
Figure 2. Development and cyclic remodelling of dWAT.
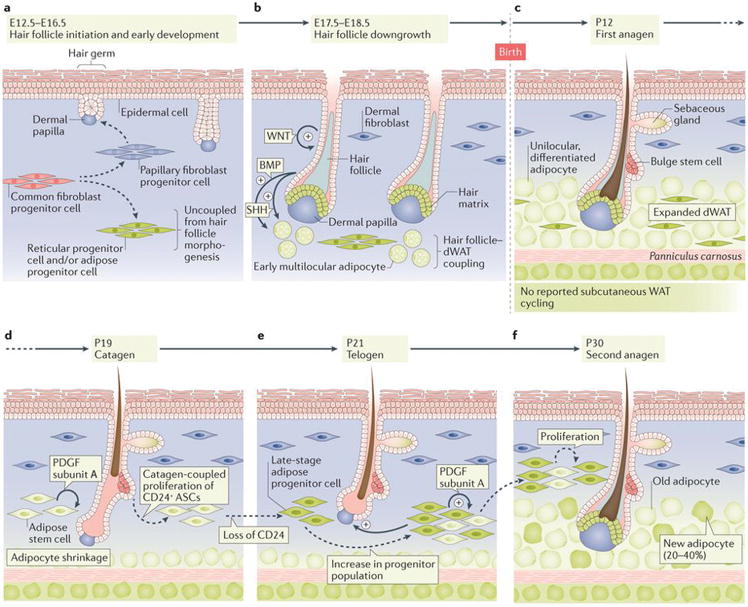
(a) In mice, the initial development of hair follicles and dWAT, which occurs between embryonic days (E) 12.5 to E16.5, are largely independent from one another41,42. Reticular progenitor cells and adipose progenitor cells arise from common fibroblast progenitors. Hair follicles develop at the interaction sites between embryonic epidermis and papillary fibroblast progenitor cells. (b) During E17.5 to E18.5, development of hair follicles and dWAT are coupled4,41,44,45,110. Enlarged hair follicles activate several pro-adipogenic signalling pathways, including Hedgehog45 and Bmp48. In response to hair follicle-derived signals, adipose progenitors rapidly differentiate. Small multilocular adipocytes appear first and then mature into large unilocular cells over several days. (c) dWAT peaks in thickness by postnatal day (P) 12, which is when anagen hair follicles fully develop4. dWAT contains differentiated adipocytes and adipocyte progenitor cells. (d, e) Between P19 to 21, hair follicles undergo regression into telogen stage. Concomitantly, dWAT collapses, partially through cell lipolysis to less than half of its anagen size4,41. At the same time, adipose stem cells (ASCs) expand via proliferation and give rise to late stage adipose progenitor cells (green)6. Autocrine Pdgfα mediates the expansion of adipose progenitor cells46. (f) During second anagen, up to 40% of adipocytes in enlarged dWAT are new, derived from expanded progenitors45.