
Figure 1.
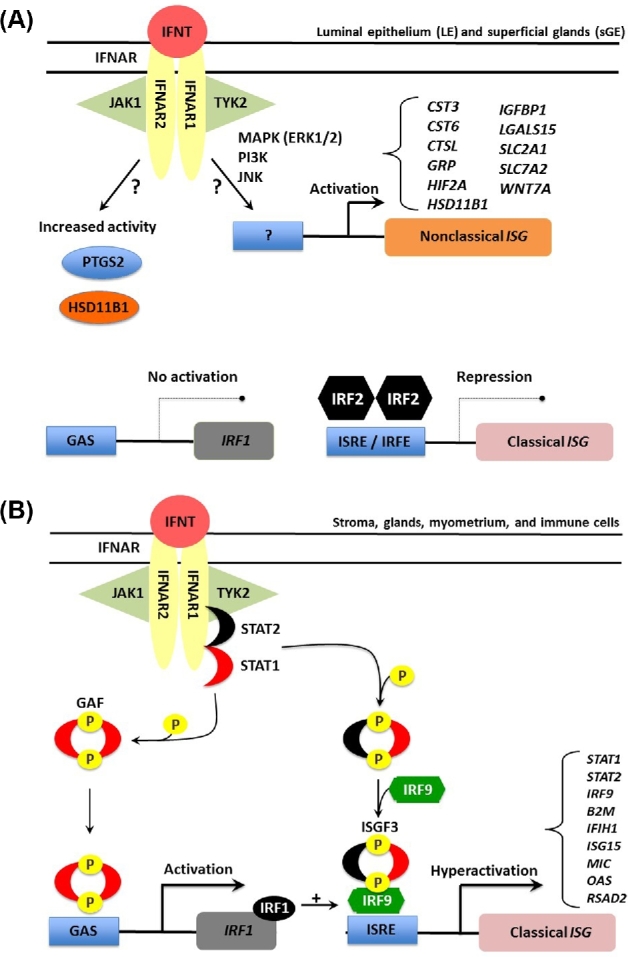
Schematic illustrating the current working hypothesis on IFNT signaling in endometrial epithelia and stroma of the ovine uterus. IFNT, produced in large amounts by the developing conceptus, binds to the Type I IFN receptor (IFNAR) present on cells of the ovine endometrium. In the cells of the endometrial luminal epithelium (LE) and superficial glands (sGE), IFNT induces IRF2 which prevents uterine LE and sGE from expressing classical ISGs such as STAT1, STAT2, and IRF9 (A). IRF2, a potent and stable repressor of transcription of genes in the nucleus, increases in abundance during early pregnancy only in uterine LE and sGE. The presence of IRF2 (interferon regulatory factor 2) inhibits binding of IRF1 to the promoter region of ISRE-containing target genes through direct competitive binding of IRF2 to ISRE and subsequent coactivator repulsion. IFNT can stimulate the transcription of a number of nonclassical IFNT-stimulated genes (ISG) as well as increase the activity of certain intracellular enzymes (prostaglandin synthase 2, PTGS2 and HSD11B1, hydroxysteroid 11-beta dehydrogenase). However, the noncanonical signaling pathways mediating these effects of IFNT in the LE/sGE are largely unknown. In cells of the stroma and middle to deep glands (B), IFNT-mediated association of the IFNAR subunits facilitates the cross-phosphorylation and activation of two Janus kinases, Tyk2 and JAK1, which in turn phosphorylate the receptor and creates a docking site for signal transducer and activator of transcription 2 (STAT2). STAT2 is then phosphorylated, thus creating a docking site for STAT1 that is then phosphorylated. STAT1 and STAT2 are then released from the receptor and can form two transcription factor complexes. Interferon-stimulated gene factor 3 (ISGF3), formed by association of the STAT1-2 heterodimer with IRF9 in the cytoplasm, translocates to the nucleus, and transactivates genes containing an ISRE, such as STAT1, STAT2, and IRF9. Gamma activation factor (GAF) is formed by binding of STAT1 homodimers, which translocates to the nucleus and transactivate genes containing a gamma activation sequence (GAS) element(s), such as IRF1. IRF1 can also bind and transactivate ISRE-containing genes. The simultaneous induction of STAT2 and IRF9 gene expression by IFNT appears to shift transcription factor formation from GAF towards predominantly ISGF3. Therefore, IFNT activation of the JAK-STAT signal transduction pathway allows for constant formation of ISGF3 and GAF transcription factor complexes and hyperactivation of ISG expression.