
Figure 2.
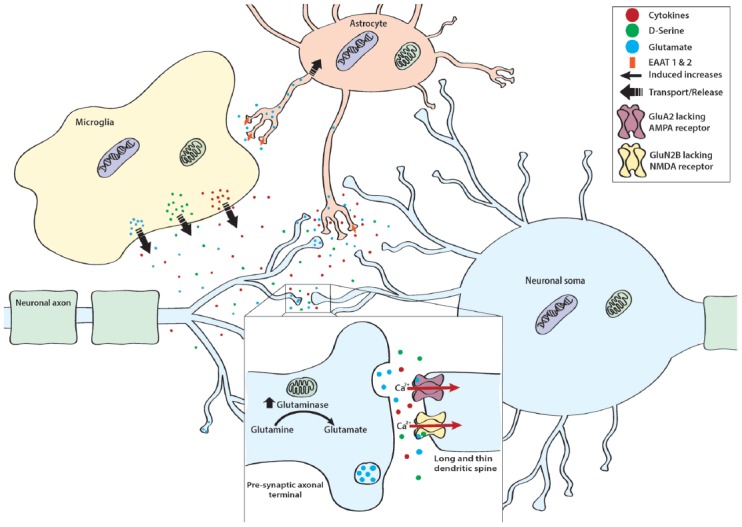
Mechanisms of stress-induced cytotoxicity. In addition to releasing cytokines, Ml type proinflammatory microglia release a variety of neurotransmitters, co-agonists, and neuromodulators such as glutamate and its co-agonist D-serine. Normally, excess glutamate is taken up by excitatory amino acid transporter (EAAT) 1 and 2 found on astrocytic processes. However, in proinflammatory conditions and in the presence of excess glutamate, EAAT 1 and 2 are down regulated, thereby resulting in excess glutamate within the synaptic cleft. Importantly, neurons also contribute to stress-induced enhancements of glutamatergic tone. This is thought to occur as stress exposure enhances mitochondrial glutaminase, the enzyme responsible for converting glutamine to glutamate. In addition to enhancing excitatory tone, stress also sensitizes neurons to the excitatory effect of glutamate. Specifically, stress promotes the expression of GluA2 lacking α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors and GLUN2B lacking NMDA receptors. These receptor subtypes allow for calcium (Ca2+) to freely pass into the cell thereby enhancing the depolarizing effect of glutamate. This cumulative increase in excitatory tone is particularly detrimental for dendritic spines that exhibit a long and thin morphology, as these spines are more sensitive to the degenerative effects induced by glutamatergic excitotoxicity.