest dose we used, up to twice that dose. Berberine has been shown to induce mitochondrial dysfunction and apoptosis at higher doses in tumor cells [19] and even in promonocytic cells [20], although this raises concerns about toxicity at such doses. Berberine and the class of compounds known as protoberberines have topoisomerase poisoning properties [21]. Although berberine has low activity on topoisomerase compared to other protoberberines [21], this activity may play a role in it’s p53 dependent induction of apoptosis at higher doses. Our results suggest that berberine may have safer effects through cell cycle arrest in a p53 independent manner at a sub-apoptotic dose. Consistent with published data, we observed a greater effect on proliferation in A549 cells than in H1299 cells. The difference in effect between these two cell lines may be due to the difference in p53 status; however, variation between cell lines is expected, and it cannot be concluded that these differences reflect p53 dependent effects. The growth and cell cycle arrest observed cannot be compared for quantitative differences between P. amurense extract and berberine because, as we later determined by HPLC analysis, the extract used in the assays contains 33% berberine. This resulted in a much lower berberine concentration in the P. amurense extract treated cells than in the berberine treated cells.
Our in-vivo tumorigenesis models suggest that and equivalent concentration of pure berberine is equally effective as P. amurense extract in preventing tumor growth; supporting the notion that berberine is indeed the major bioactive and anti-tumorigenic constituent of P. amurense extract. Although we were unable to accurately measure dietary consumption, we casually observed slightly less consumption of the high dose P. amurense extract, which may have resulted in less efficacy in preventing tumor growth. However, we did show similar levels of berberine in the lungs of mice fed berberine and P. amurense extract. The levels of berberine found in the experimental lungs were less than 1 ug/ml, which is well under the NCI maximum concentration guidelines for potential anti-cancer drugs. No toxicity was observed with P. amurense treatment in any of the mice as monitored by body weight. The highest dose used in this study is 50 fold less than the maximum safe oral dose in mice [22]. We demonstrate that P. amurense extract inhibits Akt, CREB and MAPK phosphorylation in lung tumor cells, further implicating this signaling axis in cell cycle arrest by berberine. Further investigation of these signaling molecules as underlying effectors of cell cycle arrest by berberine is warranted.
Demonstration that berberine is effective against tumorigenesis when administered orally through the diet, either alone or as a part of a whole plant extract, implicate berberine as a bio-available and effective anti-tumor compound. These results warrant further clinical investigation of berberine and berberine containing herbal products as natural and safe chemopreventive and therapeutic agents against lung cancer. Further mechanistic studies of berberine mediated anti-tumorigenic effects may delineate important targets for lung cancer prevention or treatment.
Acknowledgments
We would like to thank Albert Shih and Dr. Qi Zhang for their assistance with mouse tumorigenesis experiments; Julie Dimitri for assistance with flow cytometry; and Haris Vikis, Jay Tichelar and Pengyuan Liu for their critical input on the manuscript.
This research was supported by NIH grant R01 CA139959.
References
- 1.Jemal A, Siegel R, Ward E, et al. Cancer statistics, 2008. CA Cancer J Clin. 2008;58(2):71–96. doi: 10.3322/CA.2007.0010. [DOI] [PubMed] [Google Scholar]
- 2.Kumar AP, Bhaskaran S, Ganapathy M, et al. Akt/cAMP-responsive element binding protein/cyclin D1 network: a novel target for prostate cancer inhibition in transgenic adenocarcinoma of mouse prostate model mediated by Nexrutine, a Phellodendron amurense bark extract. Clin Cancer Res. 2007;13(9):2784–2794. doi: 10.1158/1078-0432.CCR-06-2974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Garcia GE, Nicole A, Bhaskaran S, Gupta A, Kyprianou N, Kumar AP. Akt-and CREB-mediated prostate cancer cell proliferation inhibition by Nexrutine, a Phellodendron amurense extract. Neoplasia. 2006;8(6):523–533. doi: 10.1593/neo.05745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ghosh R, Garcia GE, Crosby K, et al. Regulation of Cox-2 by cyclic AMP response element binding protein in prostate cancer: potential role for nexrutine. Neoplasia. 2007;9(11):893–899. doi: 10.1593/neo.07502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Han S, Roman J. COX-2 inhibitors suppress lung cancer cell growth by inducing p21 via COX-2 independent signals. Lung Cancer. 2006;51(3):283–296. doi: 10.1016/j.lungcan.2005.10.015. [DOI] [PubMed] [Google Scholar]
- 6.Qadri SS, Wang JH, Redmond KC, AF OD, Aherne T, Redmond HP. The role of COX-2 inhibitors in lung cancer. Ann Thorac Surg. 2002;74(5):1648–1652. doi: 10.1016/s0003-4975(02)04022-5. [DOI] [PubMed] [Google Scholar]
- 7.Kuo CL, Chi CW, Liu TY. The anti-inflammatory potential of berberine in vitro and in vivo. Cancer Lett. 2004;203(2):127–137. doi: 10.1016/j.canlet.2003.09.002. [DOI] [PubMed] [Google Scholar]
- 8.Muralimanoharan SB, Kunnumakkara AB, Shylesh B, et al. Butanol fraction containing berberine or related compound from nexrutine inhibits NFkappaB signaling and induces apoptosis in prostate cancer cells. Prostate. 2009;69(5):494–504. doi: 10.1002/pros.20899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Iizuka N, Miyamoto K, Okita K, et al. Inhibitory effect of Coptidis Rhizoma and berberine on the proliferation of human esophageal cancer cell lines. Cancer Lett. 2000;148(1):19–25. doi: 10.1016/s0304-3835(99)00264-5. [DOI] [PubMed] [Google Scholar]
- 10.Iwasa K, Moriyasu M, Yamori T, Turuo T, Lee DU, Wiegrebe W. In vitro cytotoxicity of the protoberberine-type alkaloids. J Nat Prod. 2001;64(7):896–898. doi: 10.1021/np000554f. [DOI] [PubMed] [Google Scholar]
- 11.Katiyar SK, Meeran SM, Katiyar N, Akhtar S. p53 cooperates berberine-induced growth inhibition and apoptosis of non-small cell human lung cancer cells in vitro and tumor xenograft growth in vivo. Mol Carcinog. 2008 doi: 10.1002/mc.20453. [DOI] [PubMed] [Google Scholar]
- 12.Kuo CL, Chi CW, Liu TY. Modulation of apoptosis by berberine through inhibition of cyclooxygenase-2 and Mcl-1 expression in oral cancer cells. In Vivo. 2005;19(1):247–252. [PubMed] [Google Scholar]
- 13.Mitani N, Murakami K, Yamaura T, Ikeda T, Saiki I. Inhibitory effect of berberine on the mediastinal lymph node metastasis produced by orthotopic implantation of Lewis lung carcinoma. Cancer Lett. 2001;165(1):35–42. doi: 10.1016/s0304-3835(00)00710-2. [DOI] [PubMed] [Google Scholar]
- 14.Peng PL, Kuo WH, Tseng HC, Chou FP. Synergistic tumor-killing effect of radiation and berberine combined treatment in lung cancer: the contribution of autophagic cell death. Int J Radiat Oncol Biol Phys. 2008;70(2):529–542. doi: 10.1016/j.ijrobp.2007.08.034. [DOI] [PubMed] [Google Scholar]
- 15.Choi MS, Oh JH, Kim SM, et al. Berberine inhibits p53-dependent cell growth through induction of apoptosis of prostate cancer cells. International journal of oncology. 2009;34(5):1221–1230. [PubMed] [Google Scholar]
- 16.Choi MS, Yuk DY, Oh JH, et al. Berberine inhibits human neuroblastoma cell growth through induction of p53-dependent apoptosis. Anticancer research. 2008;28(6A):3777–3784. [PubMed] [Google Scholar]
- 17.Liu Z, Liu Q, Xu B, et al. Berberine induces p53-dependent cell cycle arrest and apoptosis of human osteosarcoma cells by inflicting DNA damage. Mutation research. 2009;662(1–2):75–83. doi: 10.1016/j.mrfmmm.2008.12.009. [DOI] [PubMed] [Google Scholar]
- 18.Fu H, He J, Mei F, et al. Cancer prevention research. 6. Vol. 2. Philadelphia, Pa: 2009. Lung cancer inhibitory effect of epigallocatechin-3-gallate is dependent on its presence in a complex mixture (polyphenon E) pp. 531–537. [DOI] [PubMed] [Google Scholar]
- 19.Youn MJ, So HS, Cho HJ, et al. Berberine, a natural product, combined with cisplatin enhanced apoptosis through a mitochondria/caspase-mediated pathway in HeLa cells. Biol Pharm Bull. 2008;31(5):789–795. doi: 10.1248/bpb.31.789. [DOI] [PubMed] [Google Scholar]
- 20.Jantova S, Cipak L, Letasiova S. Berberine induces apoptosis through a mitochondrial/caspase pathway in human promonocytic U937 cells. Toxicol In Vitro. 2007;21(1):25–31. doi: 10.1016/j.tiv.2006.07.015. [DOI] [PubMed] [Google Scholar]
- 21.Qin Y, Pang JY, Chen WH, Zhao ZZ, Liu L, Jiang ZH. Inhibition of DNA topoisomerase I by natural and synthetic mono- and dimeric protoberberine alkaloids. Chemistry & biodiversity. 2007;4(3):481–487. doi: 10.1002/cbdv.200790040. [DOI] [PubMed] [Google Scholar]
- 22.Kheir MM, Wang Y, Hua L, et al. Acute toxicity of berberine and its correlation with the blood concentration in mice. Food Chem Toxicol. 48(4):1105–1110. doi: 10.1016/j.fct.2010.01.033. [DOI] [PubMed] [Google Scholar]