

Abstract
The genus Seiridium includes multiple plant pathogenic fungi well-known as causal organisms of cankers on Cupressaceae. Taxonomically, the status of several species has been a topic of debate, as the phylogeny of the genus remains unresolved and authentic ex-type cultures are mostly absent. In the present study, a large collection of Seiridium cultures and specimens from the CBS and IMI collections was investigated morphologically and phylogenetically to resolve the taxonomy of the genus. These investigations included the type material of the most important Cupressaceae pathogens, Seiridium cardinale, S. cupressi and S. unicorne. We constructed a phylogeny of Seiridium based on four loci, namely the ITS rDNA region, and partial translation elongation factor 1-alpha (TEF), β-tubulin (TUB) and RNA polymerase II core subunit (RPB2). Based on these results we were able to confirm that S. unicorne and S. cupressi represent different species. In addition, five new Seiridium species were described, S. cupressi was lectotypified and epitypes were selected for S. cupressi and S. eucalypti.
Keywords: appendage-bearing conidia, canker pathogen, Cupressus, pestalotioid fungi, systematics
INTRODUCTION
The genus Seiridium (Sordariomycetes, Xylariales, Sporocadaceae) comprises a variety of mainly plant pathogenic fungi (Boesewinkel 1983, Graniti 1986, 1998, Barnes et al. 2001, Tsopelas et al. 2007). The genus was established based on Seiridium marginatum (Nees 1817), collected from rose stems in Germany and recently epitypified by Jaklitsch et al. (2016). The Sporocadaceae includes acervular asexual-morphs producing distinctive appendaged (pestalotioid) conidia (Jaklitsch et al. 2016), including other commonly known appendaged genera such as Bartalinia, Pestalotiopsis and Seimatosporium. Morphologically, Seiridium is distinguishable by its 5-septate conidia. Based on this characteristic, Maharachchikumbura et al. (2014) suggested that the monotypic genus Pestalotia, accommodating P. pezizoides (De Notaris 1841), might also be a synonym of Seiridium.
Seiridium is particularly known for its plant pathogenic species which have manifested profound economic damage globally. Presently, three Seiridium species (S. cardinale, S. cupressi and S. unicorne) are considered responsible for a pandemic of cypress canker disease, impacting plantations for wood production and ornamental tree cultivation (Boesewinkel 1983, Graniti 1986, 1998). Infection of Cupressaceae usually occurs secondary to tissue wounds produced by various agents, including wind, frost and insects, upon which the cankers appear as necrotic lesions on the tree bark. Once infected, cell necrosis of cypress tissue progresses steadily, reaching the cortical parenchyma, the phloem and the cambium, eventually causing the plant to die (Graniti 1998). Of these three species, Seiridium cardinale is the most aggressive and was first identified in California, from where the disease has since spread to other continents. Currently, the disease is particularly advanced in the Mediterranean region (Xenopoulos & Diamandis 1985, Graniti 1993, 1998, Della Rocca et al. 2011, 2013). The other two causal agents of cypress canker, S. cupressi and S. unicorne, are less destructive, but are considered responsible for disease in cypress plantations in East Africa and Japan, respectively (Nattrass & Ciccarone 1947, Jones 1953, 1954a, b, Tabata 1991). Whereas S. cardinale and S. cupressi appear to be restricted to Cupressaceae, S. unicorne has been reported from a range of plant families, including Anacardiaceae, Caprifoliaceae, Cornaceae, Cupressaceae, Hamamelidaceae, Rosaceae and Vitaceae (Guba 1961, Boesewinkel 1983, Cho & Shin 2004).
While S. cardinale is morphologically easily distinguishable from the other two putative Cupressus pathogens, S. cupressi and S. unicorne share more similarities and have, depending on the author, been treated as one or separate species (Guba 1961, Natrass et al. 1963, Swart 1973, Sutton 1975, Boesewinkel 1983, Graniti 1986, Nag Raj 1993, Viljoen et al. 1993). Seiridium unicorne (as Pestalozzia unicornis) was originally described from Chamaecyparis thyoides in New Jersey (Cooke & Ellis 1878). Guba (1961) introduced S. cupressi as Cryptostictis cupressi from multiple specimens of Cupressus macrocarpa collected in East Africa. Based on sequence data, both species are presently accepted (Barnes et al. 2001), and considered to be morphologically highly variable (Boesewinkel 1983, Graniti 1986, Chou 1989). Apart from intraspecific morphological variation, strains of the same species also vary in aggressiveness. Guba (1961) used both herbarium and culture material for the description of S. cupressi. The two cited herbarium materials were collected from Cupressus macrocarpa by Nattrass and the cultures by Jones (1953, 1954a) from Cupressus and Juniperus, who had studied their pathogenicity and, based on this feature, distinguished different strains. Although the cultures are still available, they have not been included in previously published phylogenetic analyses addressing the identity of S. unicorne and S. cupressi (Barnes et al. 2001, Jeewon et al. 2002, Cunnington 2007, Tsopelas et al. 2007, Maharachchikumbura et al. 2015). Moreover, since Guba (1961) did not designate a holotype specimen, a lectotype needs to be selected for S. cupressi. Furthermore, it is currently unclear whether these original specimens truly represent a single phylogenetic lineage. Similarly, no DNA sequence information has been generated thus far for the type of S. unicorne. However, a holotype specimen exists, and it can be epitypified if an appropriate culture is available. Previous studies using genetic markers to construct phylogenies (Barnes et al. 2001, Cunnington 2007, Tsopelas et al. 2007), as well as the restriction length polymorphism assay for Seiridium spp. identification (Krokene et al. 2004), did not include the type specimens and have relied on morphological identification of S. unicorne and S. cupressi. It remains therefore debatable if the clades identified in these phylogenies as S. unicorne and S. cupressi truly represent the lineages in which the type specimens would nest. To resolve this problem, the original materials of S. unicorne and S. cupressi need to be re-examined and if possible lecto- and epitypified in combination with analysis of DNA sequence data. In addition, to clarify the taxonomic identity of cultures and specimens from earlier studies (Barnes et al. 2001, Krokene et al. 2004, Cunnington 2007, Tsopelas et al. 2007), which are presently available in the public collections, a phylogenetic overview of the genus is required.
Seiridium is not solely comprised of Cupressaceae pathogens. Cankers inflicted by Seiridium spp. have been observed in Eucalyptus spp. (Myrtaceae) (Yuan & Old 1995, Yuan & Mohammed 1997, 1999, 2001), and recently, S. phylicae was described (Crous et al. 2012), which is an aggressive pathogen of Phylica arborea (Rhamnaceae), endemic to the archipelago Tristan da Cunha (Ryan et al. 2014). Furthermore, less destructive and perhaps more opportunistic pathogens are common to the genus, such as the generic type S. marginatum, for example.
The most well-studied sister genus of Seiridium, namely Pestalotiopsis, is known to include many endophytes (Maharachchikumbura et al. 2014). Pestalotiopsis has received particular interest due to the surprising high number of bioactive compounds produced by different species (Xu et al. 2010), and the record number of secondary metabolite clusters that was detected in the recently sequenced genome of Pestalotiopsis fici (Wang et al. 2015). Although poorly investigated, bioactive compounds have been isolated from Seiridium spp. as well, including phytotoxins (Ballio et al. 1991, Graniti et al. 1992, Evidente et al. 1993). Being perhaps the most aggressive genus of plant pathogens in the Sporocadaceae, Seiridium might be considered a unique evolutionary lineage within the family and consequently a unique lineage with regard to novel metabolite production.
In the present study we aimed to investigate the DNA phylogeny of the genus Seiridium, and to epitypify species for which no ex-type culture is currently available. Furthermore, we aimed to identify the clade or clades representing the important Cupressaceae pathogens S. cardinale, S. cupressi and S. unicorne, and to resolve the dubiety surrounding their taxonomy. To this end we obtained all Seiridium strains available in the CBS and CPC collections (housed at the Westerdijk Fungal Biodiversity Institute, Utrecht, the Netherlands) supplemented with additional collections, holotype materials and other herbarium specimens from Kew Royal Botanic Gardens (IMI, CABI Biosciences, Egham, Surrey, England). We constructed a multi-gene phylogeny, using the internal transcribed spacer (ITS) region and partial β-tubulin (TUB), which are the DNA markers commonly sequenced for Seiridium, supplemented with the translation elongation factor 1-alpha (TEF) and RNA polymerase II core subunit (RPB2) gene, including available Seiridium sequences from previously published phylogenies (Barnes et al. 2001, Cunnington 2007, Tsopelas et al. 2007, Maharachchikumbura et al. 2015).
MATERIALS AND METHODS
Isolates and specimens
All Seiridium isolates present in the CBS and CPC collections were included in the present study. The ex-epitypes of Bartalinia robillardoides (CBS 122705), Neopestalotiopsis protearum (CBS 114178) and Seimatosporium rosae (CBS 139823) were used as outgroups in the phylogenetic analyses (Table 1). Sequences from other strains, not examined here but published in previous phylogenetic studies of the genus (Barnes et al. 2001, Jeewon et al. 2002, Cunnington 2007, Tsopelas et al. 2007, Maharachchikumbura et al. 2015, Wijayawardene et al. 2016), were retrieved from GenBank (Table 1). In addition, herbarium material was requested from IMI (housed at Kew), including the holotypes of S. cardinale and S. unicorne and the authentic material Guba (1961) used to describe S. cupressi.
Table 1.
Specimens and corresponding accession numbers used in the phylogenetic analysis.
| Species | Collection accession No.1 | Host/Substrate | Host family | Location | Clade No.2 | GenBank accession No.3 |
|||
|---|---|---|---|---|---|---|---|---|---|
| ITS | RPB2 | TEF | TUB | ||||||
| Seiridium camelliae | MFLUCC 12-0647T | Camellia reticula | Theaceae | China | 12 | JQ683725 | – | JQ683741 | JQ683709 |
| S. cancrinum | IMI 52256T; BRL 1119; CBS 226.55 | Cupressus macrocarpa | Cupressaceae | Kenya | 5 | LT853089 | LT853137 | LT853186 | LT853236 |
| CBS 907.85; CMW 320 | Cupressus lustinaca | Cupressaceae | South Africa | LT853090 | LT853138 | LT853187 | LT853237 | ||
| S. cardinale | CBS 123911 | Cupressus sempervirens | Cupressaceae | Morocco | 1 | LT853057 | LT853106 | LT853154 | LT853204 |
| CBS 122612; CMW 2133 | Cupressus sp. | Cupressaceae | Chile | LT853058 | LT853107 | LT853155 | LT853205 | ||
| CBS 172.56 | unknown | unknown | unknown | LT853059 | LT853108 | LT853156 | LT853206 | ||
| CBS 280.93; CPC 23780 | Cupressus sp. | Cupressaceae | Italy | LT853060 | LT853109 | LT853157 | LT853207 | ||
| CBS 522.82; CPC 23784; NBRC 32683 | Cupressus sempervirens | Cupressaceae | New Zealand | LT853061 | LT853110 | LT853158 | LT853208 | ||
| CBS 523.82; CPC 23793 | Cupressocyparis sp. | Cupressaceae | New Zealand | LT853062 | LT853111 | LT853159 | LT853209 | ||
| CBS 908.85; CPC 23779; CMW 616 | Cupressus lustinaca | Cupressaceae | South Africa | LT853063 | LT853112 | LT853160 | LT853210 | ||
| CBS 909.85R; CMW 635 | Cupressus lustinaca | Cupressaceae | South Africa | LT853064 | LT853113 | LT853161 | LT853211 | ||
| CBS 910.85; CPC 23781; CMW 600; CMW 638 | Cupressus sempervirens | Cupressaceae | South Africa | LT853065 | LT853114 | LT853162 | LT853212 | ||
| CMW 1644 | Cupressus sp. | Cupressaceae | Italy | – | – | – | AF320497 | ||
| CMW 1645 | Cupressus sp. | Cupressaceae | Italy | – | – | – | AF320501 | ||
| CMW 18602 | Cupressus sempervirens | Cupressaceae | Greece | – | – | – | DQ926974 | ||
| CMW 18603 | Cupressus macrocarpa | Cupressaceae | Greece | – | – | – | DQ926975 | ||
| CMW 18604 | Juniperus foetidissima | Cupressaceae | Greece | – | – | – | DQ926973 | ||
| CMW 18605 | Juniperus foetidissima | Cupressaceae | Greece | – | – | – | DQ926976 | ||
| CMW 18606 | Juniperus exelsa | Cupressaceae | Greece | – | – | – | DQ926978 | ||
| CMW 18794 | Cupressus sempervirens | Cupressaceae | Greece | – | – | – | DQ926977 | ||
| CPC 23785 | Cupressus sempervirens | Cupressaceae | France | LT853066 | LT853115 | LT853163 | LT853213 | ||
| CPC 23787 | Juniperus phoenica | Cupressaceae | Greece | LT853067 | LT853116 | LT853164 | LT853214 | ||
| CPC 23788 | Cupressus sempervirens | Cupressaceae | Italy | LT853068 | LT853117 | LT853165 | LT853215 | ||
| CPC 23790 | Cupressus sempervirens | Cupressaceae | Italy | LT853069 | LT853118 | LT853166 | LT853216 | ||
| CPC 23791 | Cupressus sempervirens | Cupressaceae | Italy | LT853070 | LT853119 | LT853167 | LT853217 | ||
| CPC 23792 | Cupressus sempervirens | Cupressaceae | Italy | – | LT853071 | LT853168 | LT853218 | ||
| CPC 23794 | unknown | unknown | Greece | LT853072 | LT853120 | LT853169 | LT853219 | ||
| CPC 23795 | unknown | unknown | Greece | LT853073 | LT853121 | LT853170 | LT853220 | ||
| CPC 23796 | Cupressus sempervirens | Cupressaceae | Italy | LT853074 | LT853122 | LT853171 | LT853221 | ||
| CPC 23797 | Cupressus sempervirens | Cupressaceae | Italy | LT853075 | LT853123 | LT853172 | LT853222 | ||
| CPC 23798 | Cupressus sempervirens | Cupressaceae | Italy | LT853076 | LT853124 | LT853173 | LT853223 | ||
| CPC 28286 | Cupressus sempervirens | Cupressaceae | Spain | LT853077 | LT853125 | LT853174 | LT853224 | ||
| CPC 28287 | Cupressus sempervirens | Cupressaceae | Spain | LT853078 | LT853126 | LT853175 | LT853225 | ||
| S. ceratosporum | PHSI2001Pathcw07 | Vitis vinifera | Vitaceae | China | 16 | AY687314 | – | – | DQ534043 |
| S. cupressi | CBS 122616; CMW 1646 | Cupressus sp. | Cupressaceae | Greece | 3 | LT853082 | LT853130 | LT853179 | LT853229 |
| IMI 52254L; BRL 1117; CBS 224.55ET | Cupressus macrocarpa | Cupressaceae | Kenya | LT853083 | LT853131 | LT853180 | LT853230 | ||
| IMI 52255; BRL 1118; CBS 225.55 | Cupressus forbesii | Cupressaceae | Kenya | LT853084 | LT853132 | LT853181 | LT853231 | ||
| IMI 52258; BRL 1120; CBS 227.55 | Cupressus macrocarpa | Cupressaceae | Uganda | LT853085 | LT853133 | LT853182 | LT853232 | ||
| CBS 320.51 | Cupressus sp. | Cupressaceae | Kenya | LT853086 | LT853134 | LT853183 | LT853233 | ||
| CMW 18607 | Cupressus sempervirens | Cupressaceae | Greece | – | – | – | DQ926979 | ||
| S. eucalypti | CBS 343.97ET; CMW 5303 | Eucalyptus delegatensis | Myrtaceae | Australia | 10 | LT853099 | LT853146 | LT853196 | LT853246 |
| S. kartense | CBS 142629T; CPC 20183 | Eucalyptus cladocalyx | Myrtaceae | Australia | 11 | LT853100 | LT853147 | LT853197 | LT853247 |
| S. kenyanium | IMI 52257T; BRL 1121; CBS 228.55 | Juniperus procera | Cupressaceae | Kenya | 9 | LT853098 | LT853145 | LT853195 | LT853245 |
| S. marginatum | CBS 140403GT | Rosa canina | Rosaceae | France | 15 | KT949914 | LT853149 | LT853199 | LT853249 |
| S. neocupressi | CMW 420 | Cupressus macrocarpa | Cupressaceae | New Zealand | 2 | – | – | – | AF320487 |
| CMW 5282; ATCC 48158 | Cupressus sp. | Cupressaceae | New Zealand | FJ430600 | – | – | AF320489 | ||
| CBS 142625T; CPC 23786 | Cupressus sempervirens | Cupressaceae | Italy | LT853079 | LT853127 | LT853176 | LT853226 | ||
| CBS 142626; CPC 23789 | Cupressus sempervirens | Cupressaceae | Italy | LT853080 | LT853128 | LT853177 | LT853227 | ||
| CBS 142627; CPC 28351 | Cupressus leylandii | Cupressaceae | Australia | LT853081 | LT853129 | LT853178 | LT853228 | ||
| VPRI 15696 | Cupressus sp. | Cupressaceae | Australia | – | – | – | EF517787 | ||
| VPRI 16083 | Cupressus sp. | Cupressaceae | Australia | – | – | – | EF517786 | ||
| VPRI 32740 | Cupressus sp. | Cupressaceae | Australia | – | – | – | EF517789 | ||
| VPRI 40658 | Cupressus sp. | Cupressaceae | Australia | – | – | – | EF517790 | ||
| VPRI 40665 | Cupressus sp. | Cupressaceae | Australia | – | – | – | EF517788 | ||
| S. papillatum | CBS 340.97T; VPRI 20827; CMW 5302 | Eucalyptus delegatensis | Myrtaceae | Australia | 17 | LT853102 | LT853150 | LT853200 | LT853250 |
| S. phylicae | CBS 133587T; CPC 19964 | Phylica arborea | Rhamnaceae | Tristan da Cunha | 6 | LT853091 | LT853139 | LT853188 | LT853238 |
| CPC 19962 | Phylica arborea | Rhamnaceae | Tristan da Cunha | LT853092 | LT853140 | LT853189 | LT853239 | ||
| CPC 19965 | Phylica arborea | Rhamnaceae | Tristan da Cunha | LT853093 | LT853141 | LT853190 | LT853240 | ||
| CPC 19970 | Phylica arborea | Rhamnaceae | Tristan da Cunha | LT853094 | – | LT853191 | LT853241 | ||
| S. podocarpi | CBS 137995T; CPC 23429 | Podocarpus latifolus | Podocarpaceae | South Africa | 13 | LT853101 | LT853148 | LT853198 | LT853248 |
| S. pseudocardinale | MFLUCC 13-0525T | Cupressus arizonica | Cupressaceae | Italy | 8 | KU848210 | – | – | – |
| CBS 122613; CMW 1648 | Cupressus sp. | Cupressaceae | Portugal | LT853096 | LT853143 | LT853193 | LT853243 | ||
| CBS 122614; CMW 1649 | Cupressus sp. | Cupressaceae | Portugal | LT853097 | LT853144 | LT853194 | LT853244 | ||
| S. spyridicola | CBS 142628T; CPC 29108 | Spyridium globosum | Rhamnaceae | Australia | 7 | LT853095 | LT853142 | LT853192 | LT853242 |
| S. unicorne | CBS 120306; CMW 5596 | Cupressus sempervirens | Cupressaceae | South Africa | 4 | LT853087 | LT853135 | LT853184 | LT853234 |
| CBS 538.82R; NBRC 32684; CMW 5443 | Cryptomeria japonica | Cupressaceae | New Zealand | LT853088 | LT853136 | LT853185 | LT853235 | ||
| S. venetum | MFLU 15-0369R | Cornus mas | Cornaceae | Italy | 14 | KT438836 | – | – | KT438837 |
| Neopestalotiopsis protearum | CBS 114178GT | Leucospermum cuneiforme | Proteaceae | Zimbabwe | Outgroup | LT853103 | LT853151 | LT853201 | LT853251 |
| Bartalinia robillardoides | CBS 122705GT | Leptoglossus occidentalis | Coreidae (Animalia) | Italy | Outgroup | LT853104 | LT853152 | LT853202 | LT853252 |
| Seimatosporium rosae | CBS 139823GT; MFLUCC 14-0621 | Rosa sp. | Rosaceae | Russia | Outgroup | LT853105 | LT853153 | LT853203 | LT853253 |
1 ATCC: American Type Culture Collection; BRL: Bristol City Library, Bristol, England, UK; CBS: Westerdijk Fungal Biodiversity Institute (formerly CBS-KNAW Fungal Biodiversity Centre), Utrecht, the Netherlands; CMW: Culture collection of the Forestry and Agricultural Biotechnology Institute (FABI), University of Pretoria, South Africa; CPC: Culture collection of P.W. Crous, held at the Westerdijk Fungal Biodiversity Institute, Utrecht, the Netherlands; IMI: Kew Royal Botanic Gardens, CABI Biosciences, Egham, Surrey, England; MFLU and MFLUCC: Mae Fah Luang University, Chiang Rai, Thailand; NBRC: National Institute of Technology and Evaluation, Kisarazu-shi, Chiba, Japan; PHSI: from Liu et al. (2007); VPRI: Victorian Department of Environment and Primary Industries, Knoxfield, Victoria, Australia.
2 Clade numbers correspond to the multi-gene phylogeny in Fig. 1 and single gene phylogenies in Fig. 2.
3 ITS: Internal transcribed spacer regions 1 and 2 and 5.8S; RPB2: RNA polymerase II core subunit; TEF: Translation elongation factor 1-alpha; TUB: Beta-tubulin.
T type or ex-type; ET ex-epitype; L lectotype; GT ex-generic type; R reference strain. Accession numbers of sequences generated in this study are in bold.
Culture preparation and morphology analysis
Lyophilized cultures from the CBS and CPC collections were reactivated on Petri dishes with 2 % malt extract agar (MEA) and isolates from the CBS collection stored in liquid nitrogen were revived on either MEA or oatmeal agar (OA). Culture characteristics were studied on Petri dishes containing cornmeal agar (CMA), MEA, potato dextrose agar (PDA), and synthetic nutrient-poor agar (SNA). All culture media were prepared according to recipes as in Crous et al. (2009). Cultures were grown at 22 °C with a 12 h daylight/darkness photoperiod for 2 wk. Colony colours were assessed using colour charts from Rayner (1970). Conidiomatal morphology was examined on both PDA and SNA and all other morphological analyses were conducted from colonies on SNA using Differential Interference Contrast (DIC) optics on a Nikon Eclipse 80i microscope and a Nikon AZ100 dissecting microscope, both equipped with Nikon DS-Ri2 high definition digital cameras. To visualize relevant morphological features of multiple conidia on composite photo plates, separate photographs of conidia were combined using Photoshop CS5.1. Measurements were made from microphotographs using the Nikon NIS-elements D v. 4.50 software. Morphology of conidiophores, conidiogenous cells and conidia was examined and measured from colonies grown on SNA. For the descriptions, a minimum of 20 measurements was taken for conidiophores and conidiogenous cells, and for cell dimensions of conidia a minimum of 30 measurements was taken per examined specimen. Length of the conidia was measured from the base of the basal end to the base of the apical appendage, and conidial width was measured at the widest point of the conidium. Each length and width values of conidia are reported as one standard deviation (SD) below the mean to one SD above, with the extreme measurements in parentheses followed by the mean ± SD. For other measurement values the full range is reported, from lowest to highest extreme.
PCR and sequencing
DNA was extracted from fungal mycelia grown on MEA with the Promega Genomic DNA purification kit (Fitchburg, Wisconsin, USA) according to the manufacturers’ protocol. PCRs were facilitated in 12.5 μL solutions as in Crous et al. (2013). The ITS rDNA region was amplified with the primers ITS5 (5’-GGAAGTAAAAGTCGTAACAAGG-3’; White et al. 1990) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3’; White et al. 1990), TEF gene with EF728 and EF2 (5’-CATYGAGAAGTTCGAGAAGG-3’ and 5’-GGARGTACCAGTSATCATGTT-3’; O’Donnell et al. 1998), region 1 of TUB with T1 and bt2b (5’-AACATGCGTGAGATTGTAAGT-3’ and 5’-ACCCTCAGTGTAGTGACCCTTGGC-3’; O’Donnell & Cigelnik 1997, Glass & Donaldson 1995) and the RPB2 subunit with 5F2 and 7cR (5’-GGGGWGAYCAGAAGAAGGC-3’ and 5’-CCCATRGCTTGYTTRCCCAT-3’; Liu et al. 1999, Sung et al. 2007). Except for the annealing temperature, the PCR programs for the different loci were identical: 7 min and 30 s of initial denaturation at 95 °C, 35 cycles of 30 s denaturation at 95 °C, 30 s annealing and 90 s elongation at 72 °C and a final elongation step of 7 min 30 s at 72 °C. For ITS the annealing step was at 55 °C and for TEF, TUB and RPB2 annealing temperatures were 53 °C. The amplicons were sequenced with both forward and reverse primers, using an ABI Prism 3730XL Sequencer (Applied Biosystems). Sequences were quality checked and assembled using DNASTAR Lasergene SeqMan Pro v. 8.1.3 software, and deposited in GenBank (Table 1).
Phylogenetic analysis
Sequences acquired in this study were supplemented with those retrieved from GenBank (Barnes et al. 2001, Cunnington 2007, Tsopelas et al. 2007, Maharachchikumbura et al. 2015) and aligned using MAFFT v. 7 (Katoh et al. 2002, Katoh & Standley 2013). Alignments were checked and concatenated in MEGA v. 6.06 (Tamura et al. 2013). Maximum-likelihood (ML) analyses for both single-locus and concatenated alignments were performed with RAxML-HPC2 on XSEDE v. 8.2.10 (Stamatakis 2014) using a GTR+GAMMA substitution model with 1 000 bootstrap iterations. For the Bayesian inference (BI) analyses, the optimal substitution model for ITS, TEF and TUB was determined to be HKY+I+G and for RPB2, GTR+I+G, using MrModeltest software v. 2.2. (Nylander 2004). The BI analyses were computed with MrBayes v. 3.2.6 (Ronquist et al. 2012) with four simultaneous Markov Chain Monte Carlo chains from random trees over 5 M generations, ending the run automatically when standard deviation of split frequencies dropped below 0.01. Both RAxML and Bayesian analyses were run on the CIPRES Science Gateway portal (Miller et al. 2012). Maximum Parsimony (MP) analyses were conducted with PAUP v. 4.0b10 (Swofford 2002), inferring trees with the heuristic search option with TBR branch swapping and 1 000 random sequence additions. The robustness of equally parsimonious trees was evaluated by 1 000 bootstrap replications. Alignments and trees were deposited in TreeBASE (www.treebase.org; study 21661).
RESULTS
Phylogenetic analysis
The concatenated four-locus alignment contained 2 863 nucleotide positions including gaps (ITS: 616, RPB2: 803, TEF: 634 and TUB: 810), comprising 70 strains, including the three outgroup taxa. The phylogram of the best ML tree (lnL = -15056.777813) of the concatenated alignment is shown in Fig. 1. The BI analysis ran 1 345 000 generations before the average standard deviation for split frequencies reached below 0.01. After discarding the first 25 % of generations, 1 010 trees remained from which 50 % consensus trees and posterior probabilities were calculated. In the MP analysis 1 708 characters were identified to be constant, 437 variable characters as parsimony-uninformative and 718 (25.1 %) characters as parsimony-informative. After the heuristic search, 1 000 equally most parsimonious trees were saved (tree length = 408 steps, CI = 0.707, RI = 0.750, RC = 0.530, HI = 0.293). The topologies of the BI and MP trees were similar to the ML tree, which was used to visualize the combined topology (Fig. 1). A node separating the genus Seiridium from the three family members used as outgroup taxa received strong support (99/1/100). Single gene ML, BI and MP trees (Fig. 2) were computed from the same alignments to investigate the suitability of DNA markers used in this study to delineate species.
Fig. 1.
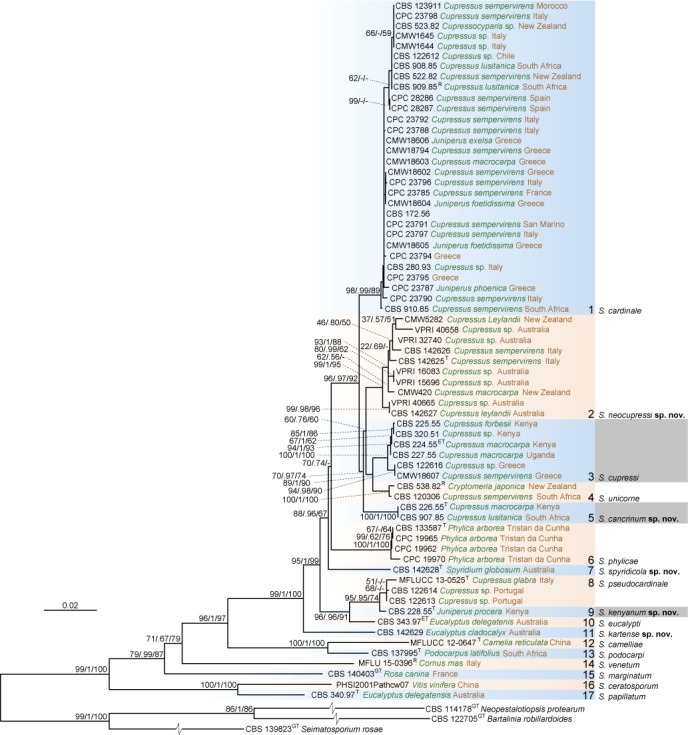
The best Maximum Likelihood tree (lnL = -15056.777813) from the multi-gene alignment (with the 4 loci ITS, RPB2, TUB and TEF) for the Seiridium acquired and sequenced in this study. Nodes are labelled with bootstrap values from RAxML/Bayesian posterior probabilities/Parsimony bootstrap values. Nodes receiving below 50 bootstrap values and 0.5 probability values are not labelled. Grey highlighted names indicate groups containing specimens that were part of the original description of S. cupressi (Guba 1961). Clades 2, 5, 7, 9 and 11 represent new lineages described here as the novel species Seiridium neocupressi, S. cancrinum, S. spyridicola, S. kenyanum and S. kartense. Ex-type culture, ex-epitype culture, ex-generic type culture and reference strains are denoted behind strain numbers with T, ET, GT and R, respectively.
Fig. 2.

The best Maximum Likelihood trees from single-locus alignments. Nodes are labelled with bootstrap values from RAxML/Bayesian posterior probabilities/Parsimony bootstrap values. Nodes receiving below 50 bootstrap values and 0.5 probability values are not labelled. Clade numbers and colours, indicating monophyletic lineages, correspond to the combined phylogeny in Fig. 1: a. The best Maximum Likelihood tree for the ITS alignment including 616 positions (lnL = -2040.052011); b. the best Maximum Likelihood tree for the RPB2 alignment including 803 positions (lnL = -3938.132103); c. the best Maximum Likelihood tree for the TEF alignment including 634 positions (lnL = -3885.959255); d. the best Maximum Likelihood tree for the TUB alignment including 810 positions (lnL = -4789.464706).
The largest clade (clade 1) comprised S. cardinale isolates that appeared to be genetically closely related and monophyletic. Isolates previously labelled as S. unicorne and S. cupressi were, however, polyphyletic. Previously labelled S. unicorne strains clustered in two different clades while labelled S. cupressi strains, including the cultures from the authentic materials (CBS 224.55, CBS 225.55, CBS 226.55, CBS 227.55 and CBS 228.55; Jones 1953, 1954a, Guba 1961) were scattered over five distinctive clades. Clade 2 comprised three yet unclassified specimens (CPC 23786 = CBS 142625, CPC 23789 = CBS 142626 and CPC 28351 = CBS 142627) and specimens identified in previous phylogenetic studies to be S. cupressi (CMW 5282, CMW 420 from Barnes et al. 2001; VPRI 40658, VPRI 32740, VPRI 15696, VPRI 16083 and VPRI 40665 from Cunnington 2007). The cultures from Jones (1953, 1954a, b), on which Guba (1961) based the description of S. cupressi, did not appear in this clade but were present in clades 3 (CBS 224.55 = IMI 52254, CBS 225.55 = IMI 52255 and CBS 227.55 = IMI 52258), 5 (CBS 226.55 = IMI 52256) and 9 (CBS 228.55 = IMI 52257).
Jones (1953, 1954a, b) distinguished different strains based on the aggressiveness of the canker pathogens. While Guba (1961) concluded these were different specimens of one species distinct from S. unicorne, the classification used by Jones (1953, 1954a, b) corresponds to the combined phylogeny in the present study, where strain B (IMI 52254 = CBS 224.55, IMI 52255 = CBS 225.55 and IMI 52258 = CBS 227.55) falls within clade 3, strain A (IMI 52256 = CBS 226.55) in clade 5 and strain D (IMI 52257 = CBS 228.55) in clade 9. Clade 3 also includes the specimens CBS 320.51, CBS 122616 (= CMW 1646 classified as S. cupressi in Barnes et al. 2001) and CMW 18607, classified as S. cupressi in Tsopelas et al. (2007). The fifth clade contains CBS 907.85 (not classified in previous studies) in addition to the strain A specimen from Jones (1953; CBS 226.55). Seiridium cupressi strain D (Jones 1953, Guba 1961; CBS 228.55) clustered together with CBS 122613, CBS 122614 (both classified as S. unicorne in Barnes et al. 2001) and the type of the recently described S. pseudocardinale (Wijayawardene et al. 2016). The fourth clade includes CBS 538.82 (= CMW 5443), which has also been classified as S. cupressi (Barnes et al. 2001).
Other clades included S. phylicae (clade 6; Crous et al. 2012); CBS 142628, isolated from Spyridium globosum in Australia (clade 7), a Seiridium eucalypti strain (CBS 343.97; clade 10) and CBS 142629, isolated from Eucalyptus cladocalyx in Australia (clade 11) and S. marginatum, the generic type (clade 15).
Taxonomy
Based on the results of the combined multi-gene phylogenies (Fig. 1), morphological observations, measurements of fungarium specimens, cultures (Fig. 3) and ecological data, five novel species of Seiridium are described, a lectotype is designated for S. cupressi, and epitypes are selected for S. cupressi and S. eucalypti. Overall, from four clades (3, 5, 7 and 8) all available cultures were sterile. However, DNA was extracted and sequenced from mycelia of these cultures and for three of the clades (3, 5 and 7) morphological characters were studied from the associated herbarium specimens.
Fig. 3.
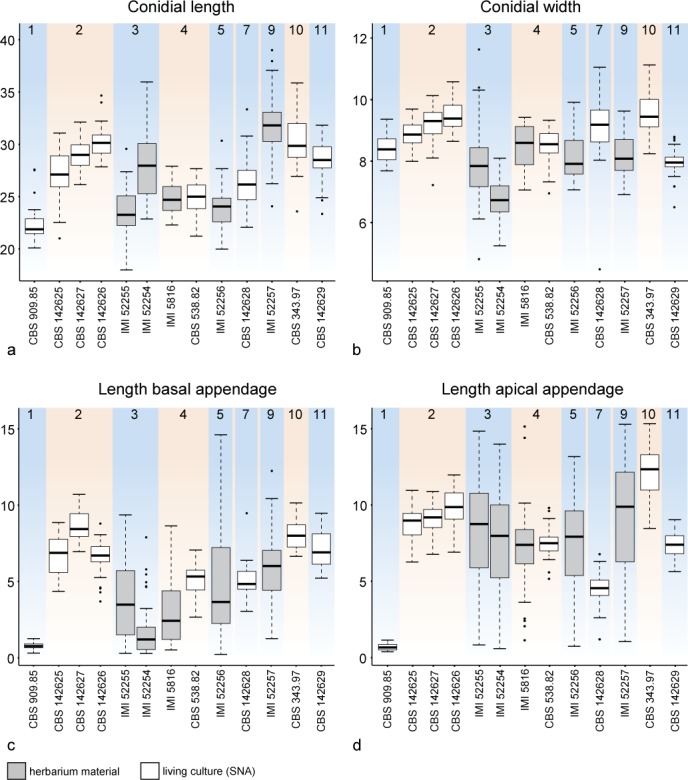
Boxplots reflecting the distribution of variance in conidial measurements in micrometres as acquired from living culture material on SNA (in white) or from herbarium material (in grey). The boxes show the first and third quartiles. Lower and upper whiskers extend from the boxes to the extreme values or 1.5 times the inter quartile range when the extreme values are outside this range, in which case outlying values are indicated by black dots. Strains are ordered by clade as in Fig. 1 and in the background coloured accordingly to the clade colours in Fig. 1: a. Conidial length; b. conidial width; c. length of basal appendage; d. length of apical appendage.
Seiridium Nees, Syst. Pilze (Würzburg): 22. 1817
Synonyms. Hyaloceras Durieu & Mont., Expl. Sci. Algerie 1: 587. 1849.
Adea Petr., Bot. Jahrb. Syst. 62: 144. 1928.
Type species. Seiridium marginatum Nees.
Ascomata perithecial, immersed to semi-erumpent, depressed, globose to pyriform, scattered or confluent; peridium dark brown, pseudoparenchymatous. Ostioles central, slightly papillate, black, periphysate. Paraphyses hyaline, smooth, filiform. Asci cylindrical, 8-spored, unitunicate, thin-walled, stipitate, with an apical amyloid ring. Ascospores cylindrical-oblong, euseptate, septa often thicker than the wall, yellow- to dark brown, guttulate. Conidiomata acervuloid to pycnidioid, semi-immersed to erumpent, uni- to plurilocular, brown or black, glabrous, dehiscing by irregular splits in the upper wall. Conidiophores lining the cavity of the conidioma, septate and sparsely branched at the base, or reduced to conidiogenous cells, hyaline, smooth. Conidiogenous cells discrete, integrated, ampulliform to lageniform or subcylindrical, hyaline, smooth, proliferating percurrently at the apex. Conidia fusiform, distoseptate (septal pores present or not), end cells hyaline, median cells dark brown to brown, wall thick, smooth or with striations, constricted at septa or not; apical cell with a single, cellular, unbranched or branched, appendage; basal cell with or without a centric, unbranched or sometimes branched appendage.
Notes — Seiridium includes coelomycetes producing versicolorous, 5-septate conidia with appendages and typically forming acervuli on the plant host. The original description of the generic type, S. marginatum, dates back over 200 years ago and was re-described by Shoemaker et al. (1966), Sutton (1980) and Nag Raj (1993). Recently, the generic type S. marginatum was epitypified (Jaklitsch et al. 2016) from which additional sequence data was generated in this study and included in the present phylogeny (Fig. 1).
Seiridium cancrinum Bonthond, Sandoval-Denis & Crous, sp. nov. — MycoBank MB823296; Fig. 4
Fig. 4.
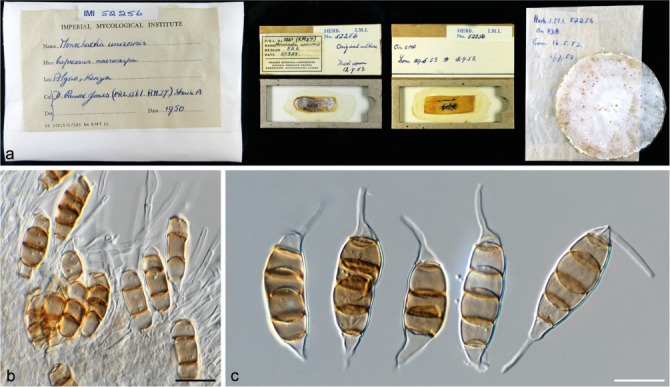
Seiridium cancrinum sp. nov. (IMI 52256, holotype). a. Herbarium specimen; b. conidiogenous cells and conidia; c. conidia. — Scale bars: b–c = 10 μm.
Etymology. From the three strains that Jones (1953, 1954a, b) conducted pathogenic experiments on, this fungus (strain A) was the most aggressive. Therefore, the fungus is named after the canker formation it induces on Cupressus.
Conidia lunate to falcate, curved, 5-septate, occasionally 4-septate, not striate, bearing two appendages, euseptate with no visible pores, (20–)22–26(–30.5) × (7–)7.5–9(–10) μm, mean ± SD = 24.0 ± 2.1 × 8.2 ± 0.7 μm (n = 35); basal cell obconic with truncate base, hyaline, walls smooth, bearing marginal frills, 1.2–5.8 μm; four median cells, varying in colour ranging from pale to dark brown, smooth, cylindrical to doliiform (second cell from base 3–5.5 μm long; third cell 3–5 μm long; fourth cell 3.5–5.5 μm long; fifth cell 3.5–6.5 μm long); apical cell conical, hyaline, smooth, 2–4.5 μm long; apical appendage single, centric, 1–13 μm; basal appendage, single, cylindrical, centric, 0.5–14.5 μm.
Known distribution — Kenya, South Africa.
Materials examined. Kenya, from cankers in branches of Cupressus macrocarpa, 1950, D.R. Jones (holotype IMI 52256, isotype BRL 1119, culture ex-type CBS 226.55). – South Africa, from Cupressus lusitanica, unknown collection date, M.J. Wingfield (CBS 907.85 = CMW 320).
Notes — The pathogenicity of the strain IMI 52256 was studied by Jones (1953, 1954a, b), who classified the fungus as Monochaetia unicornis strain A. Jones identified strain A as the most aggressive of the three strains that would eventually be described as S. cupressi (Guba 1961). However, here we show that the three strains (A: IMI 52256 = CBS 226.55, B: IMI 52254 = CBS 224.55, IMI 52255 = CBS 225.55 and IMI 52258 = CBS 227.55 and D: IMI 52257 = CBS 228.55) are three different species, which in turn correlates with the observations originally made by Jones (1953, 1954a, b). Although both cultures (CBS 226.55 = IMI 52256 and CBS 907.85) remained sterile on culture media, conidia could be studied from herbarium material (IMI 52256). Based on conidial morphology, S. cancrinum is highly similar to S. cupressi, but on average has longer basal appendages. Conidia with basal appendages up to almost 15 μm can be observed, whereas conidia of S. cupressi bear basal appendages that do not exceed 10 μm in length.
Seiridium cardinale (W.W. Wagener) B. Sutton & I.A.S. Gibson, CMI Descriptions of Pathogenic Fungi and Bacteria: 36. 1972 — Fig. 5, 6
Fig. 5.

Seiridium cardinale (CBS 909.85, reference strain). a–d. Colony morphology in 90 mm Petri dishes after 2 wk at 22 °C on PDA, CMA, MEA, SNA, respectively; e. conidioma on PDA partially immersed in agar; f. sporodochia on SNA immersed in agar; g. conidia; h–i. conidiophores; j–k. conidiogenous cells. — Scale bars: e–f = 100 μm; g–k = 10 μm.
Fig. 6.

Seiridium cardinale (IMI 75045 isotype of Coryneum cardinale). a. Herbarium specimen; b–c. conidiomata and conidia in vivo; d. conidia in vitro. — Scale bars: b–c = 20 μm; d = 10 μm.
Basionym. Coryneum cardinale W.W. Wagener, J. Agric. Res. 58: 8. 1939.
Conidiomata on PDA sporodochial, pseudostromatic, globose or clavate, at the edge of the colony, dark brown to black; on SNA sporodochial, pseudostromatic, globose, scattered, irregular in outline, mostly immersed in agar. Conidiophores septate, cylindrical, irregularly branched, branch lengths variable (20–85 μm long), hyaline, thin- and smooth-walled, invested in mucus. Conidiogenous cells discrete, hyaline, subcylindrical to lageniform, thin- and smooth-walled, 4–15 × 1.5–2.5 μm, proliferating percurrently multiple times, with collarettes and minute periclinal thickenings. Conidia fusiform, straight to slightly curved, 5-septate, not striate, bearing two short appendages, euseptate with pores clearly visible, (20–)20.5–24(–27.5) × (7.5–)8–9(–9.5) μm, mean ± SD = 22.3 ± 1.8 × 8.4 ± 0.4 μm (n = 30); basal cell obconic with a truncate base, hyaline, walls smooth, bearing minute marginal frills, 2–3 μm; four median cells, smooth, cylindrical to doliiform, young conidia concave cylindrical to subcylindrical, pale brown to brown, septa darker than the rest of the cells (second cell from base 3.9–4.8 μm long; third cell 3.8–4.8 μm long; fourth cell 4–4.9 μm long; fifth cell 3.9–4.8 μm long); apical cell conical, hyaline, smooth, 2–3 μm long; apical appendage single, centric, short, < 1 μm; basal appendage, single when present, cylindrical, centric, < 1 μm.
Culture characteristics — Colonies on PDA circular, reaching 60–61 mm diam after 14 d at 22 °C, flat at centre and margin, white- to pale luteous-coloured, with aerial mycelium abundant on the surface, sporulating poorly at the margin of the colony and not within 2 wk with pycnidioid conidiomata. On CMA circular, reaching 79–81 mm diam after 14 d at 22 °C, flat at the centre and margin, white-, buff- to hazel-coloured in the centre, hazel-coloured at the inner margin to white at the outer margin, with aerial mycelium formed abundantly on the surface, sporulating poorly, after at least 4 wk with black spore masses produced in sporodochia. On MEA circular, reaching 45–49 mm diam after 14 d at 22 °C, hazel-coloured at the centre surrounded by a grey olivaceous to hazel ring and a white margin, flat at centre and margin, with aerial mycelium abundantly on the surface, sporulating poorly with few sporodochia producing black spore masses. On SNA circular to irregular, reaching 37–39 mm diam after 14 d at 22 °C, flat at centre and margins, with moderate aerial mycelium mostly at the margin, sporulation after approximately 2 wk, sporodochia scattered and immersed in agar.
Known distribution — Africa, Asia, Australia, Europe, New Zealand, North and South America.
Materials examined. New Zealand, Christchurch, 1981, from Cupressocyparis sp., H.J. Boesewinkel (CBS H-18011, culture CBS 523.82 = CPC 23793). – South Africa, East Transvaal, Mac Mac State Forest, 1985, M.J. Wingfield (CBS H-18015, culture CBS 908.85 = CMW 616); Algeria State forest, from Cupressus lusitanica, 1985, M.J. Wingfield (CBS H-18012 reference strain designated here, culture CBS 909.85 = CMW 635). – USA, California, Atherton, from Cupressus macrocarpa, 16 Mar. 1934, W.W. Wagener (isotype of Coryneum cardinale IMI 75045).
Notes — Seiridium cardinale (clade 1) forms a separate cluster in the combined phylogeny with four sister clades including specimens collected from Cupressaceae: S. neocupressi (clade 2), S. cupressi (clade 3), S. unicorne (clade 4) and S. cancrinum (clade 5). Morphologically, S. cardinale is clearly distinct from members of these sister clades by its reduced basal and apical appendages. Since a culture from the correct locality is currently not available, we designated CBS 909.85 as a reference strain, which matches the characteristics of the isotype (IMI 75045). The isotype consists of two slides. One of the slides includes conidia but lacks conidiophores and conidiogenous cells. The second slide is better preserved and contains sections of the fungus in the host, forming acervuli (Fig. 6). By designating a reference strain, we aim to provide a specimen of S. cardinale that contains both a detailed morphological description and is represented by multiple loci in GenBank to promote consistent use in future studies until an appropriate culture is collected from Cupressus macrocarpa in California that can be designated as epitype to supplement the current materials.
Seiridium cupressi (Guba) Boesew., Trans. Brit. Mycol. Soc. 80: 545. 1983 — Fig. 7, 8
Fig. 7.
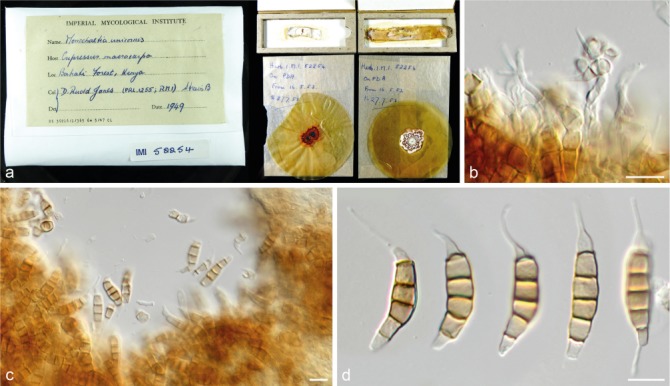
Seiridium cupressi (IMI 52254, lectotype of Cryptostictis cupressi). a. Herbarium specimen; b–c. conidiogenous cells and conidia; d. conidia. — Scale bars: b–d = 10 μm.
Fig. 8.
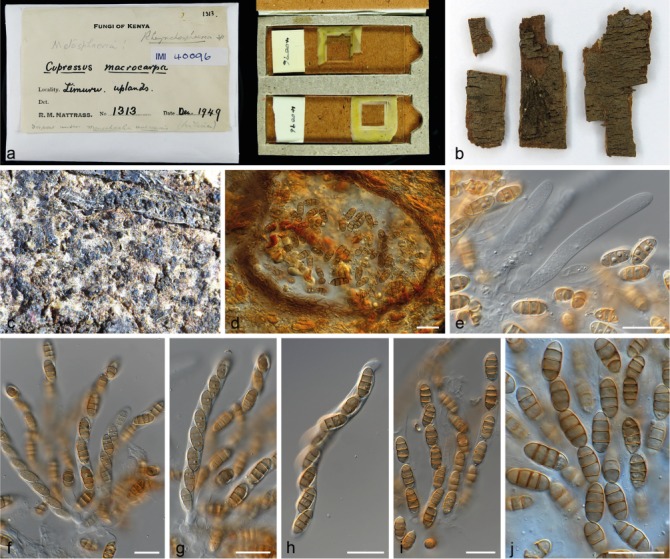
Rhynchosphaeria cupressi (IMI 40096). a–b. Herbarium specimen; c. ascomata; d. ascoma with asci; e–j. paraphyses and asci. — Scale bars: d–j = 20 μm.
Basionym. Cryptostictis cupressi Guba, Monograph of Monochaetia and Pestalotia: 47. 1961.
Synonyms. Rhynchosphaeria cupressi Nattrass et al., Trans. Brit. Mycol. Soc. 46: 103. 1963.
Lepteutypa cupressi (Nattrass et al.) H.J. Swart, Trans. Brit. Mycol. Soc. 61: 79. 1973.
Conidia on dried PDA cultures from Jones (1953) lunate to falcate, curved, 5-septate, not striate, bearing two appendages, euseptate, (18–)22–29.5(–36) × (5–)6–8.5(–11.5) μm, mean ± SD = 25.8 ± 3.6 × 7.4 ± 1.2 μm (n = 61); basal cell obconic with a truncate base, hyaline, 1.5–6 μm; four median cells, smooth, cylindrical to doliiform, pale to dark brown (second cell from base 3–6.5 μm long; third cell 2.5–5.5 μm long; fourth cell 2.5–6 μm long; fifth cell 3–7.5 μm long); apical cell conical, hyaline, 1.5–7 μm long; appendages cylindrical, attenuated; apical appendage single, centric, 0.5–9.5 μm long, occasionally branched near the tip; basal appendage consistently present, single, cylindrical, excentric, 0.5–15 μm long.
Known distribution — Greece, Kenya, Uganda.
Materials examined. Greece, from Cupressus sp., A. Graniti (CBS 122616 = CMW 1646). – Kenya, non pathogenic isolate from Cupressus sp., collection data unknown (CBS 320.51); on Cupressus macrocarpa, July 1948, R.M. Nattrass (IMI 37158 holotype of Rhynchosphaeria cupressi); on Cupressus macrocarpa, Dec. 1949, R.M. Nattrass (IMI 40096); from cankers in branches of Cupressus macrocarpa, 1953, D.R. Jones (IMI 52254 lectotype of Cryptostictis cupressi designated here (MBT379187), BRL 1117 isolectotype, CBS 224.55 epitype designated here (MBT379208), metabolically inactive culture, culture ex-epitype CBS 224.55); from cankers in branches of Cupressus forbesii, 1949, D.R. Jones (IMI 52255 dried culture = BRL 1118, living culture CBS 225.55).
Notes — Clade 3 of the multi-gene phylogeny (Fig. 1) is in the present study identified as S. cupressi based on morphological comparison of the authentic material listed in the protologue (Guba 1961). These materials include one specimen with the sexual morph on Cupressus macrocarpa, collected in Dec. 1949 in the ‘Uplands’, Kenya. Within the IMI fungarium only IMI 40096, Lepteutypa cupressi, carries exactly this label. The other herbarium specimen Guba (1961) cited is from the same host, collector and date, but from Nairobi instead of the Uplands, Kenya and is without mention of a sexual morph. Therefore, we conclude that IMI 40096 is authentic and the specimen from Nairobi was either not preserved, or represents the same strain (Nairobi and the Uplands are only ~ 30 km apart). In addition, Guba (1961) used dried cultures from cankers on Cupressus and Juniperus, collected by Jones in Kenya and Uganda, including the strains A, B, C and D. The IMI fungarium contains five dried cultures that were deposited by Jones (IMI 52254 from Cupressus macrocarpa, IMI 52255 from C. forbesii, IMI 52256 from C. macrocarpa, IMI 52257 from Juniperus procera and IMI 52258 from C. macrocarpa). Together with IMI 40096, these specimens represent the authentic material of S. cupressi. Cultures of the same strains were also deposited in the CBS collection and the dried materials are thus linked to the living cultures CBS 224.55, CBS 225.55, CBS 226.55, CBS 228.55 and CBS 227.55, respectively. Jones (1953, 1954a, b) studied the pathogenic characteristics of the fungus prior to the formal description of S. cupressi by Guba (1961), and classified the fungi in four different strain categories, based on their aggressiveness. Whereas strain A was the most aggressive pathogen to different Cupressus hosts and strain B a mild pathogen, strain D was only pathogenic on Juniperus and appeared to be saprophytic on Cupressus. Strain C comprised mutated clones that had arisen in culture and was never deposited in the IMI or the CBS (see Jones 1954a). To re-evaluate the species and select a lectotype, we examined the herbarium specimen collected by Nattrass (IMI 40096) as well as the dried cultures from Jones (1953, 1954a), with the exception of the specimen from Uganda (IMI 52258 = CBS 227.55). In addition, all loci used in this study were sequenced from the living cultures, which are linked to the authentic dried cultures in the IMI collection. None of the living cultures were fertile, nor were any other cultures examined in this clade (CBS 320.51 and CBS 122616). Despite thorough examination of the material from IMI 40096 only the sexual-morph was observed (Fig. 8), making it impossible to compare this strain to the other authentic material or the original description itself. Therefore, IMI 40096 could not be used for the re-evaluation of S. cupressi. The same applied to the holotype of Rhynchosphaeria cupressi (Nattrass 1963; IMI 37158), in which also no asexual-morph was found, impeding us to link the sexual-morph to the other examined specimens. Furthermore, phylogenetic analysis revealed that the remaining five specimens (the dried cultures from Jones 1953, 1954a) were distributed over three distinct clades (clades 3, 5 and 7; Fig. 1), thus constituting three different species. Morphological comparison (see Fig. 3) showed the specimens in clade 3 (CBS 224.55 = IMI 52254 and CBS 225.55 = IMI 52255) to be similar to the original description. This clade also includes the Ugandan specimen (CBS 227.55 = IMI 52258) and thus the majority of the authentic material. We therefore designate IMI 52254 as lectotype of Cryptostictis cupressi (basionym of S. cupressi), while CBS 224.55, a strain derived from the same collection is designated as epitype in order to provide a stable platform for DNA data comparisons. Because the work of Jones (1953, 1954a, b) preceded the formal description of Seiridium cupressi, each of the dried cultures carries the label ‘Monochaetia unicornis’, including the here designated lectotype IMI 52254. The lectotype material consists of four parts; two dried slant cultures on CMA and two dried plate cultures on PDA. Similar to IMI 52255 the material is in relatively poor condition, but the features of the conidia are clearly recognizable as S. cupressi (Fig. 7). Cultures derived from the other two specimens, CBS 226.55 (= IMI 52256) and CBS 228.55 (= IMI 52257), clustered in clades 5 (S. cancrinum) and 9 (S. kenyanum), respectively. Furthermore, the pathogenic differences that Jones (1953, 1954a, b) identified between the strains (A, B and D) are congruent with the topology of the multi-gene phylogeny (Fig. 1), indicating the three species (S. cancrinum, S. cupressi and S. kenyanum) have different pathology. Morphologically, IMI 52257 deviates most from the other specimens and the original description by bearing considerably larger conidia. This strain was also the only isolate retrieved from Juniperus sp. Instead of Cupressus sp. Specimen IMI 52256 (S. cancrinum; clade 5) is morphologically more similar to the fungarium specimens of S. cupressi (IMI 52254 and IMI 52255; clade 3) and the original description. However, conidia generally have a longer basal appendage (mean ± SD = 5.8 ± 2.4) compared to IMI 52254 and IMI 52255 (mean ± SD = 2.0 ± 2.0 and 3.9 ± 2.6, respectively). Since the living cultures presently available are sterile, future collections from Kenya are required to add more morphological details of this fungus in culture.
Seiridium eucalypti Nag Raj, Coelomycetous anamorphs with appendage-bearing conidia (Ontario): 862. 1993 — Fig. 9
Fig. 9.

Seiridium eucalypti (CBS 343.97, culture ex-epitype). a–d. Colony morphology in 90 mm dishes after 2 wk at 22 °C on PDA, CMA, MEA, SNA, respectively; e–g. sporodochia on SNA; h. conidia; i–j. conidiophores; k. conidiophore; l. conidiogenous cell. — Scale bars: e–g = 100 μm; h–l = 10 μm.
Conidiomata on PDA sterile. On SNA erumpent sporodochia, mostly globose, solitary and compact, producing large black spore masses, sporodochia immersed in agar more aggregated and amorphous. Conidiophores relatively long, varying in length, scattered, septate, cylindrical, irregularly branched, hyaline, smooth- and thin-walled, occasionally reduced to conidiogenous cells, 13–85 μm long. Conidiogenous cells discrete, hyaline, subcylindrical to cylindrical, smooth- and thin-walled, 5–18 × 1.5–3.5 μm, percurrent proliferation rarely observed, with minute periclinal thickenings. Conidia lunate to falcate, straight to curved, 5-septate, not striate, bearing two appendages, euseptate with visible pores, (23.5–)27.5–33(–36) × (8–)9–10.5(–11) μm, mean ± SD = 30.3 ± 2.7 × 9.6 ± 0.7 μm (n = 32); basal cell obconic with a truncate base, hyaline, walls smooth, 4–6 μm; four median cells, smooth, cylindrical to doliiform, brown to dark brown, and septa darker than the rest of the cells (second cell from base 4.5–8 μm long; third cell 4–6.5 μm long; fourth cell 3.5–6 μm long; fifth cell 4.5–7 μm long); apical cell conical, hyaline, smooth, 2.5–6 μm long; apical appendage single, centric, 8.5–15.5 μm long; basal appendage, single, cylindrical, centric and excentric, 6.5–10 μm long.
Culture characteristics — Colonies on PDA irregular, reaching 15–20 mm diam after 14 d at 22 °C, flat at centre, white to pale luteous-coloured at the centre, slightly raised at the margin, white-coloured, with abundant aerial mycelium, not sporulating. On CMA almost circular, reaching 37–42 mm diam after 14 d at 22 °C, flat at centre and margin, white-coloured, with abundant aerial mycelium, sporulating poorly and not within 4 wk, pycnidioid sporodochia producing black spore masses. On MEA irregular, reaching 21–25 mm diam after 14 d at 22 °C, flat at centre and margin, white-coloured, with abundant aerial mycelium, no sporulation. On SNA circular, reaching 33–35 mm diam after 14 d at 22 °C, flat at centre and margin, white-coloured, abundant aerial mycelium, sporulation after 2 wk with scattered compact sporodochia.
Known distribution — Continental Australia, Tasmania.
Materials examined. Australia, South Australia, Adelaide, Mt Lofty Summit, on leaves of Eucalyptus sp., 16 Oct. 1979, B. Kendrick (holotype DAOM 215255); Tasmania, on Eucalyptus delegatensis, 13 Sept. 1996, Z.Q. Yuan (epitype designated here CBS H-23145 (MBT379188), ex-epitype culture CBS 343.97).
Notes — Seiridium eucalypti (clade 10 in Fig. 1) forms a separate clade in the combined phylogeny, sister to S. pseudocardinale and S. kenyanum (clades 8 and 9, respectively). Morphologically, S. eucalypti is characterised by conidia with distinctively long appendages, in particular the apical appendage (Fig. 3). Other Seiridium spp. isolated from Eucalyptus spp. (including S. kartense sp. nov. and S. papillatum) are not monophyletic in the combined phylogeny, nor in any of the single-locus phylogenies (Fig. 2). Seiridium eucalypti is known to inflict lesions on a wide range of Eucalyptus spp. Its pathogenicity to members of Eucalyptus was studied in detail by Yuan & Old (1995) and Yuan & Mohammed (1997, 1999, 2001). The strain examined in this work (CBS 343.97) matches the protologue of S. eucalypti (Nag Raj 1993), and is therefore designated as epitype.
Seiridium kartense Bonthond, Sandoval-Denis & Crous, sp. nov. — MycoBank MB823297; Fig. 10
Fig. 10.
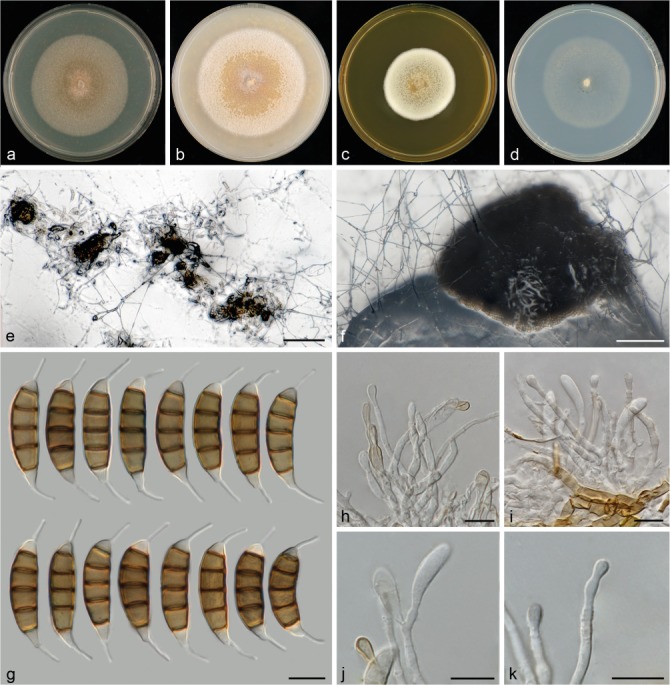
Seiridium kartense sp. nov. (CBS 142629, culture ex-holotype). a–d. Colony morphology in 90 mm dishes after 2 wk at 22 °C on PDA, CMA, MEA, SNA, respectively; e. sporodochia on SNA immersed in agar; f. sporodochia on SNA erumpent from agar; g. conidia; h–i. conidiophores; j. polyblastic conidiogenesis; k. conidiogenous cell proliferating percurrently showing multiple collarettes. — Scale bars: e–f = 100 μm; g–k = 10 μm.
Etymology. Named after the location where the fungus was collected which is known as ‘Kangaroo Island’ and by aboriginals from the mainland of Australia as Karta, meaning ‘Island of the dead’.
Conidiomata on SNA sporodochial, globose to amorphous, solitary, immersed, 160–300 μm diam. Conidiophores short and compact, septate, cylindrical, irregularly branched, hyaline, smooth- and thin-walled, 27–44 μm long. Conidiogenous cells discrete, hyaline, cylindrical to globose, smooth- and thin-walled, 5.4–11.5 × 2.1–3.5 μm, proliferating multiple times percurrently, with minute periclinal thickenings and collarets visible, occasionally polyblastic. Conidia lunate to falcate, curved, 5-septate, not striate, bearing two appendages, euseptate with visible pores, (23.5–)27.5–30.5(–32) × (6.5–)7.5–8.5(–9) μm, mean ± SD = 28.5 ± 1.8 × 8.0 ± 0.4 μm (n = 38); basal cell obconic with a truncate base, hyaline, walls smooth, 4–5.5 μm; four median cells, smooth, cylindrical to doliiform, dark brown (second cell from base 5.5–6.5 μm long; third cell 5–5.5 μm long; fourth cell 4.5–5 μm long; fifth cell 4.5–5.5 μm long); apical cell conical, hyaline, smooth, 3–4 μm long; appendages cylindrical, attenuated, unbranched, slightly spathulate; apical appendage single, centric, 6.5–8.5 μm long; basal appendage consistently present, single, cylindrical, excentric, 6–8.5 μm long.
Culture characteristics — Colonies on PDA circular, reaching 56–59 mm diam after 14 d at 22 °C, flat, from centre to margin; salmon- to umber- to white-coloured, with moderate aerial mycelium, no sporulation. On CMA circular, reaching 65–68 mm diam after 14 d at 22 °C, flat to slightly crateriform, at the centre salmon-coloured, at the margin white-coloured, with abundant aerial mycelium, no sporulation. On MEA circular, reaching 42–44 mm diam after 14 d at 22 °C, raised, ochreous- and honey-coloured at the centre and white-coloured at the margin, with dense aerial mycelium, no sporulation. On SNA circular, reaching 51–54 mm diam after 14 d at 22 °C, flat, white-coloured, with moderate areal mycelium more densely produced at the margin, sporulation after 2 wk with scattered compact erumpent sporodochia in the centre.
Known distribution — Kangaroo Island, Australia.
Material examined. Australia, Kangaroo Island, on leaves of Eucalyptus cladocalyx, 15 Dec. 2012, W. Quaedvlieg (holotype CBS H-23146, ex-type culture CBS 142629 = CPC 20183).
Notes — Seiridium kartense forms a monotypic clade (clade 11 in Fig. 1), and was collected from Kangaroo Island (Australia) on Eucalyptus cladocalyx. Seiridium eucalypti (Nag Raj 1993; clade 10 in Fig. 1) and S. papillatum (Yuan & Mohammed 1997; clade 17 in Fig. 1) have both also been collected from Australia on Eucalyptus spp. Morphologically, S. kartense differs from S. eucalypti based on its conidial length and length of its apical appendage (Fig. 2), which are in both cases shorter than those of S. eucalypti. Seiridium papillatum (see Yuan & Mohammed 1997) is different from S. kartense by its shorter appendages.
Seiridium kenyanum Bonthond, Sandoval-Denis & Crous, sp. nov. — MycoBank MB823301; Fig. 11
Fig. 11.

Seiridium kenyanum (IMI 52257, holotype). a. Herbarium specimen; b–c. conidia. — Scale bars: b–c = 10 μm.
Etymology. Named after the country where it was isolated, Kenya.
Conidia lunate to falcate, curved, 5-septate, not striate, bearing two appendages, euseptate with no visible pores, (20–)22–26(–30.5) × (7–)7.5–9(–10) μm, mean ± SD = 24.0 ± 2.1 × 8.2 ± 0.7 μm (n = 33); basal cell obconic with truncate base, hyaline, walls smooth, bearing marginal frills, 1–6 μm; four median cells, varying in colour ranging from pale to dark brown, smooth, cylindrical to doliiform (second cell from base 3–5.5 μm long; third cell 3–5 μm long; fourth cell 3.5–5.5 μm long; fifth cell 3.5–6.5 μm long); apical cell conical, hyaline, smooth, 2–4.5 μm long; apical appendage single, centric, 0.5–13 μm; basal appendage, single, cylindrical, centric and excentric, 0.5–14.5 μm.
Materials examined. Kenya, from cankers in branches of Juniperus procera, 1951, D.R. Jones (holotype IMI 52257, culture ex-type CBS 228.55); BRL 1121 isotype.
Notes — Phylogenetic analysis and morphological examination of IMI 52257 (= CBS 228.55) revealed this strain to be different from S. cupressi (see notes on S. cupressi). Jones (1953, 1954a, b) classified this fungus as Monochaetia unicornis strain D and already determined that it was ecologically different from Seiridium cupressi and S. cancrinum (Monochaetia unicornis strain B and M. unicornis strain A, respectively) by being non-pathogenic to Cupressus spp. The source of this fungus was Juniperus procera, on which it was a pathogen and induced cankers. Furthermore, its conidia deviate from S. cupressi and S. cancrinum by being considerably larger.
The fungus appears to be genetically highly similar to the sterile cultures CBS 122613 and CBS 122614, as well as to the type of S. pseudocardinale. However, the conidia bear long appendages, and are morphologically clearly different from the latter taxon, as this fungus is characterised by lacking or having reduced appendages (Wijayawardene et al. 2016). Since only an ITS sequence is available for S. pseudocardinale, it is likely that the short distance in the phylogenetic tree between the two species is a result of the limited ability of the ITS rDNA region to delineate species within Seiridium (Fig. 2a but see also Viljoen et al. 1993, Barnes et al. 2001, Maharachchikumbura et al. 2015).
Seiridium neocupressi Bonthond, Sandoval-Denis & Crous, sp. nov. — MycoBank MB823299; Fig. 12
Fig. 12.
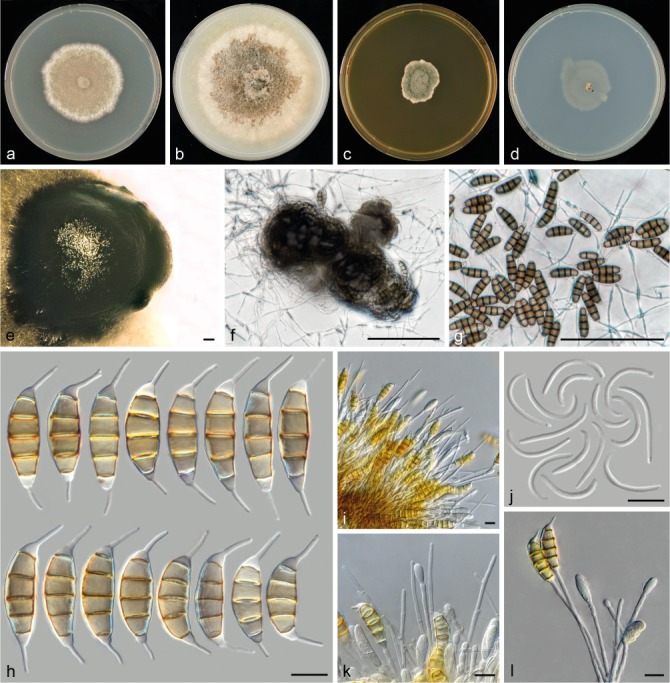
Seiridium neocupressi sp. nov. (CBS 142625, culture ex-holotype). a–d. Colony morphology in 90 mm dishes after 2 wk at 22 °C on PDA, CMA, MEA, SNA, respectively; e. conidioma on PDA; f–g. sporodochia on SNA immersed in agar; h. conidia; i. conidiophores and paraphyses-like structures; j. microconidia (spermatia) on PDA; k–l. conidiophores. — Scale bars: e–g = 100 μm; h–l = 10 μm.
Etymology. neo- cupressi denotes the fact that this phylogenetic clade has been repeatedly incorrectly assumed to represent S. cupressi.
Conidiomata on PDA pycnidioid sporodochial, globose to amorphous, in some strains exuding hyaline mucus, solitary, immersed. On SNA sporodochial, amorphous, immersed in agar. Conidiophores compact, septate, cylindrical, irregularly branched, hyaline, thin-walled, smooth, 13–160 μm long, in some strains conidiophores are intermingled with unbranched, hyaline, paraphyses-like hyphae, up to 115 μm long. Conidiogenous cells discrete, hyaline, cylindrical to globose, sometimes elongated, smooth- and thin-walled, (4–)3.5–24(–65) × (1.5–)2–3(–4) μm, with numerous percurrent proliferations. Conidia lunate to falcate, curved, 5-septate, not striate, bearing two appendages, euseptate without visible pores, (21–)26.5–31(–35.5) × (7–)8.5–10(–10.5) μm, mean ± SD = 28.7 ± 2.3 × 9.2 ± 0.6 μm (n = 94); basal cell obconic with a truncate base, hyaline, 2.5–5 μm; four median cells, smooth, cylindrical to doliiform, pale to brown (second cell from base 2.5–7 μm long; third cell 4–6.5 μm long; fourth cell 4–6.5 μm long; fifth cell 3.5–6.5 μm long); apical cell conical, hyaline, 2.5–5 μm long; appendages cylindrical, attenuated; apical appendage single, centric, 6.5–12 μm long, occasionally branched near the tip; basal appendage consistently present, single, cylindrical, centric and excentric, 3.5–10.5 μm long. Microconidia or spermatia, when produced only on CMA and PDA, cylindrical, hyaline, on PDA (14.5–)18.5–24(–26) μm long.
Culture characteristics — Colonies on PDA circular to irregular, reaching 43–55 mm diam after 14 d at 22 °C, flat at the centre and flat to elevated at the margin, white to pale luteous, sometimes with a white outer ring, with abundant aerial mycelium, sporulating poorly and not within 2 wk with black, scattered pycnidioid conidiomata. On CMA circular, reaching 51–68 mm diam after 14 d at 22 °C, flat at centre and margin to slightly crateriform, buff to saffron with hazel coloured patches in the centre, buff to white coloured at the margin, with moderate aerial mycelium, sporulating after at least 4 wk with black, pycnidioid conidiomata. On MEA irregular, reaching 19–30 mm diam after 14 d at 22 °C, elevated at the centre, flat at the margin, buff to pale green, with aerial mycelium on the surface, no sporulation. On SNA circular to rhizoid, reaching 19–49 mm diam after 14 d at 22 °C, flat, with moderate aerial mycelium, sporulation after a few days with black sporodochial conidiomata.
Known distribution — Australia, Italy.
Materials examined. Australia, Victoria, Torquay, from Cupressus leylandi, 18 Dec. 2006, A. Hoffert (CBS H-23149, culture CBS 142627 = CPC 28351 = VPRI 40665). – Italy, Bari, from Cupressus sempervirens, 23 Nov. 2007, unknown collector (CBS H-23148, culture CBS 142626 = CPC 23789); Bari, from Cupressus sempervirens, 23 Nov. 2007, unknown collector (holotype CBS H-23147, ex-type culture CBS 142625 = CPC 23786).
Notes — The morphology of S. neocupressi bears similarity to the original description of S. cupressi and to the authentic strains that Jones (1953, 1954a) designated as ‘strain B’. Strains CMW 420, CMW 5282, VPRI 15696, VPRI 16083, VPRI 32740, VPRI 40658 and VPRI 40665 were for this reason originally identified as S. cupressi (Barnes et al. 2001, Cunnington 2007, Tsopelas et al. 2007). However, although in the combined phylogenetic analysis the strains A, B and D from which S. cupressi was described clustered in genetically different clades (3, 5 and 9 in Fig. 1), none clustered within clade 2, and thus this clade is distinct from isolates representing S. cupressi. Morphologically, conidia are slightly longer and wider and bear longer basal appendages in comparison to S. cancrinum and S. cupressi.
Seiridium phylicae Crous & M.J. Wingf., Persoonia 29: 187. 2012
Description and illustration — Crous et al. (2012).
Known distribution — UK, Overseas territory of Saint Helena, Ascension, Tristan da Cunha islands.
Material examined. uk, Saint Helena, Ascension and Tristan da Cunha, Inaccessible Island, Blenden Hall, from stems of Phylica arborea, Sept. 2011, P.G. Ryan (holotype CBS H-21089, ex-type cultures CBS 133587 = CPC 19964, CPC 19962, CPC 19965, CPC 19970).
Notes — Seiridium phylicae was described and treated in detail in Crous et al. (2012). Similar to S. spyridicola, this fungus was isolated from a plant host in the Rhamnaceae. Given the sole isolation of this fungus from the Tristan da Cunha archipelago, it seems to be geographically isolated. The species is a problematic pathogen to the Tristan da Cunha endemic Phyilica arborea (Ryan et al. 2014).
Seiridium pseudocardinale Wijayaw. et al., Fung. Diversity 77: 248. 2016
Description and illustration — Wijayawardene et al. (2016).
Known distribution — Italy, Portugal.
Materials examined. Portugal, from Cupressus sp, collection date unknown, A. Graniti (CBS 122613 = CMW 1648); from Cupressus sp., collection date unknown, A. Graniti (CBS 122614 = CMW 1649).
Notes — Seiridium pseudocardinale was recently described by Wijayawardene et al. (2016). In GenBank, only an ITS sequence is available for the type specimen (MFLUCC 13-0525) and thus in the combined phylogeny (Fig. 1) the other loci are missing data for this species. Given the limited resolution on the ITS region alone (Fig. 2a), its phylogenetic placement should thus be interpreted with care. This fungus was isolated from Cupressus glabra in Italy and based on ITS sequence seems to be closely related to cultures CBS 122612 and CBS 122613, which were isolated from Cupressus species in Portugal, and CBS 228.55 (= IMI 52257; S. kenyanum sp. nov.). However, morphologically, the fungus is different from IMI 52257 by the lack of conidial appendages and the poor delineation from CBS 228.55 is likely a result of the absence of informative loci. Since none of the two cultures from Portugal sporulated, we were unable to investigate this clade morphologically and compare it with the description in the protologue. To resolve the phylogenetic relation of S. pseudocardinale to S. kenyanum and the sterile cultures CBS 122613 and CBS 122614, sequence data of at least TUB, but preferably RPB2 and TEF are required from the type (MFLUCC 13-0525).
Seiridium spyridicola Bonthond, Sandoval-Denis & Crous, sp. nov. — MycoBank MB823300; Fig. 13
Fig. 13.
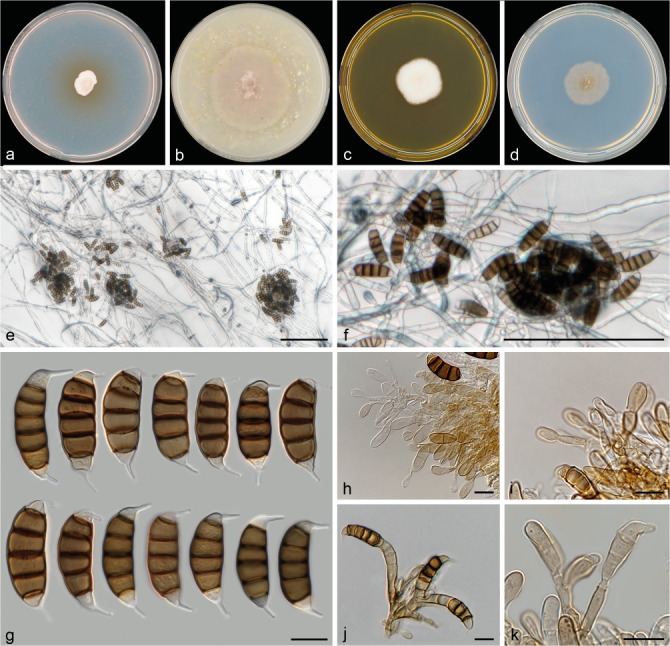
Seiridium spyridicola sp. nov. (CBS 142628, culture ex-holotype). a–d. Colony morphology in 90 mm dishes after 2 wk at 22 °C on PDA, CMA, MEA, SNA, respectively; e–f. sporodochia on SNA immersed in agar; g. conidia; h–j. conidiophores; k. conidiogenous cell. — Scale bars: e–f = 100 μm; g–k= 10 μm.
Etymology. Named after the host genus, Spyridium, from which it was isolated.
Conidiomata on PDA sporodochial, solitary or aggregated, immersed in agar, dark brown to black. Conidiophores septate, cylindrical, irregularly branched, hyaline, smooth- and thin-walled, 9–52 μm long. Conidiogenous cells discrete, hyaline, subcylindrical to globose, smooth- and thin-walled, 5.5–12 × 2–4 μm, proliferating multiple times percurrently, with minute periclinal thickenings. Conidia straight to falcate, straight to slightly curved, 5-septate, sporadically 6-septate, not striate, bearing two appendages, euseptate, pores visible, (22–)24–28.5(–33.5) × (4.5–)8–10.5(–11) μm, mean ± SD = 26.4 ± 2.3 × 9.1 ± 1.2 μm (n = 31); basal cell obconic with a truncate base, hyaline, walls smooth, bearing minute marginal frills, 3–4.5 μm long; four median cells, smooth, doliiform, brown to dark brown, septa darker than the rest of the cells (second cell from base 5–6 μm long; third cell 4.5–5.5 μm long; fourth cell 4.5–5.5 μm long; fifth cell 4.5–5.5 μm long); apical cell conical, hyaline, smooth, 2.5–4 μm long; apical appendage single, excentric and typically oriented perpendicular to conidium, cylindrical, often spatulate, 4–6.5 μm; basal appendage single, cylindrical, often spatulate, mostly centric, 3.5–5.5 μm.
Culture characteristics — Colonies on PDA irregular, reaching 12–16 mm diam after 14 d at 22 °C, flat, from centre to margin, white-coloured, with dense aerial mycelium, sterile. On CMA circular, reaching 46–50 mm diam after 14 d at 22 °C, flat to slightly umbonate, white-coloured, with abundant aerial mycelium on the surface in the centre, mycelium at the margin immersed in agar, sterile. On MEA circular to irregular, reaching 27–29 mm diam after 14 d at 22 °C, flat, white-coloured, with dense aerial mycelium, sterile. On SNA circular, reaching 22–27 mm diam after 14 d at 22 °C, slightly umbonate, cinnamon-coloured at the centre, cinnamon- to white-coloured at the margin, with moderate areal mycelium, sporulation within 2 wk with scattered sporodochia around the centre.
Known distribution — Australia.
Material examined. Australia, Western Australia, from Spyridium globosum, 19 Sept. 2015, P.W. Crous (holotype CBS H-23150, ex-type culture CBS 142628 = CPC 29108).
Notes — This species was isolated from Spyridium globosum (Rhamnaceae) and in the combined phylogeny forms a basal lineage to clades 1 to 6, including most of the Cupressaceae pathogens as well as S. phylicae (Crous et al. 2012), which was isolated from Phylica arborea, a host also belonging to the Rhamnaceae. Conidia are, in comparison to the other studied Seiridium spp. of average length, but the appendages are notably shorter and the apical appendage is typically oriented perpendicular to the long axis of the conidium.
Seiridium unicorne (Cooke & Ellis) B. Sutton, Mycol. Pap. 138: 74. 1975 — Fig. 14, 15
Fig. 14.
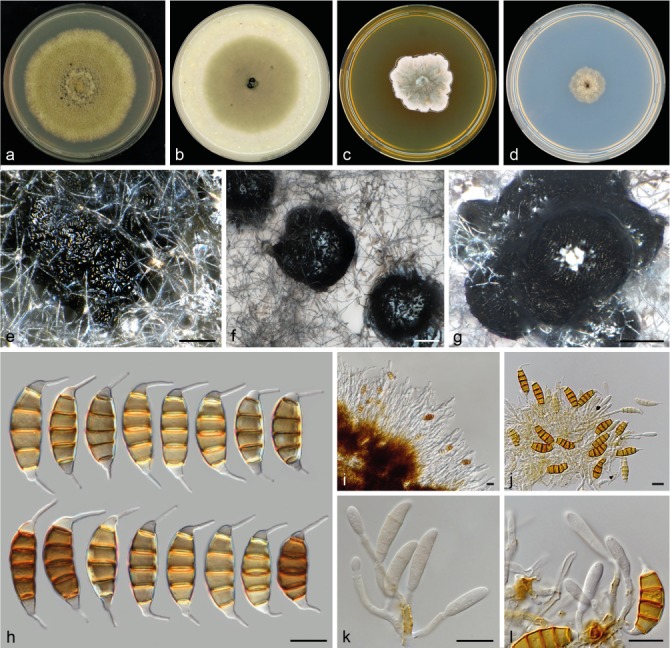
Seiridium unicorne (CBS 538.82, reference strain). a–d. Colony morphology in 90 mm dishes after 2 wk at 22 °C on PDA, CMA, MEA, SNA, respectively; e. conidioma on PDA erumpent from agar partially covered with mycelium; f–g. sporodochia on SNA erumpent from agar; h. conidia; i–j. conidiophores produced from sporodochia; k. conidiophore; l. conidiogenous cells — Scale bars: e–g = 100 μm; h–l = 10 μm.
Fig. 15.
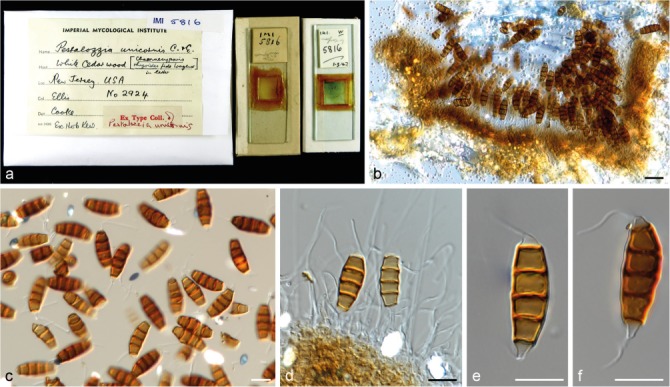
Seiridium unicorne (IMI 5816, holotype of Pestalozzia unicornis). a. Herbarium specimen; b. conidiomata and conidia in vivo; c–f. conidiogenous cells and conidia in vitro. — Scale bars: b = 20 μm; c–f = 10 μm.
Basionym. Pestalotia unicornis Cooke & Ellis, Grevillea 7: 6. 1878.
Synonym. Monochaetia unicornis (Cooke & Ellis) Sacc. & D. Sacc., Syll. Fung. 18: 485. 1906.
Conidiomata on PDA pycnidial to sporodochial, globose or clavate, mostly solitary, erumpent from agar, partially immersed in mycelium, producing large black spore masses. On SNA, sporodochial, mostly aggregated, erumpent from agar, producing large black spore masses. Conidiophores septate, cylindrical, relatively long and irregularly branched, brown or colourless, thin-walled, 29–68 μm long. Conidiogenous cells discrete, hyaline, cylindrical, smooth- and thin-walled, 3.5–29.5 × 1.5–3 μm, proliferating percurrently, with visible collarettes and minute periclinal thickenings. Conidia lunate to falcate, curved, 5-septate, rarely 4- or 6-septate, not striate, bearing two appendages, euseptate with no visible pores, (21.5–)23.5–26.5(–27.5) × (7–)8–9(–9.5) μm, mean ± SD = 24.9 ± 1.6 × 8.5 ± 0.6 μm (n = 31); basal cell obconic with a truncate base, hyaline, walls smooth, bearing minute marginal frills, 3–5.5 μm; four median cells, varying in colour from pale to dark brown, smooth, cylindrical to doliiform (second cell from base 3.5–5.5 μm long; third cell 3.5–5 μm long; fourth cell 4–5.5 μm long; fifth cell 4–5.5 μm long); apical cell conical, hyaline, smooth, 2–5 μm long; apical appendage single, mostly centric, 5–10 μm; basal appendage, single, cylindrical, mostly excentric, 4–6 μm long.
Culture characteristics — Colonies on PDA circular, reaching 65–68 mm diam after 14 d at 22 °C, slightly umbonate, citrine to olivaceous coloured, with aerial mycelium abundant on the surface, sporulating in between centre and margin of the colony within 2 wk. On CMA circular, reaching 58–59 mm diam after 14 d at 22 °C, flat at the centre and margin, citrine to olivaceous coloured, with moderate aerial mycelium on the surface, sporulating in the centre within 2 wk. On MEA irregular, reaching 35–40 mm diam after 14 d at 22 °C, flat to crateriform, slightly sunk into the agar, malachite green coloured at the centre becoming white at the margin with saffron spots or areas in between centre and margin, with dense mycelium on the surface, not sporulating. On SNA circular, reaching 20–21 mm diam after 14 d at 22 °C, umbonate, with aerial mycelium at the surface, sporulation within 2 wk.
Known distribution — New Zealand, South Africa, USA.
Materials examined. New Zealand, from Cryptomeria japonica, 1981, H.J. Boesewinkel (CBS H-23151 reference strain designated here, culture CBS 538.82 = CPC 23783 = IFO 32684). – South Africa, from Cupressus sempervirens, 1999, I. Barnes (culture CBS 120306 = CMW 5596). – USA, New Jersey, associated with Chamaecyparis thyoides, 1878, J.B. Ellis (IMI 5816 holotype of Pestalotia unicornis).
Notes — Culture CBS 120306 remained sterile on all media tested. The holotype material of S. unicorne (IMI 5816) consists of two slides. One of the slides includes conidia, but conidiophores and conidiogenous cells are not discernible, while the second slide contains sections of the fungus in host tissue (Fig. 15). Conidia were produced by CBS 538.82 which were morphologically, apart from slightly longer basal appendages (mean ± SD = 5.35 ± 1.14 opposed to 2.75 ± 2.10), highly similar to the examined holotype material from P. unicornis (see Fig. 3). Despite the highly similar morphological characters, both host and geographic origin of CBS 538.82 (Cryptomeria japonica and South Africa) are different from the holotype which was collected from Cupressus sempervirens in New Jersey. We therefore designate CBS 538.82 as reference strain of S. unicorne, providing a new description of the species and linking DNA sequence data to the morphological description. Pestalozzia unicornis (Cooke & Ellis 1878) was placed in Seiridium by Sutton (1980) and the typically broad host range has traditionally been regarded as an important character of the species. As described by Guba (1961), S. unicorne was found on the Cupressaceae genera Chamaecyparis, Cupressus and Juniperus, but also on Anacardiaceae (Rhus), Caprifoliaceae (Lonicera), Hamamelidaceae (Hamamelis), Rosaceae (Crataegus, Malus, Rosa) and Vitaceae (Vitis). Given that these observations preceded the use of DNA sequence data, it cannot be ruled out that these observations included additional cryptic species.
SPECIES NOT EXAMINED
Seiridium camelliae Maharachch. & K.D. Hyde, Mycol. Progr. 14. 85. 2015
Description and illustration — Maharachchikumbura et al. (2015).
Known distribution — China.
Notes — Seiridium camelliae was isolated from Camellia reticulata (Theaceae) and was described and treated in detail in Maharachchikumbura et al. (2015). In the phylogeny of the concatenated alignment (Fig. 1) as well as the single-locus phylogenies of ITS, TEF and TUB (Fig. 2) S. camelliae resolved as a distinct lineage, closely related to S. podocarpi, which was isolated from Podocarpus latifolus (Podocarpaceae).
Seiridium ceratosporum (De Not.) Nag Raj, Coelomycetous anamorphs with appendage-bearing conidia (Ontario): 859 (1993)
Basionym. Stilbospora ceratospora De Not., Mem. Reale Accad. Sci. Torino Ser. 2, 3: 67. 1841.
Synonyms. Monochaetia ceratospora Guba, Monograph of Monochaetia and Pestalotia: 50. 1961.
Pestalotia ceratospora (De Not.) Arx, The genera of fungi sporulating in pure culture: 226. 1981.
Description and illustration — Nag Raj (1993).
Known distribution — China.
Notes — Seiridium ceratosporum was first described as Stilbospora ceratospora by De Notaris (1841). Nag Raj (1993) examined holotype material and transferred it to Seiridium, providing a re-description of the fungus. Presently, DNA sequence data of the type is not available and the strain included in our analyses (PHSI2001Pathcw07) represents the only strain labelled S. ceratosporum for which multiple sequences are available (including ITS and TUB) and is from Liu et al. (2007) who did not examine morphological characters. Similarly, the strain was included in the ITS phylogeny in Maharachchikumbura et al. (2015) without morphological examination. PHSI2001Pathcw07 was isolated from Vitis vinifera, in contrast to the type which is from Pyrus malus. Therefore, it remains uncertain whether, and perhaps even unlikely, this strain truly is S. ceratosporum. Nonetheless, both single-locus and combined phylogenies indicate it is most closely related to S. papillatum, but separate and could thus be a yet undescribed species in Seiridium as well. Morphological comparison of available or newly collected strains to the protologue (Nag Rag 1993) is required to confirm the identity of this clade as S. ceratosporum.
Seiridium marginatum Nees, Syst. Pilze (Würzburg): 23. 1817
Synonyms. Coryneum marginatum (Nees) Fr., Syst. Mycol. 3: 473. 1832.
Massaria marginata Fuckel, Jahrb. Nassauischen Vereins Naturk. 27–28: 28. 1874.
Blogiascospora marginata (Fuckel) Shoemaker, E. Müll. & Morgan-Jones, Canad. J. Bot. 44: 248. 1966.
Description and illustration — Jaklitsch et al. (2016).
Known distribution — Austria, France, Germany, Switzerland.
Notes — Seiridium marginatum was recently epitypified and treated in detail by Jaklitsch et al. (2016).
Seiridium papillatum Z.Q. Yuan, Austral. Syst. Bot. 10: 70. 1997
Description and illustration — Yuan & Mohammed (1997).
Known distribution — Australia.
Notes — Seiridium papillatum was described and treated in detail by Yuan & Mohammend (1997). Sequence data for LSU, TUB and HIS was generated from ex-type material by Barnes et al. (2001). Additional sequence data was acquired in the present work (ITS, RPB2 and TEF).
Seiridium podocarpi Crous & A.R. Wood, Persoonia 32: 251. 2014
Description and illustration — Crous et al. (2014).
Known distribution — South Africa.
Notes — Seiridium podocarpi was described and treated in detail by Crous et al. (2014).
Seiridium venetum (Sacc.) Nag Raj, Mycotaxon 35: 293. 1989
Basionym. Pestalotia veneta Sacc., Michelia 1: 92. 1877.
Synonyms. Pestalotia corni Allesch., Bot. Zbl. 42: 106. 1890.
Monochaetia veneta (Sacc.) Sacc. & D. Sacc., Syll. Fung. (Abellini) 18: 485. 1906.
Seiridium corni (Allesch.) B. Sutton, Canad. J. Bot. 47: 2091. 1969.
Descriptions and illustrations — Nag Raj (1989), Maharachchikumbura et al. (2015).
Known distribution — Italy.
Notes — Seiridium venetum was re-described and transferred to Seiridium by Nag Raj (1989). Maharachchikumbura et al. (2015) examined the holotype and designated a reference strain (MFLU 15-0396), of which ITS and TUB sequences are included in the combined phylogeny (Fig. 1).
DISCUSSION
The primary character distinguishing Seiridium (Nees 1817) from other pestalotioid genera in the Sporocadaceae (Corda 1842) is its five-septate conidia. For instance the four-septate Pestalotiopsis, and six-septate Truncatella, have in the past been accommodated together with Seiridium in the genus Pestalotia (De Notaris 1841), which underwent many taxonomic rearrangements (Steyaert 1949, Guba 1956, 1961, Sutton 1969, 1980), and now remains a monotypic genus with the type species P. pezizoides from Vitis vinifera. Similar to Seiridium, P. pezizoides produces five-septate conidia and might be a synonym of Seiridium (Maharachchikumbura et al. 2014). Conidial appendages of P. pezizoides are branched, which is also typical for S. corni, S. indicum and S. venetum (Nag Raj 1993), and the host is not unique, as Seiridium spp. have also been isolated from Vitis (Guba 1961). New collections of P. pezizoides are required to select an epitype and generate DNA sequence data to investigate whether Pestalotia is indeed a synonym of Seiridium.
Guba (1961) synonymized Seiridium and Monochaetia and argued to conserve Monochaetia over the older name Seiridium, as the name had been applied more commonly. Shoemaker (1966), however, provided evidence that multiple morphological characters distinguished the two genera from each other. The two names have subsequently been treated as separate genera (Sutton 1980, Nag Raj 1993). Species in Monochaetia share the character of six-celled conidia with Seiridium but typically only bear a single apical appendage. Recent phylogenetic analyses including the species Monochaetia kansensis support the separate identity of the genus (Maharachchikumbura et al. 2014, 2015). However, DNA sequence data linked to the type M. monochaeta has so far not been generated and is essential to provide conclusive evidence on the validity of Monochaetia and its relation to Seiridium and other genera in the Sporocadaceae.
Previous studies have provided phylogenies of Seiridium using ITS, TUB and histone protein H3 (HIS) sequences (Viljoen et al. 1993, Barnes et al. 2001, Cunnington 2007, Tsopelas et al. 2007, Maharachchikumbura et al. 2015, Wijayawardene et al. 2016). As evident from those studies that used solely ITS as phylogenetic marker and in the ITS tree presented in this study (Fig. 2a) this locus provides insufficient information for species delineation within Seiridium. Besides an overall lack of bootstrap support in the phylogeny of this locus alone, S. cardinale does not group under a single node and certain strains (CBS 123911, CBS 523.82 and CPC 23789) cluster with S. eucalypti, S. kenyanum and S. pseudocardinale. In addition to the limited delineation at the species level, separation at the generic level is also problematic, which is reflected in the ITS phylogeny where the outgroup strain Seimatosporium rosae (CBS 139823) clusters within Seiridium. Similarly, in the ITS phylogeny of Maharachchikumbura et al. (2015) Seiridium was not monophyletic. Barnes et al. (2001) showed regions of TUB and HIS to successfully separate Seiridium species. In subsequent studies (Cunnington 2007, Tsopelas et al. 2007) the use of TUB as a species marker was continued and as a result, TUB is at the moment the best available locus in GenBank and has sufficient resolution for delineating species in Seiridium currently known by sequence data. For sequence based species identification of Seiridium isolates this locus is therefore recommended. In addition to ITS and TUB, we sequenced TEF and RPB2, of which the phylogenies (Fig. 2a, b) show these loci are also informative for delineation of Seiridium at the species level. The phylogeny produced from the concatenated alignment, represents the currently most complete phylogeny of Seiridium. However, for many species DNA sequence data have not yet been generated. Our analysis comprises 12 of the 39 names recorded in Index Fungorum and MycoBank for which sequence data are presently available (17 of 44 names, including the novel species).
Apart from re-describing and designating a reference strain for S. cardinale (CBS 909.85 from Cupressus lusitanica), we provide a new description of S. unicorne with reference strain (CBS 538.82 from Cryptomeria japonica). Both species still require an epitype. However, by selecting a reference strain, descriptions of S. cardinale and S. unicorne are linked to DNA sequences in GenBank. From examination of the authentic materials in combination with the multi-gene phylogeny (Fig. 1), we have concluded that the original concept of Cryptosticis cupressi (basionym of S. cupressi) was based on three distinct Seiridium species. One of those species retains the name S. cupressi and the application of this name is here fixed by selection of both a lectotype and epitype. The other two species are introduced as S. cancrinum and S. kenyanum. These three species match the different pathogenic strains (strains A, B and D) Jones (1953, 1954a, b) identified, of which S. cancrinum is the most aggressive, and S. cupressi mildly aggressive. Seiridium kenyanum is only pathogenic to Juniperus (Jones 1953, 1954a, b).
Interestingly, S. kenyanum is more distantly related to the other Cupressaceae pathogens and seems to be related to the recently described S. pseudocardinale (Wijayawardene et al. 2016). This fungus, isolated from Cupressus glabra in Italy, was named after S. cardinale because of the morphological similarity, having reduced to absent conidial appendages. Morphologically, this is somewhat surprising given the large appendages observed for S. kenyanum. However, since only an ITS sequence is available for S. pseudocardinale, the poor delineation of S. kenyanum from S. pseudocardinale is most likely a result of the limited genetic resolution based on the ITS marker (Viljoen et al. 1993). Sequence data of at least TUB, but ideally also TEF and RPB2 of S. pseudocardinale are needed to resolve its phylogenetic position in the genus and relation to S. kenyanum.
In addition to S. cancrinum, S. cupressi and S. kenyanum we introduce S. neocupressi, which represents a lineage previously classified as S. cupressi (Barnes et al. 2001, Cunnington 2007, Tsopelas et al. 2007). Morphologically, conidia and appendages of S. neocupressi are longer compared to the genetically related S. cupressi, S. unicorne and S. cancrinum. Seiridium neocupressi includes canker pathogens primarily from Australia and New Zealand (Barnes et al. 2001, Cunnington 2007, Tsopelas et al. 2007), and most likely represents the most important causal agent of cypress canker in this region.
In summary, the present work provides a re-evaluation of the genus Seiridium. We re-described both S. cardinale and S. unicorne, designating a reference strain for each. Importantly, we examined the authentic material of S. cupressi and sequenced four DNA loci of culture material linked to the holotypes. This resulted in the selection of a lecto- and epitype for S. cupressi and the introduction of two novel species, S. cancrinum and S. kenyanum. Therewith, this work resolves the longstanding confusion surrounding the species S. cupressi. Our analyses included four loci and provide a phylogenetic basis for the genus, including 17 of 44 currently accepted species in Seiridium. We show that in contrast to the ITS region, RPB2, TEF and TUB provide sufficient information for species delineation and recommend future work identifying or introducing species in Seiridium to prioritize the use of TUB over that of the ITS region.
Acknowledgments
We thank Mrs Angela Bond, the Fungarium Collections Manager at Kew for providing the herbarium materials examined in this study.
REFERENCES
- Ballio A, Castiglione Morelli MA, Evidente A, et al. 1991. Seiricardine A, a phytotoxic sesquiterpene from three Seiridium species pathogenic for cypress. Phytochemistry 30: 131–136. [Google Scholar]
- Barnes I, Roux J, Wingfield MJ, et al. 2001. Characterization of Seiridium spp. associated with cypress canker based on ß-tubulin and Histone sequences. Plant Disease 85: 317–321. [DOI] [PubMed] [Google Scholar]
- Boesewinkel H. 1983. New records of the three fungi causing cypress canker in New Zealand, Seiridium cupressi (Guba) comb. nov. and S. cardinale on Cupressocyparis and S. unicorne on Cryptomeria and Cupressus. Transactions of the British Mycological Society 80: 544–547. [Google Scholar]
- Cho W, Shin H. 2004. List of plant diseases in Korea. Korean Society of Plant Pathology, Seoul, Republic of Korea. [Google Scholar]
- Chou C. 1989. Morphological and cultural variation of Seiridium spp. from cankered Cupressaceae hosts in New Zealand. Forest Pathology 19: 435–445. [Google Scholar]
- Cooke MC, Ellis JB. 1878. New Jersey fungi. Grevillea 7: 37–42. [Google Scholar]
- Corda ACJ. 1842. Icones fungorum hucusque cognitorum 5: 1–92. [Google Scholar]
- Crous PW, Braun U, Hunter G, et al. 2013. Phylogenetic lineages in Pseudocercospora. Studies in Mycology 75: 37–114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Shivas RG, Quaedvlieg W, et al. 2014. Fungal Planet description sheets: 214–280. Persoonia 32: 184–306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Shivas RG, Wingfield MJ, et al. 2012. Fungal Planet description sheets: 128–153. Persoonia 29: 146–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crous PW, Verkley GJM, Groenewald JZ, et al. (eds). 2009. Fungal Biodiversity. CBS Laboratory Manual Series 1. CBS-KNAW Fungal Biodiversity Centre, Utrecht, Netherlands. [Google Scholar]
- Cunnington J. 2007. Seiridium cupressi is the common cause of cypress canker in south-eastern Australia. Australasian Plant Disease Notes 2: 53–55. [Google Scholar]
- De Notaris G. 1841. Micromycetes Italici novi vel minus cogniti, decas 2. Memorie della Reale Accademia delle Scienze di Torino Ser. 2, 3: 69–82. [Google Scholar]
- Della Rocca G, Eyre C, Danti R, et al. 2011. Sequence and simple-sequence repeat analyses of the fungal pathogen Seiridium cardinale indicate California is the most likely source of the Cypress canker epidemic for the Mediterranean region. Phytopathology 101: 1408–1417. [DOI] [PubMed] [Google Scholar]
- Della Rocca G, Osmundson T, Danti R, et al. 2013. AFLP analyses of California and Mediterranean populations of Seiridium cardinale provide insights on its origin, biology and spread pathways. Forest Pathology 43: 211–221. [Google Scholar]
- Evidente A, Motta A, Sparapano L. 1993. Seiricardines B and C, phytotoxic sesquiterpenes from three species of Seiridium pathogenic for cypress. Phytochemistry 33: 69–78. [Google Scholar]
- Glass NL, Donaldson GC. 1995. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Applied and Environmental Microbiology 61: 1323–1330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graniti A. 1986. Seiridium cardinale and other cypress cankers. EPPO Bulletin 16: 479–486. [Google Scholar]
- Graniti A. 1993. Seiridium blight of cypress-another ecological disaster? Plant Disease 77: 544. [Google Scholar]
- Graniti A. 1998. Cypress canker: a pandemic in progress. Annual Review of Phytopathology 36: 91–114. [DOI] [PubMed] [Google Scholar]
- Graniti A, Sparapano L, Evidente A. 1992. Cyclopaldic acid, a major phytotoxic metabolite of Seiridium cupressi, the pathogen of a canker disease of cypress. Plant Pathology 41: 563–568. [Google Scholar]
- Guba EF. 1956. Monochaetia and Pestalotia vs. Truncatella, Pestalotiopsis and Pestalotia. Annals of Microbiology 7: 74–76. [Google Scholar]
- Guba EF. 1961. Monograph of Monochaetia and Pestalotia. Harvard University Press, Cambridge, Massachusetts, USA. [Google Scholar]
- Jaklitsch W, Gardiennet A, Voglmayr H. 2016. Resolution of morphology-based taxonomic delusions: Acrocordiella, Basiseptospora, Blogiascospora, Clypeosphaeria, Hymenopleella, Lepteutypa, Pseudapiospora, Requienella, Seiridium and Strickeria. Persoonia 37: 82–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeewon R, Liew EC, Hyde KD. 2002. Phylogenetic relationships of Pestalotiopsis and allied genera inferred from ribosomal DNA sequences and morphological characters. Molecular Phylogenetics and Evolution 25: 378–392. [DOI] [PubMed] [Google Scholar]
- Jones DR. 1953. Studies on a canker disease of Cypresses in East Africa, caused by Monochaetia unicornis (Cooke & Ellis) Sacc. I. Observations on the pathology, spread and possible origins of the disease. Annals of Applied Biology 40: 323–343. [Google Scholar]
- Jones DR. 1954a. Studies on a canker disease of cypresses in East Africa, caused by Monochaetia unicornis (Cooke & Ellis) Sacc. II. Variation in the morphology and physiology of the pathogen. Transactions of the British Mycological Society 37: 286–305. [Google Scholar]
- Jones DR. 1954b. Studies on a canker disease of cypresses in East Africa, caused by Monochaetia unicornis (Cooke & Ellis) Sacc. III. Resistance and susceptibility of species of Cupressus and allied genera. Annals of Applied Biology 41: 325–335. [Google Scholar]
- Katoh K, Misawa K, Kuma K, et al. 2002. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Research 30: 3059–3066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katoh K, Standley DM. 2013. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution 30: 772–780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krokene P, Barnes I, Wingfield BD, et al. 2004. A PCR-RFLP based diagnostic technique to rapidly identify Seiridium species causing cypress canker. Mycologia 96: 1352–1354. [PubMed] [Google Scholar]
- Liu A, Xu T, Guo L. 2007. Molecular and morphological description of Pestalotiopsis hainanensis sp. nov., a new endophyte from a tropical region of China. Fungal Diversity 24: 23–36. [Google Scholar]
- Liu YJ, Whelen S, Hall BD. 1999. Phylogenetic relationships among ascomycetes: evidence from an RNA polymerse II subunit. Molecular Biology and Evolution 16: 1799–1808. [DOI] [PubMed] [Google Scholar]
- Maharachchikumbura SS, Camporesi E, Liu Z, et al. 2015. Seiridium venetum redescribed, and S. camelliae, a new species from Camellia reticulata in China. Mycological Progress 14: 85. [Google Scholar]
- Maharachchikumbura SS, Hyde KD, Groenewald JZ, et al. 2014. Pestalotiopsis revisited. Studies in Mycology 79: 121–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller MA, Pfeiffer W, Schwartz T. 2012. The CIPRES science gateway: enabling high-impact science for phylogenetics researchers with limited resources. In: Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the extreme to the campus and beyond: 1–8. Association for Computing Machinery, USA. [Google Scholar]
- Nag Raj T. 1989. Redisposals and redescriptions in the Monochaetia-Seiridium, Pestalotia-Pestalotiopsis complexes. IX: On the status of Pestalotia eupyrena, Pestalotia gastrolobi and Pestalotia veneta. Mycotaxon 35: 287–296. [Google Scholar]
- Nag Raj T. 1993. Coelomycetous anamorphs with appendage-bearing conidia. Mycologue Publications, Waterloo, Canada. [Google Scholar]
- Nattrass RM, Booth C, Sutton BC. 1963. Rhynchosphaeria cupressi sp. nov., the causal organism of Cupressus canker in Kenya. Transactions of the British Mycological Society 46: 102–106. [Google Scholar]
- Nattrass RM, Ciccarone A. 1947. Monochaetia canker of Cupressus in Kenya. Empire Forestry Review 26: 289–290. [Google Scholar]
- Nees von Esenbeck CGD. 1817. System der Pilze und Schwämme. Würzburg, Germany. [Google Scholar]
- Nylander J. 2004. MrModeltest v2. Program distributed by the author. Evolutionary Biology Centre, Uppsala University. [Google Scholar]
- O’Donnell K, Cigelnik E. 1997. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Molecular Phylogenetics and Evolution 7: 103–116. [DOI] [PubMed] [Google Scholar]
- O’Donnell K, Kistler HC, Cigelnik E, et al. 1998. Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Sciences of the United States of America 95: 2044–2049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rayner RW. 1970. A mycological colour chart. CMI and British Mycological Society, Kew, Surrey, UK. [Google Scholar]
- Ronquist F, Teslenko M, Van der Mark P, et al. 2012. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61: 539–542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryan PG, Ortmann HE, Herian K. 2014. Cascading effects of introduced scale insects on Nesospiza finches at the Tristan da Cunha archipelago. Biological Conservation 176: 48–53. [Google Scholar]
- Saccardo PA, Saccardo D. 1906. Supplementum universale. Pars VII. Discomycetae-Deuteromycatae. Sylloge Fungorum 18: 1–838. [Google Scholar]
- Schoch CL, Seifert KA, Huhndorf S, et al. 2012. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proceedings of the National Academy of Sciences of the United States of America 109: 6241–6246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoemaker R, Müller E, Morgan-Jones G. 1966. Fuckel’s Massaria marginata and Seiridium marginatum Nees ex Steudel. Canadian Journal of Botany 44: 247–254. [Google Scholar]
- Stamatakis A. 2014. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30: 1312–1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steyaert RL. 1949. Contribution à l’étude monographique de Pestalotia de Not. et Monochaetia Sacc. (Truncatella gen. nov. et Pestalotiopsis gen. nov.). Bulletin du Jardin botanique de l’Etat, Bruxelles/Bulletin van den Rijksplantentuin, Brussel 19: 285–354. [Google Scholar]
- Sung G, Sung J, Hywel-Jones NL, et al. 2007. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Molecular Phylogenetics and Evolution 44: 1204–1223. [DOI] [PubMed] [Google Scholar]
- Sutton BC. 1969. Forest microfungi. III. The heterogeneity of Pestalotia de Not. section sexloculatae Klebahn sensu Guba. Canadian Journal of Botany 47: 2083–2094. [Google Scholar]
- Sutton BC. 1975. Coelomycetes. 5. Coryneum. Commonwealth Mycological Institute, Kew. [Google Scholar]
- Sutton BC. 1980. The Coelomycetes. Fungi imperfecti with pycnidia, acervuli and stromata. Commonwealth Mycological Institute, Kew. [Google Scholar]
- Sutton BC, Gibson IAS. 1972. Seiridium cardinale. CMI Descriptions of Pathogenic Fungi and Bacteria 326: 1–2. [Google Scholar]
- Swart HJ. 1973. The fungus causing cypress canker. Transactions of the British Mycological Society 61: 71–82. [Google Scholar]
- Swofford DL. 2002. PAUP*: phylogenetic analysis using parsimony (* and other methods), version 4.0b10. Sinauer Associates Google Scholar, Sunderland, Massachusetts, USA. [Google Scholar]
- Tabata M. 1991. Distribution and host range of Seiridium unicorne in Japan. Transactions of the Mycological Society of Japan 32: 259–264. [Google Scholar]
- Tamura K, Stecher G, Peterson D, et al. 2013. MEGA 6: Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and Evolution 30: 2725–2729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsopelas P, Barnes I, Wingfield MJ, et al. 2007. Seiridium cardinale on Juniperus species in Greece. Forest Pathology 37: 338–347. [Google Scholar]
- Viljoen CD, Wingfield BD, Wingfield MJ. 1993. Comparison of Seiridium isolates associated with cypress canker using sequence data. Experimental Mycology 17: 323–328. [Google Scholar]
- Von Arx JA. 1981. The genera of fungi sporulating in pure culture. Cramer, Vaduz, Liechtenstein. [Google Scholar]
- Wagener WW. 1939. The canker of Cupressus induced by Coryneum cardinale n. sp. Journal of Agricultural Research 58: 1–46. [Google Scholar]
- Wang X, Zhang X, Liu L, et al. 2015. Genomic and transcriptomic analysis of the endophytic fungus Pestalotiopsis fici reveals its lifestyle and high potential for synthesis of natural products. BMC Genomics 16: 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White TJ, Bruns T, Lee S, et al. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innes MA, Gelfand DH, Sninsky et al. (eds), PCR protocols: a guide to methods and applications: 315–322. Academic Press, USA. [Google Scholar]
- Wijayawardene NN, Hyde KD, Wanasinghe DN, et al. 2016. Taxonomy and phylogeny of dematiaceous coelomycetes. Fungal Diversity 77: 1–316. [Google Scholar]
- Xenopoulos S, Diamandis S. 1985. A distribution map for Seiridium cardinale causing the cypress canker disease in Greece. Forest Pathology 15: 223–226. [Google Scholar]
- Xu J, Ebada SS, Proksch P. 2010. Pestalotiopsis a highly creative genus: chemistry and bioactivity of secondary metabolites. Fungal Diversity 44: 15–31. [Google Scholar]
- Yuan ZQ, Mohammed C. 1997. Seiridium papillatum, a new species (mitosporic fungus) described on stems of Eucalypts in Australia. Australian Systematic Botany 10: 69–75. [Google Scholar]
- Yuan ZQ, Mohammed C. 1999. Pathogenicity of fungi associated with stem cankers of eucalypts in Tasmania, Australia. Plant Disease 83: 1063–1069. [DOI] [PubMed] [Google Scholar]
- Yuan ZQ, Mohammed C. 2001. Lesion development in stems of rough- and smooth-barked Eucalyptus nitens following artificial inoculations with canker fungi. Forest Pathology 31: 149–161. [Google Scholar]
- Yuan ZQ, Old K. 1995. Seiridium eucalypti, a potential stem canker pathogen of eucalypts in Australia. Australasian Plant Pathology 24: 173–178. [Google Scholar]