
Figure 1.
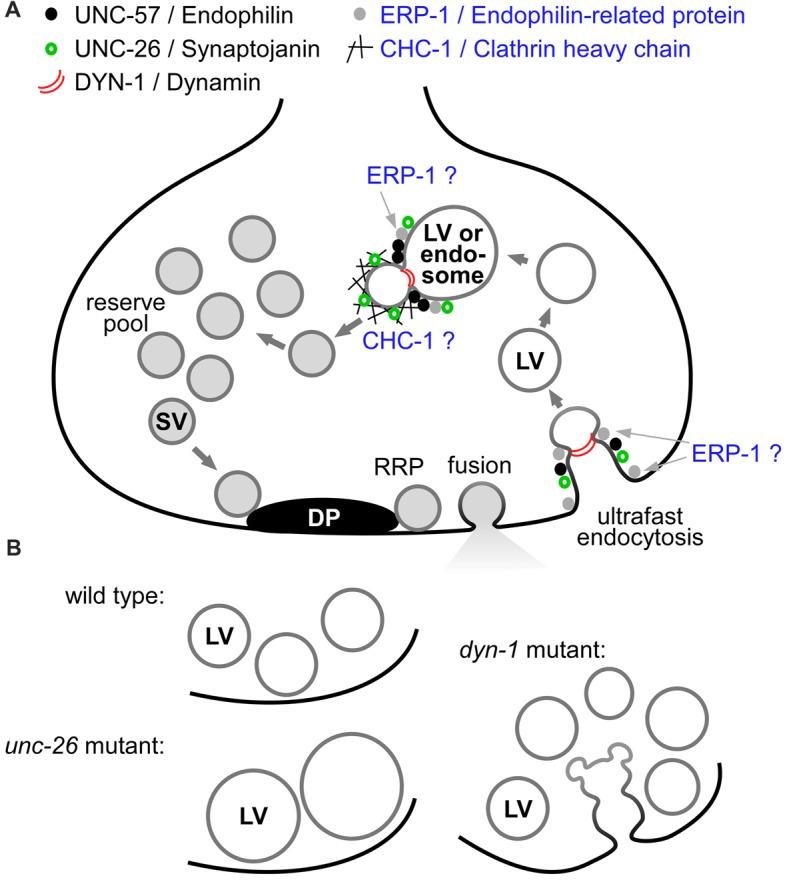
Model of the synaptic vesicle (SV) cycle and select proteins involved in distinct steps of SV recycling. (A) SVs are formed by budding from synaptic endosomes, entering a reserve pool (RP) after filling with neurotransmitter (gray lumen). At the dense projection (DP), SVs approach the plasma membrane (PM; black line) and become docked and primed, forming the readily releasable pool (RRP). After fusion, ultrafast endocytosis retrieves the SV membrane and proteins, forming clear-core large vesicles (LVs), following membrane severing by dynamin (red arcs). The LVs fuse with or transform into synaptic endosomes. Several proteins studied either previously (black dots: UNC-57/endophilin A, green open circles: UNC-26/synaptojanin, red arcs: DYN-1/dynamin; Kittelmann et al., 2013), or as part of this work (black mesh: CHC-1/clathrin heavy chain, gray dots: ERP-1/endophilin B), affect different steps of the recycling pathway, acting either at the PM or at the synaptic endosome. Roles/sites of action of clathrin and endophilin-related protein (ERP-1; blue text) are not firmly established as yet in C. elegans. (B) Summary of mutant phenotypes as observed at the ultrastructural level, following photostimulation of cholinergic neurons (Kittelmann et al., 2013): in wild type, LVs form transiently. In the unc-26 mutant, less LVs form, but they do form and are persistent, as they cannot be efficiently broken down. In the dyn-1 mutant, LVs are formed but cannot be pinched off the PM, and smaller vesicles emerge from these structures (possibly new SVs) that can also not be pinched off. LVs that were formed nonetheless are accumulating and cannot be processed to SVs efficiently.