

Abstract
The trithorax genes encode an evolutionarily conserved family of proteins that function to maintain specific patterns of gene expression throughout cellular development. Members of this protein family contain a highly conserved 130- to 140-amino acid motif termed the SET domain. We report the purification and molecular identification of the subunits of a protein complex in the yeast Saccharomyces cerevisiae that includes the trithorax-related protein Set1. This protein complex, which we have named COMPASS (Complex Proteins Associated with Set1), consists of seven polypeptides ranging from 130 to 25 kDa. The same seven proteins were identified in COMPASS purified either by conventional biochemical chromatography or tandem-affinity tagging of the individual subunits of the complex. Null mutants missing any one of the six nonessential subunits of COMPASS grow more slowly than wild-type cells under normal conditions and demonstrate growth sensitivity to hydroxyurea. Furthermore, gene expression profiles of strains missing either of two nonessential subunits of COMPASS are altered in similar ways, suggesting these proteins have similar roles in gene expression in vivo. Molecular characterization of trithorax complexes will facilitate defining the role of this class of proteins in the regulation of gene expression and how their misregulation results in the development of human cancer.
Keywords: transcription‖gene expression‖MLL‖leukemia
During development of metazoans, different cells become committed to different fates, in part through heritable, quasi-stable changes in gene expression. Two families of proteins, the trithorax (trx) group and the polycomb (Pc) group, have been demonstrated to play important roles in this process (1, 2). These two chromatin-associated classes of proteins are known to function by activating or repressing transcription. Both trx and Pc group proteins contain a 130- to 140-amino acid motif called the SET domain, which is found in a variety of chromatin-associated proteins (3, 4). This domain takes its name from the Drosophila proteins Su(var)3–9, Enhancer of zeste [E(z)], and trithorax (trx) (5, 6). Known targets of the trx and Pc group proteins are the clustered homeotic genes in the bithorax and Antennapedia gene complexes in Drosophila (7). Mutations in the trx group of genes suggest that they function as positive effectors of gene expression, whereas mutations in the Pc group of genes suggest that they repress transcription (8–10).
Mammalian cells seem to possess Trx and Pc group proteins whose roles seem to be similar to those of their Drosophila counterparts (11–13). Mutations and/or translocations into the trx (MLL) gene in mammals result in the development of several types of hematological malignancies (14). Despite the fundamental role of the trx group of proteins in development and their role in oncogenesis, very little is know about their molecular properties and biochemical functions. Here, we report the purification and molecular characterization of a trithorax-containing complex from the yeast Saccharomyces cerevisiae, which we have named COMPASS (Complex Proteins Associated with Set1).
Materials and Methods
Materials.
Media and reagents were purchased from Sigma. Western development reagents were purchased from ICN. Glycerol (spectra-analyzed grade), potassium chloride, Hepes, Tris, and ammonium sulfate were purchased from Fisher. Anti-SET1 antibodies (BL2 and BL2,3) were obtained from Lorraine Pillus (University of California at San Diego, La Jolla, CA).
Conventional Purification and Tandem Affinity Purification (TAP) of COMPASS and Protein Identification.
Purification of the Set1 complex was carried out at 4°C. All purification steps were performed in duplicate following the purification scheme shown in Fig. 1C. Fractions were tested for Set1 by Western analysis with BL2,3 or BL2 anti-Set1 polyclonal antibodies. For the affinity purification of COMPASS, subunits of COMPASS were tagged by chromosomal integration via the C-terminal domain of each COMPASS subunit following a published method (15). Tagged COMPASS complexes were purified essentially as described on IgG and calmodulin columns from extracts obtained from 5-liter yeast cultures grown in yeast extract peptone dextrose medium to an OD600 of 1.0–1.5 as before (15, 16). The protein bands were reduced, alkylated, and subjected to in-gel tryptic digestion, and the peptides were then purified and identified by matrix-assisted laser desorption ionization–time-of-flight (MALDI-TOF) spectrometry by using a PerSeptive DE STR (17). Selected mass values from the MALDI-TOF experiments were taken to search the protein nonredundant database (NR; National Center for Biotechnology Information, Bethesda, MD) using the PeptideSearch algorithm. MS/MS spectra were inspected for y" ion series to compare with the computer-generated fragment ion series of the predicted tryptic peptides (18, 19).
Figure 1.

Conventional purification of the SET1-containing complex. (A) Western analysis of yeast strain 17A1 total extracts with polyclonal anti-Set1 antibodies BL2,3 and BL2. Both antibodies immunoreacted with a single band of ≈130 kDa. (B) Western analysis of 17A1 total extracts on HW 65F size-exclusion chromatography column. Set1 complex eluted with a putative molecular mass of ≈1 MDa. (C) Chromatography scheme for the conventional biochemical purification of COMPASS: 17A1 total extract, 0–40% (NH4)2SO4, HW 65F, DEAE-5PW, Phenyl-5PW, SP-NPR, and Superose-6 PC. (D) Silver stain and Western analysis of purified COMPASS following our purification scheme. Silver stain and Western analysis of the final purified Set1 fraction indicated that Set1 copurified with six other polypeptides of ≈25, 30, 35, 40, 50, and 60 kDa. (E) Western analysis of Superose-6 PC size exclusion fractions. COMPASS eluted with a molecular mass of ≈1 MDa, indicating that no large polypeptides were lost from the COMPASS complex during the course of conventional purification.
Results
Set1 Is in a Large Macromolecular Complex.
We purified COMPASS by monitoring the purification using polyclonal antibodies generated against the SET domain of Set1 (a generous gift from L. Pillus). The two polyclonal antibodies reacted with a polypeptide with an approximate molecular mass of 130 kDa, the predicted mass of Set1 (Fig. 1A). Also, Western analysis of the extract derived from a strain deleted for Set1 did not demonstrate the presence of the same band (data not shown). Set1 elutes from a size-exclusion column with an approximate molecular mass of 1 MDa (Fig. 1B), suggesting that Set1 exists in a large macromolecular complex.
Conventional Purification of COMPASS.
Extract from ≈300 liters of yeast was subjected to column chromatography as shown in Fig. 1C, and fractions were tested for the presence of Set1 using the Set1 antibodies. Silver stain and Western blot analysis of fractions from the final size-exclusion chromatography step revealed that Set1 copurifies with six other polypeptides of about 25, 30, 35, 40, 50, and 60 kDa (Fig. 1D), which we have named COMPASS 25 (Cps25), COMPASS 30 (Cps30), COMPASS 35 (Cps35), COMPASS 40 (Cps40), COMPASS 50 (Cps50), and COMPASS 60 (Cps60). The recovery of this complex by conventional purification is about 3–5%. The solution molecular mass of the homogenously purified COMPASS was estimated to be about 1 MDa (Fig. 1E), approximately the same size as for the COMPASS in the total yeast extract (Fig. 1B), which suggests that we did not lose any large polypeptides from the complex during the process of purification.
Each of the proteins in COMPASS was identified by tandem and MALDI-TOF mass spectrometry. The protein Cps60 is the product of the BRE2/YLR015w gene, which encodes a protein similar to both Drosophila and human ASH2L proteins. ASH2L is a member of the trithorax family of homeodomain DNA-binding proteins that is thought to regulate gene expression, morphogenesis, and differentiation in humans (20, 21). Cps50 (Fig. 1D) is Yar003, which contains a WD (WD-40) domain. The human homologue of Cps50 (YAR003) binds to the retinoblastoma protein E1A-binding pocket B and also contains a WD domain (22). The WD-40 domains have been shown to be involved in mediating protein–protein interactions (23). Cps40 is the product of the YPL138 gene, which contains a conserved region found in human trithorax (MLL). Mammalian Cps40 homologue recognizes unmethylated CpG islands and is thought to function as a transcriptional activator (24). Cps35 (Fig. 1D) is YKL018, which contains several WD repeats. Cps35 (Ykl018) is the only essential subunit of the COMPASS. Cps30 is encoded by the YBR175 gene and also contains several WD-40 repeats. The smallest subunit of this complex that we can detect, Cps25, is encoded by the YDR469 gene. It has weak similarity to a protein of Caenorhabditis elegans (dpy-30) involved in dosage compensation of expression of genes on the X chromosome (25). Interestingly, two pairs of the COMPASS components, Cps30-Cps50 and Cps25-Cps60, have been found to interact in the yeast two-hybrid assay (26, 27). Although a seven-protein complex must have at least six protein–protein interactions, none of the other interactions has been detected in comprehensive two-hybrid screens.
TAP of COMPASS.
To confirm that the subunits of COMPASS are actually part of a complex, we purified COMPASS by using an affinity tag on Set1. Western blot analysis using antibodies specific to Set1 indicated that Set1 is present in the affinity tag-purified complex (Fig. 2B). We observed two forms of Set1 from the affinity purification (Fig. 2 A and B). It is possible that the second form of Set1 is either a degradation product of the larger form or a modified form of Set1. This point is not clear at this time. However, our study indicates that the ratio of the components of COMPASS appears to be 1:1. Tagged Set1 copurified with six polypeptides of approximately the same molecular weights as those initially found in COMPASS (Fig. 2A). MALDI-TOF analysis indicated that all of the polypeptides found in the conventionally purified COMPASS are also found in the Set1 complex purified using the TAP-affinity tag (compare Figs. 1D and 2A). Additionally, TAP-tag affinity purification of one of the COMPASS subunits (Cps60/Ylr015) yielded Set1 (Fig. 2D, identified by Western blotting with Set1 antibody) and all of the other components of COMPASS (compare Figs. 1D, 2A, and 2C) as identified by MALDI-TOF. We have also TAP-tag affinity-purified Cps40 and have demonstrated that the same complex (COMPASS) is purified with Cps40 (data not shown).
Figure 2.

TAP of COMPASS. (A) Silver stain analysis of TAP-tag affinity-purified Set1. Tagged Set1 copurified with six other polypeptides of ≈25, 30, 35, 40, 50, and 60 kDa, similar to the conventionally purified COMPASS. (B) Western analysis of affinity-purified tagged Set1. Anti-Set1 polyclonal antibody immunoreacted with affinity-purified Set1 tag. (C) Silver stain analysis of TAP-tag affinity-purified COMPASS 60. Tagged COMPASS 60 affinity-purified with six other polypeptides of ≈25, 30, 35, 40, 50, and 130 kDa, similar to the conventionally purified and COMPASS. (D) Western analysis of affinity-purified tagged COMPASS 60 with anti-Set1 polyclonal antibody. Set1 antibody immunoreacted with Set1 in the affinity-purified COMPASS 60 fraction.
Delayed Growth Phenotypes and Hydroxyurea Sensitivity of Null Mutants of the Nonessential Subunit COMPASS.
Mutants missing individual (nonessential) proteins of COMPASS, including Set1, were found to grow more slowly than wild-type cells on standard rich media when diluted for growth from stationary phase (Fig. 3A). In addition, COMPASS mutants are sensitive to hydroxyurea (Fig. 3B) even when plated from log-phase culture, a phenotype caused by mutations in other genes encoding chromatin-associated factors, such as RSC, INO80, and POB3 (28–30). Most recently, Geli and coworkers (31) have demonstrated that a set1 mutation unveils a novel signaling pathway relayed by the Rad53-dependent hyperphosphorylation of replication protein A. They also demonstrated that such a mutation in set1 leads to transcriptional activation of repair genes. Our observation that COMPASS mutants are sensitive to hydroxyurea may be explained by a role of set1 in Rad53-dependent pathways.
Figure 3.

Phenotypic characterization of cps deletion strains. (A) Slow-growth phenotype of cps and set1 deletion strains. Strains containing the indicated cps and set1 deletions were grown in liquid yeast extract peptone dextrose starting from overnight cultures. Time points indicate when an aliquot of cells was removed and the A600 nm was read. The strains harboring the COMPASS deletions are diploid strains that have both copies of the appropriate cps gene deleted. (B) Hydroxyurea sensitivity of cps deletion strains. Strains harboring the indicated cps deletions were plated from midlog growing cultures. Approximately 2,000 cells were plated onto YPD containing 100 mM hydroxyurea and grown for 2 days. Under these conditions, growth of all of the strains is similar when cells are plated in the absence of hydroxyurea (data not shown).
COMPASS Functions as a Complex in Vivo.
Because we expect a protein complex containing trithorax to be involved in the regulation of gene expression, we examined whether COMPASS is important for gene expression in vivo. Indeed, mutants missing either of two of the COMPASS components, Cps50 and Cps40, displayed similar changes in gene expression for a similar set of genes (Fig. 4C). A subset of about 20 genes had expression levels reduced at least 1.5-fold when the COMPASS deletion mutants were compared with the wild type (Fig. 4B). Also, a subset of about 30 genes had mRNA levels that were increased at least 1.5-fold when a COMPASS gene was deleted (Fig. 4A). Most important, the effects of the Cps50Δ and Cps40Δ mutations were highly correlated (Fig. 4C). Because the expression of the same subset of genes was altered by the mutations in the components of COMPASS, our data strongly suggest that the subunits of COMPASS function as a complex in vivo.
Figure 4.

The effects of cps50 and cps40 deletions on gene expression are similar. (A) Genes whose mRNA levels are down-regulated [cps50 (003) and cps40 (138)] at least 1.5-fold in both COMPASS deletion strains. The black box indicates that the effect of gene expression for that point is less than 1.5-fold. (B) Genes whose mRNA levels are up-regulated at least 1.5-fold in both COMPASS deletion strains. In each case (e.g., cpsΔ_wt), the first name listed corresponds to cDNA labeled with Cy5-dCTP, and the second represents cDNA that has been labeled with Cy3-dCTP. Equal amounts of the two types of cDNA were mixed and hybridized to the same array. After normalization, the results are always expressed as cpsΔ/wt. A black box indicates that the effect of the cps deletion on gene expression is less than 1.5-fold. (C) The mRNA level for each gene in a cps deletion strain was calculated, after normalization, relative to that of the same gene in a wild-type strain. Using scatter plots in which each point represents one gene, these data were then used to graphically compare the effect of cps50 (003) deletion with the effects of cps40 (138) deletion.
Similar to Set1, Mutations in the COMPASS Components Cause a Defect in Silencing of Gene Expression at Telomeres.
Set1 mutants have defects in silencing gene expression near chromosomal telomeres (32). If the components of COMPASS function as a complex with Set1 in vivo, their inactivation would be expected to result in relief of silencing of gene expression near telomeres. The genes encoding the compass components shown in Fig. 5 were deleted in a strain carrying URA3 located near telomeres (ucc1001) following a previously published method (32). Five-fold serial dilutions were applied to SD-ura (Fig. 5 Left) and SD complete plate (Fig. 5 Right). Growth was compared with that of parental stain ucc1001 after 48 h (Fig. 5). Our results indicate that the inactivation of any of the COMPASS components (as well as a protein—Fyv9—that is thought to interact with one of the COMPASS components) relieves silencing of a URA3 gene residing near the telomere of chromosome VII. This phenotype of the COMPASS subunits is similar to that of Set1 mutants, again suggesting that COMPASS subunits are part of a functional complex with Set1 in vivo (32).
Figure 5.

Deletion of all of the subunits of COMPASS results in a defect in silencing of gene expression at telomeres. The nonessential subunits of COMPASS (CPS60, 50, 40, 30, and 25) were deleted in the URA3 telomeric reporter strain ucc1001. Five-fold serial dilutions from 5 × 104 − 500 cells were applied to SD-ura (Left) and SD complete plate (Right). After 48 h, growth was compared with that of parental stain ucc1001.
Discussion
Set1 and Set2 are the two proteins in yeast most similar to the Drosophila and human trithorax proteins (trithorax and MLL, respectively) (32). Very little is known about the function of any of the trithorax-related proteins. The trithorax protein is a putative DNA-binding protein that seems to be a positive regulator of gene expression. In humans, mutations and translocations within the mammalian trithorax gene (MLL) result in the development of hematological malignancies. Our characterization of the protein complex we call COMPASS is a step toward understanding the function of trithorax-related SET domain-containing proteins.
Only one of the COMPASS components, Cps35/Ykl018, is essential for yeast growth. It is conceivable that this protein may also be a component of a different complex. Null mutants missing any of the six nonessential subunits of COMPASS display delayed growth on rich medium as well as sensitivity to hydroxyurea. Whereas sensitivity to this compound is usually taken as evidence for a role of the missing protein in DNA replication or its regulation, recent work has shown that mutations in genes that encode factors involved in transcription and/or chromatin remodeling, such as Pob3, Rsc1, and Ino80, also cause sensitivity to hydroxyurea (28–30). Our microarray analysis shows that removal of either of two components of COMPASS has a similar effect on gene expression, suggesting that these polypeptides function together in vivo. Also, our results indicate that the inactivation of any of the COMPASS components relieves silencing of a URA3 gene residing near the telomere. This phenotype of the COMPASS subunit mutants is similar to that of set1 mutants. Together, these results suggest that COMPASS functions with Set1 as a complex in vivo.
Whereas isolation of COMPASS from S. cerevisiae should enable us to clarify the roles of the SET domain proteins in yeast, identification of Set1-interacting proteins of S. cerevisiae should also allow us to identify their homologue complexes from Drosophila and mammals. Several subunits of COMPASS are trithorax-related proteins, including the Set1 protein, which is the yeast homologue to the Drosophila and mammalian trithorax. Cps60/Bre2/Ylr015 has weak similarity to both the Drosophila and human ASH2 proteins, which belong to the trithorax gene family and has a nuclear localization signal and a PHD finger motif. Ash2 positively regulates expression of homeotic selector genes and is implicated in early development and formation of various disk patterns. Cps40/Ypl138 is also similar to trithorax family members, and the mammalian homologue of Cps40/Ypl138 is thought to be a transcriptional activator containing a conserved region found in human trithorax MLL.
It has been postulated that the trithorax proteins function in chromatin remodeling and transcriptional activation through interactions with DNA. COMPASS 40 (Cps40/Ypl138) is similar to a human CpG-binding protein (hCGBP), which recognizes unmethylated CpG and has been suggested to be involved in transcriptional activation (24). Although we detected a CpG-dependent gel shift of the same DNA sequence reported by Voo et al. (24) using HeLa cell nuclear extracts, we were unable to detect binding to this sequence by purified COMPASS (data not shown). Perhaps Cps40/Ypl138 and its mammalian homologue have different sequence specificities for DNA binding. This can be examined directly.
Several SET domain-containing proteins [notably, human SUV39H1 and mouse Suv39h1, mammalian homologues of Drosophila Su(var)3–9 and Schizosaccharomyces pombe clr4] were demonstrated to methylate lysine 9 of histone H3 in vitro (33). We have tested Set1 for the possible methyltransferase activity and do not detect any methyltransferase activity associated with COMPASS from either free histone or nucleosomes (data not shown). It is, however, significant that a subunit of COMPASS (Cps50) interacts in the yeast two-hybrid assay with a Cps50/Yar003 protein containing a motif characteristic of S-adenosylmethionine-dependent methyltransferases. Although this putative methyltransferase was not found in any of our purified COMPASS complexes, it is possible that it dissociated from COMPASS during the process of purification. Because this methyltransferase is a small protein, its dissociation from the complex might not have been detected by size-exclusion chromatography (Fig. 1).
This molecular and biochemical purification and identification of a trithorax-containing complex will open the way for biochemical studies regarding the cellular function of this class of proteins in development. Also, because the mammalian trithorax protein has been implicated in a number of hematological malignancies (34), characterization of all of the subunits of a trithorax complex should be helpful in defining the role of this class of proteins in regulation of gene expression and how their misregulation results in development of human cancer.
Figure 6.
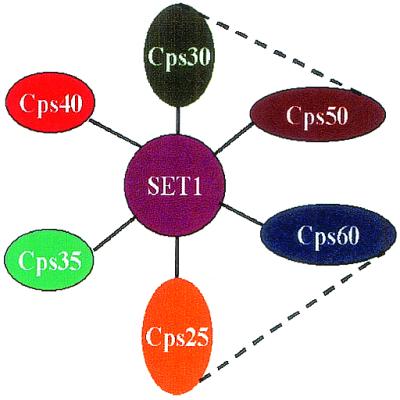
The proposed structure of COMPASS. Graphic representation of COMPASS. Solid bars between the polypeptides of COMPASS are determined by either conventional or affinity purifications. The dashed bars indicate interactions determined by yeast two-hybrid.
Acknowledgments
We are grateful to Dr. Lorraine Pillus for her generous gift of antibody to Set1. We thank Dr. Dale Dorsett for technical advice and assistance. We also thank G. Zhong for excellent technical support. N.J.K. was supported by a PGS-B Scholarship Award from the Natural Sciences and Engineering Research Council of Canada. This work was supported in part by grants from the Medical Research Council of Canada and the National Cancer Institute of Canada with funds from the Canadian Cancer Society (to J.F.G.) and American Cancer Society Grant RP69921801, National Institutes of Health Grant 1R01CA089455, and a Mallinckrodt Foundation Award (to A.S.). Work in the laboratory of M.J. was supported by the James S. McDonnell Foundation. J.F.G. is an International Research Scholar of the Howard Hughes Medical Institute. A.S. is a scholar of the Leukemia and Lymphoma Society.
Abbreviations
- MALDI-TOF
matrix-assisted laser desorption ionization–time-of-flight
- TAP
tandem affinity purification
References
- 1.Pirrotta V. Cell. 1998;93:333–336. doi: 10.1016/s0092-8674(00)81162-9. [DOI] [PubMed] [Google Scholar]
- 2.Mahmoudi T, Verrijzer C P. Oncogene. 2001;20:3055–3066. doi: 10.1038/sj.onc.1204330. [DOI] [PubMed] [Google Scholar]
- 3.Stassen M J, Bailey D, Nelson S, Chinwalla V, Harte P J. Mech Dev. 1995;52:209–223. doi: 10.1016/0925-4773(95)00402-m. [DOI] [PubMed] [Google Scholar]
- 4.Jenuwein T, Laible G, Dorn R, Reuter G. Cell Mol Life Sci. 1998;54:80–93. doi: 10.1007/s000180050127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jones R S, Gelbart W M. Mol Cell Biol. 1993;13:6357–6366. doi: 10.1128/mcb.13.10.6357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tschiersh B, Hofmann A, Krauss V, Dorn R, Korge G, Reuter G. EMBO J. 1994;13:3822–3831. doi: 10.1002/j.1460-2075.1994.tb06693.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.LaJeunesse D, Shearn A. Mech Dev. 1995;53:123–139. doi: 10.1016/0925-4773(95)00430-0. [DOI] [PubMed] [Google Scholar]
- 8.Gellon G, Harding K W, McGinnis N, Martin M M, McGinnis W. Development (Cambridge, UK) 1997;124:3321–3331. doi: 10.1242/dev.124.17.3321. [DOI] [PubMed] [Google Scholar]
- 9.Jones R S, Gelbart W M. Genetics. 1990;126:185–199. doi: 10.1093/genetics/126.1.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pelegri F, Lehmann R. Genetics. 1994;136:1341–1353. doi: 10.1093/genetics/136.4.1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hanson R D, Hess J L, Yu B D, Ernst P, van Lohuizen M, Berns A, van der Lugt N M, Shashikant C S, Ruddle F H, Seto M, Korsmeyer S J. Proc Natl Acad Sci USA. 1999;96:14372–14377. doi: 10.1073/pnas.96.25.14372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Laible G, Haynes A R, Lebersorger A, O'Carroll D, Mattei M G, Denny P, Brown S D, Jenuwein T. Mamm Genome. 1999;10:311–314. doi: 10.1007/s003359900993. [DOI] [PubMed] [Google Scholar]
- 13.O'Carroll D, Erhardt S, Pagani M, Barton S C, Surani M A, Jenuwein T. Mol Cell Biol. 2001;21:4330–4336. doi: 10.1128/MCB.21.13.4330-4336.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rowley J D. Annu Rev Genet. 1998;32:495–519. doi: 10.1146/annurev.genet.32.1.495. [DOI] [PubMed] [Google Scholar]
- 15.Puig O, Caspary F, Rigaut G, Rutz B, Bouveret E, Bragado-Nilsson E, Wilm M, Seraphin B. Methods. 2001;24:218–229. doi: 10.1006/meth.2001.1183. [DOI] [PubMed] [Google Scholar]
- 16.Rigaut G, Shevchenko A, Rutz B, Wilm M, Mann M, Seraphin B. Nat Biotechnol. 1999;17:1030–1032. doi: 10.1038/13732. [DOI] [PubMed] [Google Scholar]
- 17.Mann M, Hojrup P, Roepstorff P. Biol Mass Spectrom. 1993;22:338–345. doi: 10.1002/bms.1200220605. [DOI] [PubMed] [Google Scholar]
- 18.Erdjument-Bromage H, Lui M, Lacomis L, Grewal A, Annan R S, MacNulty D E, Carr S A, Tempst P. J Chromatogr. 1998;826:167–181. doi: 10.1016/s0021-9673(98)00705-5. [DOI] [PubMed] [Google Scholar]
- 19.Geromanos S, Freckleton G, Tempst P. Anal Chem. 2000;72:777–790. doi: 10.1021/ac991071n. [DOI] [PubMed] [Google Scholar]
- 20.Adamson A L, Shearn A. Genetics. 1996;144:621–633. doi: 10.1093/genetics/144.2.621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ikegawa S, Isomura M, Koshizuka Y, Nakamura Y. Cytogenet Cell Genet. 1999;84:167–172. doi: 10.1159/000015248. [DOI] [PubMed] [Google Scholar]
- 22.Saijo M, Sakai Y, Kishino T, Niikawa N, Matsuura Y, Morino K, Tamai K, Taya Y. Genomics. 1995;27:511–519. doi: 10.1006/geno.1995.1084. [DOI] [PubMed] [Google Scholar]
- 23.Smith T F, Gaitatzes C, Saxena K, Neer E J. Trends Biochem Sci. 1999;24:181–185. doi: 10.1016/s0968-0004(99)01384-5. [DOI] [PubMed] [Google Scholar]
- 24.Voo K S, Carlone D L, Jacobsen B M, Flodin A, Skalnik D G. Mol Cell Biol. 2000;20:2108–2121. doi: 10.1128/mcb.20.6.2108-2121.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hsu D R, Chuang P T, Meyer B J. Development (Cambridge, UK) 1995;121:3323–3334. doi: 10.1242/dev.121.10.3323. [DOI] [PubMed] [Google Scholar]
- 26.Uetz P, Giot L, Cagney G, Mansfield T A, Judson R S, Knight J R, Lockshon D, Narayan V, Srinivasan M, Pochart P, et al. Nature (London) 2000;403:623–627. doi: 10.1038/35001009. [DOI] [PubMed] [Google Scholar]
- 27.Ito T, Chiba T, Ozawa R, Yoshida M, Hattori M, Sakaki A. Proc Natl Acad Sci USA. 2001;98:4569–4574. doi: 10.1073/pnas.061034498. . (First Published March 13, 2001; 10.1073/pnas.061034498) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cairns B R, Schlichter A, Erdjument-Bromage H, Tempst P, Kornberg R D, Winston F. Mol Cell. 1999;4:715–723. doi: 10.1016/s1097-2765(00)80382-2. [DOI] [PubMed] [Google Scholar]
- 29.Shen X, Mizuguchi G, Hamiche A, Wu C. Nature (London) 2000;406:541–544. doi: 10.1038/35020123. [DOI] [PubMed] [Google Scholar]
- 30.Schlesinger M B, Formosa T. Genetics. 2000;155:1593–1606. doi: 10.1093/genetics/155.4.1593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Schramke V, Neecke H, Brevet V, Corda Y, Lucchini G, Longhese M P, Gilson E, Geli V. Genes Dev. 2001;15:1845–1858. doi: 10.1101/gad.193901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nislow C, Ray E, Pillus L. Mol Biol Cell. 1997;8:2421–2436. doi: 10.1091/mbc.8.12.2421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rea S, Eisenhaber F, O'Carroll D, Sun Z, Schmid M, Opravil S, Mechtler K, Ponting C P, Allis C D, Junuwein T. Nature (London) 2000;406:593–599. doi: 10.1038/35020506. [DOI] [PubMed] [Google Scholar]
- 34.Shilatifard A, Lane W S, Jackson K W, Conaway R C, Conawy J W. Science. 1996;271:1873–1876. doi: 10.1126/science.271.5257.1873. [DOI] [PubMed] [Google Scholar]