

Abstract
The contribution of epigenetic variation to phenotypic variation is unclear. Imprinted genes, because of their strong association with epigenetic modifications, represent an opportunity for the discovery of such phenomena. In mammals and flowering plants, a subset of genes are expressed from only one parental allele in a process called gene imprinting. Imprinting is associated with differential DNA methylation and chromatin modifications between parental alleles. In flowering plants imprinting occurs in a seed tissue - endosperm. Proper endosperm development is essential for the production of viable seeds. We previously showed that in Arabidopsis thaliana intraspecific imprinting variation is correlated with naturally occurring DNA methylation polymorphisms. Here, we investigated the mechanisms and function of allele-specific imprinting of the class IV homeodomain leucine zipper (HD-ZIP) transcription factor HDG3. In imprinted strains, HDG3 is expressed primarily from the methylated paternally inherited allele. We manipulated the methylation state of endogenous HDG3 in a non-imprinted strain and demonstrated that methylation of a proximal transposable element is sufficient to promote HDG3 expression and imprinting. Gain of HDG3 imprinting was associated with earlier endosperm cellularization and changes in seed weight. These results indicate that epigenetic variation alone is sufficient to explain imprinting variation and demonstrate that epialleles can underlie variation in seed development phenotypes.
Author summary
The contribution of genetic variation to phenotypic variation is well-established. By contrast, it is unknown how frequently epigenetic variation causes differences in organismal phenotypes. Epigenetic information is closely associated with but not encoded in the DNA sequence. In practice, it is challenging to disentangle genetic variation from epigenetic variation, as what appears to be epigenetic variation might have an underlying genetic basis. DNA methylation is one form of epigenetic information. HDG3 encodes an endosperm specific transcription factor that exists in two states in A. thaliana natural populations: methylated and expressed and hypomethylated and repressed. We show that pure epigenetic variation is sufficient to explain expression variation of HDG3 - a naturally lowly expressed allele can be switched to a higher expressed state by adding DNA methylation. We also show that expression of HDG3 in strains where it is normally hypomethylated and relatively repressed causes a seed development phenotype. These data indicate that naturally circulating epialleles have consequences for seed phenotypic variation.
Introduction
DNA methylation is a heritable epigenetic mark that can, on occasion, effect gene transcription and influence development. DNA methylation is a particularly influential regulator of gene expression in endosperm, a triploid extraembryonic seed tissue that supports embryo development. In endosperm, developmentally programmed DNA demethylation causes maternally inherited endosperm genomes to be hypomethylated compared to the paternally inherited endosperm genome [1–3]. Methylation differences between maternal and paternal alleles identify their parent-of-origin and establish imprinting, an epigenetic phenomenon in which a gene is expressed primarily from one parental allele [4]. Imprinting is theorized to have evolved over conflict between maternally and paternally inherited alleles in offspring over the extent of maternal investment [5,6]. Under the kinship theory, over evolutionary time silencing of the maternally inherited allele and expression of the paternally inherited allele is predicted to result for genes where the paternally inherited allele’s optimum expression level in offspring is higher than the maternally inherited allele’s [7]. Comparison of imprinting between species in the Arabidopsis genus has provided empirical support for this hypothesis [8,9].
Recent genomic approaches have revealed extensive natural DNA methylation variation within Arabidopsis thaliana [10,11]. Whereas the contribution of genetic variation to phenotypic diversity is well-established, the impact of epigenetic variation, or epialleles, on phenotype is only beginning to be understood [12,13]. Processes affected by epialleles include patterns of floral development, sex determination, fruit ripening, fruit nutritional content, and senescence, among others [14–19]. We previously demonstrated that natural variation in DNA methylation is associated with imprinting variation, with as many as 10% of imprinted genes estimated to be variably imprinted within A. thaliana and maize [20,21]. Seed development varies extensively among Arabidopsis accessions and has previously been shown to be influenced by parent-of-origin effects [20,22], thus raising the possibility that variation in imprinting could influence seed phenotypes. One of these variably imprinted genes, HOMEDOMAIN GLABROUS3 (HDG3), is a member of the class IV homeodomain leucine zipper transcription factor (HD-ZIP) family, which regulates diverse aspects of plant patterning and development [23,24]. Studies on the function of class IV HD-ZIP genes in trichome differentiation, sepal giant cell formation, and suppression of somatic embryogenesis, among others, have led to the conclusion that class IV HD-ZIP family genes promote endoreduplication and cell differentiation [24–27]. Several members of the class IV HD-ZIPs are primarily expressed in endosperm and exhibit imprinted expression patterns, including FWA/HDG6, HDG8, HDG9 and HDG3 [2,23,28]. FWA, HDG8, and HDG9 are maternally expressed imprinted genes (MEGs), whereas HDG3 is a paternally expressed imprinted gene (PEG) [2,28]. The function of the imprinted class IV HD-ZIP genes during seed development, if any, is unknown.
The activity of HDG3 alleles is correlated with DNA methylation. In endosperm of imprinted strains, the highly expressed paternal HDG3 allele is methylated and the lowly expressed maternal allele is hypomethylated over a Helitron TE sequence 5’ of the transcriptional start site [2]. Maternally inherited endosperm alleles are demethylated by the 5-methylcytosine DNA glycosylase gene DME; in dme mutants, maternal alleles retain their methylation and are also expressed [2,29]. Of 927 Arabidopsis accessions with sufficient methylation data [11], 32 (3.5%) have no methylation in the HDG3 5’ region and 871 (94%) have greater than 50% methylation. When strains where HDG3 methylation is low, such as Cvi or Kz_9, are the paternal parent in crosses with Col, where the maternal allele is demethylated, there is no methylation difference between maternal and paternal alleles in endosperm and HDG3 is biallelically expressed [20]. Together, these data suggest that (1) DNA demethylation promotes repression of the maternally-inherited HDG3 allele whereas DNA methylation promotes expression (or inhibits repression) of the paternal HDG3 allele and that (2) imprinting variation is due to cis epigenetic variation at HDG3 [20]. However, a cis or trans genetic contribution to imprinting variation cannot be excluded because of DNA sequence polymorphisms between the strains and alleles that do and do not exhibit imprinting.
Here, we show that a naturally occurring epiallele can contribute to variation in seed phenotypes in Arabidopsis. We tested whether cis epigenetic variation is sufficient to explain imprinting variation by generating a methylated HDG3 Cvi allele that mimicked a methylated HDG3 Col allele. We found that the HDG3 Cvi allele switched from a hypomethylated, non-imprinted, repressed state to an imprinted, paternally biased, expressed state. Additionally, gain of HDG3 imprinting altered endosperm development and final seed size. These data indicate that naturally occurring epialleles can have phenotypic consequences in endosperm, a tissue where methylation is dynamic as a programmed part of development.
Results
Natural variation in HDG3 imprinting is associated with gene expression differences
We previously showed that several genes that are imprinted in endosperm when Col is the paternal parent are not imprinted when Cvi is the paternal parent [20]. To further examine naturally occurring endosperm gene expression variation, we sequenced the transcriptomes of endosperm from Col x Col and Col x Cvi F1 seeds. Comparison of these transcriptomes identified 957 genes that were expressed two-fold or higher in Col x Col and 1187 that were expressed two-fold or higher in Col x Cvi endosperm (Fig 1A; S1 Table). The gene with the lowest expression in Col x Cvi relative to Col is HDG3, which is expressed 64-fold lower in Col x Cvi endosperm (Fig 1A).
Fig 1. Natural variation in imprinting is associated with differences in HDG3 expression levels.

(A) HDG3 expression is decreased in Col x Cvi endosperm compared to Col, as determined by mRNA-seq. (B) Schematic of the HDG3 locus. DMR, differentially methylated region. (C) In situ hybridization of HDG3 (purple) in F1 seeds from the indicated crosses. Female parent is written first. In crosses where Col is the male parent, HDG3 is detected in the micropylar (MCE), peripheral (PEN), and chalazal (CZE) endosperm. Arrowheads indicate nuclear-cytoplasmic domains in the PEN. Number of seeds with shown pattern out of total seeds assayed is in corner of each image. Scale bars, 50 μm. (D) RT-qPCR analysis of relative HDG3 transcript abundance in F1 endosperm. Values are the average of 3 biological replicates, bars represent upper and lower range. (E) Schematic representation of relationship between HDG3 methylation, expression, and imprinting in endosperm. Thickness of arrows denotes relative expression level. Lollipops represent methylated (filled) and unmethylated (open) cytosines.
We previously reported that HDG3 is a PEG in Cvi x Col crosses but is biallelically expressed in Col x Cvi [20]. To further explore the expression variation of HDG3, we performed in situ hybridization on developing seeds (Fig 1B–1C; S1 Fig). In Col x Col seeds, HDG3 is expressed specifically in the micropylar, peripheral, and chalazal endosperm, with the highest expression at the heart stage of development (Fig 1C). The same pattern was observed in Cvi x Col (Fig 1C). Whereas HDG3 expression was detected by in situ hybridization in F1 endosperm when Col was the paternal parent, it was not detected in endosperm when Cvi was the paternal parent (Fig 1C). Additionally, we performed RT-qPCR on biological triplicates of Col, Cvi, and Col-Cvi F1 endosperm. Expression in Col x Col and Cvi x Col was approximately 10-fold higher than in Cvi x Cvi or Col x Cvi, indicating that HDG3 expression is higher when it is imprinted (Fig 1D), consistent with the mRNA-seq (Fig 1A) and in situ data (Fig 1C). Thus, although expression is from both maternally and paternally inherited alleles in Col x Cvi crosses (and presumably Cvi x Cvi crosses) as detected by mRNA-seq [20], the total expression in those crosses is lower than when HDG3 is imprinted. As we previously showed that the Cvi allele is naturally hypomethylated [20], together these results suggest that DNA methylation of the HDG3 5’ region promotes HDG3 expression (Fig 1E).
There is also evidence for imprinting variation of HDG3 in other species. In Arabidopsis lyrata, expression of HDG3 is specific to the endosperm but levels differ between two accessions, MN47 and Kar, and their reciprocal crosses (S2 Fig). In endosperm with high HDG3 expression (Kar x MN47), expression is strongly paternally biased (76% paternal instead of the expected 33%), whereas in the reciprocal cross expression of HDG3 is much lower and more reflective of the 2:1 maternal:paternal ratio in the endosperm (79% maternal) (S2 Fig) (8). The correlation between high expression of HDG3 and paternal allele bias in A. lyrata thus mirrors A. thaliana.
Reduced HDG3 expression affects seed development
To examine if HDG3 influenced endosperm development, we compared seeds from hdg3 mutant plants and segregating wild-type siblings in the Col background. We confirmed predominantly paternal expression of HDG3 [2,20] by reciprocal crosses between wild type and hdg3-1 mutants (Fig 2A). When hdg3 was crossed as a female to a wild-type sibling male, expression of HDG3 was detected in endosperm in a similar manner as in Col x Col (Fig 2A). In contrast, when wild-type females were crossed to hdg3 mutant males, the accumulation of HDG3 transcript in endosperm was dramatically affected, with no transcript detected in most cases, despite the presence of a wild-type maternally inherited allele (Fig 2A). We assessed embryo stage and the extent of endosperm cellularization for sectioned wild-type and hdg3 seeds at 5 days after pollination. Embryo development was more variable in hdg3, although this difference was not statistically significant, and endosperm cellularization was significantly delayed compared to wild-type seeds (Fig 2B, S2 and S3 Table). Reciprocal crosses between wild-type and hdg3 mutant plants indicated that the endosperm cellularization phenotype was dependent on paternal genotype, consistent with HDG3 function being primarily supplied from the paternally-inherited allele (S2 and S3 Table). Additionally, the weight and area of hdg3 seeds was slightly reduced compared to Col, suggesting that in the Col background HDG3 promotes seed growth or filling (Fig 2C and 2D). Several PEGs have been shown to influence seed abortion phenotypes in interploidy crosses [30,31], but we found no effect of hdg3 on this process (S3 Fig). To understand the potential molecular consequences of the loss of hdg3, we profiled endosperm gene expression in wild-type Col and hdg3-1 by RNA-seq at 7 days after pollination (DAP) (Fig 3). 150 genes had at least two-fold higher expression upon loss of hdg3, while 238 genes had at least two-fold lower expression in hdg3 mutant endosperm (Fig 3, S4 Table). Differentially expressed genes included developmental regulators such as HOMEOBOX 3 (WOX9) and gibberellin oxidases, which effect the level of a key phytohormone necessary for typical seed development [32] (Fig 3). The loss of hdg3 also impacted the expression of ten imprinted genes, including the MEG HDG9 (Fig 3). We hypothesized that the endosperm gene expression phenotypes associated with low expression of HDG3 from Cvi paternal alleles might in some respects mimic hdg3 mutants. Indeed, of the 238 genes that are down-regulated in hdg3 mutants, 100 are also down regulated in Col x Cvi crosses, where HDG3 expression is also low (Fig 3). This is a highly significant overlap (hypergeometric test in R, p = 6.079e-69) (Fig 3). These data suggest that the Cvi HDG3 allele, in its hypomethylated and relatively transcriptionally repressed state, could be important for some of the accession-specific developmental traits imparted by Cvi [20,22,33]. Thus, to test both the imprinting mechanism and function of HDG3 further, we introduced methylation at the HDG3 locus in Cvi.
Fig 2. Phenotypic effects of mutation of HDG3 in Col.

(A) In situ hybridization of HDG3 (purple) in seeds from the indicated crosses. Female parent is written first. Scale bars, 50 μm. (B) Endosperm cellularization is slightly delayed in hdg3 compared to wild-type at 5 DAP. For each seed the embryo stage was determined and then the stage of endosperm cellularization was defined as normal, early, or delayed given that embryo stage. (C) Seed weight of wild-type and hdg3 seeds. Individual data points and mean +/- SD shown. P-value from unpaired two-tailed t-test. (D) Seed area is significantly reduced in hdg3 seed (n = 275) compared to wild-type siblings (n = 376) (p = 8.51e-11 by Welch’s two tailed t-test). Seeds were quantified with ImageJ.
Fig 3. Transcriptional effects associated with low HDG3 expression.
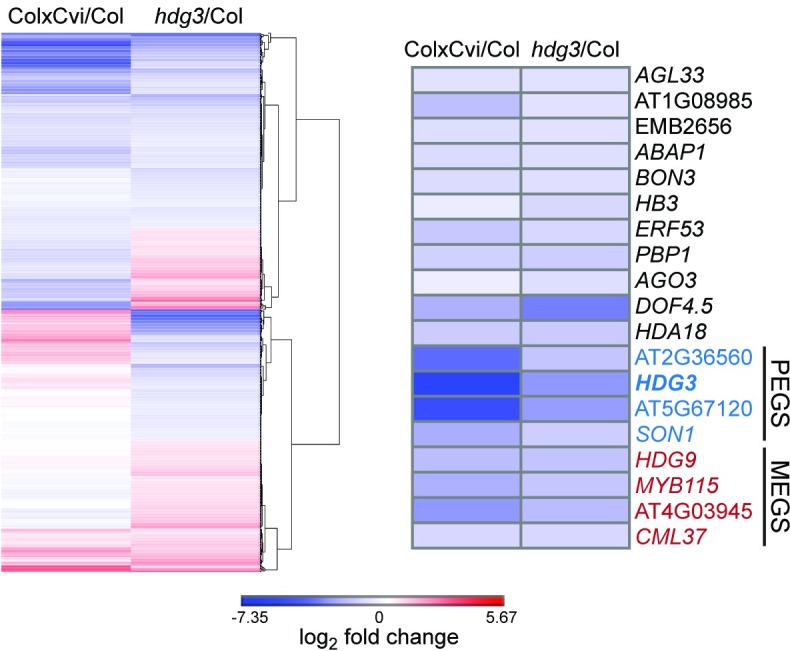
(left panel) Genes downregulated in hdg3 mutant endosperm also have reduced expression in Col x Cvi endosperm compared to Col. The plot shows the expression profile of genes with significantly altered expression in hdg3-1 endosperm (p<0.05). Genes were hierarchically clustered by Euclidean distance and complete linkage using Gene-E. (right panel) A subset of putative developmental regulators with reduced expression in hdg3 and Col x Cvi endosperm.
An inverted repeat induces methylation in the region 5’ of HDG3 in Cvi
To distinguish the importance of genetic variation from epigenetic variation for HDG3 expression and imprinting, we generated transgenic lines in which the endogenous HDG3 Cvi allele gained methylation in the same region that is methylated in Col. Cvi was transformed with a transgene consisting of an inverted repeat (HDG3 IR) of the 450 bp HDG3 5’ region from Cvi under the control of the constitutive 35S promoter. Processing of the expressed hairpin RNA into small RNAs is expected to direct methylation to the endogenous HDG3 Cvi locus. We identified multiple independent transgenic lines in which the HDG3 5’ region gained methylation in leaves (S4 Fig). DNA methylation was present in the same region as in Col, although non-CG methylation was considerably higher (S4 Fig).
To determine whether the Cvi allele remained methylated when paternally inherited in endosperm, Cvi HDG3 IR plants from three independent transgenic lines were crossed as males to wild type Col females and DNA methylation was evaluated in F1 endosperm by locus-specific bisulfite-PCR. Although the 35S promoter has been reported to have no activity in syncytial endosperm [34], we detected transcripts from the hairpin RNA in endosperm at 7 DAP (S5 Fig). Bisulfite sequencing showed that the paternally inherited HDG3 Cvi allele from the IR line was hypermethylated relative to the paternally inherited HDG3 Cvi allele in Col x Cvi endosperm (Fig 4; S6 Fig). The HDG3 Cvi allele from Col x Cvi HDG3 IR endosperm was methylated in both CG and non-CG contexts, indicative of RNA-directed DNA methylation, although at a lower level than in leaves or in F1 embryos (S6 Fig). Examination of the bisulfite clones indicated some variation in paternal allele methylation, with clones with 0% methylation detected, unlike naturally methylated paternal alleles from Cvi x Col crosses (S6 Fig). This could be due to stochastic silencing of the IR transgene in individual siliques/seeds or ineffective RNA-directed DNA methylation. The maternally inherited Col allele was unaffected in Col x Cvi HDG3 IR endosperm, remaining hypomethylated like in Col x Cvi endosperm. Thus, we successfully established an alternate epigenetic state specifically for the Cvi HDG3 allele in endosperm.
Fig 4. Gain of Cvi HDG3 paternal allele methylation in endosperm.

Total methylcytosine 5’ of HDG3 in F1 endosperm, determined by bisulfite-PCR. Maternally inherited Col allele in orange, paternally inherited Cvi allele in blue. Scale to 100%, tick marks below line indicate unmethylated cytosines. Col x Cvi data published in [20].
Methylation of the HDG3 5’ region is sufficient to promote expression and imprinting
Having established a methylated Cvi HDG3 allele, we tested whether paternal allele methylation was sufficient to switch HDG3 from a non-imprinted, repressed state to an imprinted, more active state. In two independent lines, in situ hybridization of F1 seeds from Col x Cvi HDG3 IR crosses indicated the presence of HDG3 transcript in endosperm, in contrast to Col x Cvi endosperm (Fig 5A). Hybridization signal was primarily detected in uncellularized endosperm on the chalazal side of the peripheral endosperm (Fig 5A). However, the penetrance of Cvi HDG3 expression was variable, with about half of the seeds exhibiting HDG3 expression detectable by in situ (Fig 5A). This might be related to the variation in methylation of the HDG3 Cvi allele in Col x Cvi HDG3 IR seeds (S6 Fig). Analysis of total HDG3 transcript abundance by RT-qPCR at 6–7 days after pollination showed that HDG3 expression was 2-3-fold higher in Col x Cvi HDG3 IR endosperm compared to Col x Cvi endosperm (Fig 5B). Higher expression of HDG3 in Col x Cvi HDG3 IR endosperm is consistent with HDG3 being more highly expressed when imprinted (Fig 1). Thus, to measure allele-specific expression of HDG3, Col and Cvi alleles were distinguished using TaqMan probes in an RT-qPCR assay. In crosses between Col females and three independent Cvi HDG3 IR lines, the fraction of transcript derived from the Cvi allele increased compared to control crosses between Col females and Cvi males. In Col x Cvi, the Cvi allele accounts for 23% of the transcripts by this assay, in good agreement with prior allele-specific mRNA-seq results [20]. In Col x Cvi HDG3 IR lines, the Cvi fraction was between 50–60%, indicating paternal allele bias (the expectation for non-imprinted genes is 33% paternal) (Fig 5C). This is slightly less than the fraction of paternal allele expression in Cvi x Col crosses by mRNA-seq (79%) [20]. Together, these data indicate that the naturally occurring methylation variation at HDG3 is sufficient to explain imprinting variation. We conclude that the methylated Cvi HDG3 allele in Cvi HDG3 IR plants is active and the gene is imprinted.
Fig 5. HDG3 is imprinted in Cvi HDG3 IR lines.

(A) HDG3 in situ hybridization for indicated genotypes. Arrowheads indicate regions of in situ signal. Right panels shows magnification of chalazal region. Scale bars, 50 μm. (B) RT-qPCR of relative HDG3 transcript abundance in F1 endosperm at 6–7 DAP. Dashed line separates experiments done at different times. Left, avg of 3 technical replicates. Right, avg of biological duplicates. Bars show upper and lower range. (C) % of HDG3 from Cvi allele in endosperm by TaqMan RT-qPCR assay.
Expression of HDG3 in Cvi promotes endosperm cellularization
Does the change of HDG3 expression and imprinting in Cvi affect seed development? To test the phenotypic consequences of expressing HDG3 from previously repressed Cvi alleles, we compared the phenotypes of seeds from Col x Cvi (low HDG3 expression) and Col x Cvi HDG3 IR (2-4-fold increased HDG3 expression) seeds by sectioning and staining (Fig 6). In crosses with the HDG3 IR lines, endosperm cellularization occurred at a significantly earlier stage of embryo development, where it was observed as early as the globular stage of embryogenesis (Fig 6A and 6B, S2 and S3 Table). Whereas endosperm development appeared accelerated, embryo development was significantly delayed (Fig 6B, S2 and S3 Table). The effect on endosperm cellularization was also observed in Cvi x Cvi HDG3 IR F1 seeds, although to a lesser extent (S2, S3 Table). Mature selfed seeds from Cvi HDG3 IR plants weighed significantly less than selfed seeds from Cvi and had reduced area (Fig 6C and 6D). This is consistent with known correlations between early endosperm cellularization and the production of smaller seeds [35–37]. These observations support the hypothesis that hypomethylation and repression of the Cvi HDG3 allele is important for Cvi-directed developmental programs and that epiallelic variation contributes to the natural variation in seed development in Arabidopsis.
Fig 6. Effects of HDG3 imprinting on Cvi seed development.

(A) Aniline blue and safranin O staining of seed sections at 5 DAP from the indicated F1 seeds. Scale bars, 50 μm. (B) Phenotypic characterization of sectioned seeds, assaying degree of endosperm cellularization relative to embryo stage. (C) Seed weight in selfed Cvi and Cvi HDG3 IR plants. Individual data points and mean +/- SD shown. P-value from unpaired two tailed t-test. (D) Seed area for self-fertilized Cvi (n = 287 seeds), Cvi HDG3 IR 2–5 (n = 496) and Cvi HDG3 IR 3–4 (n = 386). Differences between IR seeds and Cvi are significant at p < 2.2e-16 as determined by Welch’s two-tailed t-test.
Discussion
The establishment, maintenance, and inheritance of DNA methylation are fairly well understood processes. Disruption of methylation patterns by mutations in DNA methyltransferase enzymes have clear gene expression consequences. However, whether or not methylation is regulatory during development–meaning that dynamic loss or gain of methylation is a normal aspect of gene regulation–is less well understood. An exception to this is in the endosperm, where active DNA demethylation in the female gamete before fertilization establishes differential DNA methylation after fertilization, a step that is essential for normal seed development [38]. We thus hypothesized that the phenotypic impact of naturally occurring epialleles might be particularly evident in the endosperm, because the differential methylation between maternal and paternal alleles that is required for gene imprinting could be variable across accessions [20]. We have shown that HDG3 represents a case study of this proposed phenomenon. By placing a methylation trigger in Cvi (the HDG3 IR transgene), we were able to convert the Cvi HDG3 allele from a hypomethylated to a methylated state. This switch in methylation was sufficient to promote expression of the paternally inherited Cvi HDG3 allele in endosperm to 3-fold higher levels. Because we altered methylation at the endogenous HDG3 Cvi locus, which retains all DNA sequence polymorphisms, we have shown that methylation variation alone is sufficient to cause expression, and thus imprinting, variation. However, our results also show that it is unlikely that methylation of the proximal TE accounts for all of the expression differences between paternal Col and Cvi HDG3 alleles in endosperm. The paternally inherited methylated Cvi allele, while more highly expressed than paternally inherited naturally hypomethylated Cvi allele, was not as highly expressed as paternally inherited methylated Col alleles in endosperm (Figs 1 and 5). Additional cis genetic or trans genetic or epigenetic variation likely also affects HDG3 expression levels. Finally, it is not possible to determine from the experiments presented here whether the original difference in methylation between naturally methylated and non-methylated alleles lacks any genetic basis. Cvi lacks the small RNAs associated with the 5’ TE that are found in many other accessions, but the ultimate cause of this difference remains unknown (S4 Fig).
Our experiments also shed light on the relative receptiveness of maternal and paternal endosperm genomes to de novo methylation. The HDG3 inverted repeat transgene should create endosperm small RNAs that are homologous to both Col and Cvi alleles (there are only 4 SNPs and a 3 bp indel between Col and Cvi in the IR target region). Yet, in endosperm from Col x Cvi HDG3 IR crosses, the paternally inherited Cvi allele had high levels of non-CG methylation, whereas the maternally inherited Col alleles remained hypomethylated despite the presence of the IR transcript (Fig 4, S5 Fig, S6 Fig). In contrast, F1 embryos from the same crosses were indeed more highly methylated in the non-CG context on maternal Col alleles compared to maternal Col alleles from Col x Cvi crosses (S6 Fig). Thus, maternally inherited HDG3 alleles in endosperm are refractory to de novo methylation even when a methylation trigger is present, in contrast to maternally inherited HDG3 alleles in embryos. These results further support findings that once a region is actively demethylated on the maternally inherited endosperm genome, it is “protected” from de novo methylation even when triggering small RNAs are present [31].
Finally, although the direct targets of the HDG3 transcription factor are still unknown, we have shown that natural variation in HDG3 expression (expressed in Col, low expression in Cvi) has consequences for seed gene expression programs and development (Figs 2, 3, 6 and 7). Expression of HDG3 in seeds fathered by Cvi caused dramatically earlier endosperm cellularization and the seeds were smaller and lighter at maturity (Fig 6). These findings are consistent with class IV HD-ZIP genes inhibiting the cell cycle and promoting cellular differentiation [24,27]. However, mutation of hdg3 in Col, while displaying the predicted opposite effect on endosperm cellularization timing, also resulted in smaller seeds weighing slightly less than wild-type (Figs 2, 6 and 7). Although the effects on final seed size are seemingly contradictory and the physiological basis remains incompletely understood, these results are predicted under the aegis of the kinship theory [7]. The theory predicts that PEGs promote maternal investment in offspring, which is consistent with the effects of the hdg3 mutation in Col (i.e. less maternal investment results in smaller seeds). Our results suggest that this effect is specific to a Col seed developmental program. In Cvi endosperm, expression of HDG3 is seemingly maladaptive, leading to the production of smaller seeds. Cvi naturally produces much larger seeds than Col or Ler, although fewer in number [20,22,33] (Figs 2 and 6). Our results suggest that the loss of HDG3 expression in Cvi was an important part of the evolutionary process that resulted in these phenotypic differences.
Fig 7. Schematic summary of relative seed development at 5 DAP.

Shapes represent phenotypic space occupied by the indicated genotypes.
In summary, we have demonstrated that seed phenotypic differences can be caused by methylation differences at single genes. This study provides further evidence that epigenetic differences underlie developmental adaptations in plants. We have previously shown that the imprinting status of many genes varies between accessions; our current study argues that intraspecific variation in imprinting is an important determinant of seed developmental variation.
Materials and methods
Plant material
The SALK insertion mutant was obtained from the Arabidopsis Biological Resource Center [39]. hdg3-1 (SALK_033462) was backcrossed to Col-0 three times before experimentation. For experiments comparing or crossing wild-type and hdg3 mutant plants, plants were F3 segregants from selfed progeny of HDG3/hdg3-1. Plants were grown in a growth chamber or greenhouse with 16-hour days at 22° C. For crosses, flowers were emasculated and then pollinated after 2 days.
In situ hybridization
Controlled floral pollinations were performed for each specified cross. At least two independent in situ experiments were performed for each genotype. Siliques were harvested 5 or 6 days after pollination (DAP) and fixed in FAA overnight at 4°C. Following dehydration and clearing (HistoClear, National Diagnostics), samples were embedded in Paraplast Plus (McCormick Scientific), and sectioned at 9 μM (Leica RM 2065 rotary microtome). Ribbons were mounted with DEPC water on ProbeOn Plus slides (Fisher) at 42°C and dried overnight at 37°C. For probes, a 278 bp region of HDG3 (S5 Table) and previously published 602 bp probe for PDF1 [40] were amplified from endosperm cDNA and cloned into P-GEM T vectors (Promega). Plasmids containing sense and antisense oriented fragments were identified and linear templates were amplified using M13 forward and reverse primers for probe synthesis. Antisense and sense RNA probes were synthesized in vitro with digoxigenin-UTPs using T7 or SP6 polymerase (DIG RNA labeling kit, Roche/Sigma Aldrich). Probes were subsequently hydrolyzed and dot blots were performed to estimate probe concentration. Pre-hybridization steps were preformed according to [41] except Pronase digestion occurred for 15 minutes at 37°C. Hybridization and post-hybridizations were performed according to [42], with minor modifications. For higher confidence in directly comparing expression patterns, slides corresponding to the cross and its reciprocal were processed face to face in the same pairs for hybridization, antibody, and detection steps. Negative controls consisted of hybridizing sense probes to wild-type tissue and antisense probes to hdg3 tissue. The sense probe lacked signal (S1 Fig). A probe to PDF1, which is expressed in the L1 embryo layer [43], served as a positive control for successful in situ hybridization (S1 Fig). Hybridization was performed overnight at 55°C, slides were then washed twice in 0.2X SSC for 60 mins each at 55°C, then twice in NTE for 5 min at 37°C and RNaseA treated for 20 min at 37°C, followed by two more 5 min NTE washes. Slides were incubated at room temperature for 1 hour with Anti-DIG antibody (Roche/Sigma Aldrich) diluted 1:1250 in buffer A [42]. Slides were then washed four times for 20 min each at room temperature with buffer A and once for 5 min with detection buffer [42]. Colorimetric detections were performed using NBT/BCIP Ready-To-Use Tablets (Roche/Sigma Aldrich) dissolved in water. Slides were allowed to develop 16–24 hours before stopping. Slides were dehydrated, mounted in Permount (Electron Microscopy Sciences) and imaged using a Zeiss Axio Imager M2. Minor level adjustments and smart sharpen were applied to images to compensate for image transfer from live to digital (Adobe Photoshop).
Seed staining
Plant material was fixed and embedded as previously described and sectioned at 9 μm. Slides were dewaxed twice in xylenes for 5 minutes, rehydrated through a graded ethanol/0.85% salt series from 100%-30%, 1 minute each, stained in 0.6% Safranin O Solution (Cat# 2016–03, Sigma Aldrich) for 5 minutes, washed with water, stained with a saturated 2.5% Aniline blue (Harleco–EMD Millipore, #128–12) in 2% glacial acetic acid aqueous solution for 3 minutes, washed with water, rapidly dehydrated though graded ethanol/salt series to 100%, 5 seconds for each step, and then twice in xylenes for 5 minutes each. Slides were briefly drained, cover slipped and mounted with Cytoseal™ 60 (Thermo Scientific) and imaged using a Zeiss Axio Imager M2.
Seed phenotypic analysis
Previously processed slides from double staining and in situ hybridization experiments were re-examined and used for embryo and endosperm developmental analyses. Using previously published endosperm cellularization and embryogenesis stages [34,44], individual seeds at 5 DAP were scored first for embryo stage and then for respective endosperm stage. Endosperm stage was given a numerical score (-3 to +5) depending on the relative stage of endosperm cellularization compared to the expected endosperm cellularization stage given the embryo stage. Individual seeds with matching embryogenesis and endosperm cellularization stages were scored “normal” and ranged from 0–1; seeds that were scored “early” were defined as being +1.5 to +5 stages further along in the cellularization process compared to normal. Seeds that were scored “delayed” were defined as being -1 to -3 stages behind in the cellularization process compared to normal. To determine whether any developmental differences in endosperm cellularization or embryogenesis were statistically significant, we implemented the asymptotic generalized Pearson chi-squared test from the coin package [45] in R with default scoring weights (S3 Table). Developmental stage was treated as an ordinal variable, while cross genotype was treated as a non-ordinal, nominal variable. Pairwise comparisons were carried out with the R function pairwiseOrdinalIndependence from the rcompanion package. For all tests, embryo development data was collapsed into three categories young (pre-globular to globular), middle (late globular to early heart), and older (heart to torpedo) and detailed endosperm cellularization data was collapsed into the categories delayed, normal, and early.
Inverted repeat transgene
The 450 bp sequence 5' of HDG3 corresponding to a fragment of AT2TE60490 from Chr2: 13740010–13740460 was amplified from Cvi (S5 Table) and cloned into the directional entry vector pENTR-TOPO-D (Invitrogen). The sequence was then inserted twice in an inverted repeat conformation into the vector pFGCGW [46] with a LR clonase reaction (Invitrogen). Cvi plants were transformed with the inverted repeat transgene by floral dipping and T1 lines were screened for DNA methylation using bisulfite sequencing. T3 plants homozygous for the IR transgene and with a methylated HDG3 5’ region in leaves, or their T4 progeny, were identified and used for subsequent experiments.
Quantitative RT-PCR
RNA was isolated from endosperm dissected from seeds at 6 or 7 DAP as described [47] using the RNAqueous Micro Kit (Ambion, Life Technologies Corporation). DNAse I-treated RNA (Invitrogen, Life Technologies Corporation) was used for cDNA synthesis with oligo-dT primer using Superscript II reverse transcriptase (Invitrogen) according to manufacturer’s instructions. Quantitative RT-PCR (RT-qPCR) was performed using Fast SYBR Green Master Mix or TaqMan Gene Expression Master Mix (Applied Biosystems). All reactions were performed in three or four technical replicates using a StepOne Plus Real-Time PCR system (Applied Biosystems). For SYBR Green based assays, relative expression was calculated using the ddCt method as described [48]. The reference gene was AT1G58050 [49]. For allele-specific expression in Col-Cvi crosses, a multiplex TaqMan assay was developed by designing primers and PrimeTime® Double-quenched Custom Probes with online tool http://www.idtdna.com/pages/products/gene-expression/custom-qpcr-probes. Cycling conditions were 15 cycles: 95°C for 15 seconds, 63°C for 30 seconds, 72°C for 30 seconds followed by 25 cycles: 95°C for 15 seconds, 63°C for 30 seconds with touchdown 0.05°C/cycle, and 72°C for 30 seconds. The relative expression of each allele within each genotype was calculated using a standard curve (R2 value >0.99) as reference. Primer and probe sequences are in S5 Table.
Bisulfite sequencing
Genomic DNA was isolated from leaves, endosperm, and embryo at 6 or 7 days after pollination using a CTAB procedure. Bisulfite treatment was performed using the MethylCode Bisulfite Conversion Kit (Invitrogen, Life Technologies Corporation) or BisulFlash DNA Bisulfite Conversion Easy Kit (Epigentek Group Inc.) following the manufacturer’s protocols. 2 μl bisulfite treated DNA was used in PCR reactions with 2.5 U ExTaq DNA polymerase (Takara) and 0.4 μM primers using the following cycling conditions (95°C 3 minutes, 40 cycles of [95°C for 15 seconds, 50°C for 30 seconds, 72°C for 45 seconds], 72°C for 5 minutes). PCR products were gel purified, cloned using a TOPO-TA (Invitrogen) or CloneJet (Life Technologies) PCR cloning kit and individual colonies were sequenced. Sequences were aligned using SeqMan and methylation was quantified using CyMate [50].
mRNA-seq
RNA was isolated from endosperm of Col-0, hdg3-1 and Col-0 x Cvi seeds at 7 DAP as described above. Three replicates for each cross were obtained. DNAse treated RNA was used as input for the SmartSeq Clontech Ultralow RNA-Seq kit. Libraries were constructed by the Genome Technology Core at Whitehead Institute. Six libraries were multiplexed per lane in a Hi-Seq 2500 Standard mode, 40 base, single read run. Each replicate was sequenced to a depth of between 33 and 41 million reads. Reads were processed with Trim_galore using the command “trim_galore -q 25—phred64—fastqc—stringency 5—length 18”. Processed reads were aligned to the TAIR10 genome with Tophat2 [51] using the command “tophat -i 30 -I 3000 —b2-very-sensitive—solexa1.3-quals -p 5—segment-mismatches 1—segment-length 18”. Differential gene expression was detected with Cuffdiff2 [52] and the ARAPORT11 annotation (S1 and S2 Tables). Reads are deposited in GEO GSE118371.
Supporting information
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(XLSX)
(PDF)
(PDF)
(XLSX)
(PDF)
Acknowledgments
We thank R. Jaenisch and P. Reddien for sharing equipment for in situ hybridization, B. Williams for comments on the manuscript, and R. Povilus for assistance with statistical analysis.
Data Availability
All mRNA-seq files are available from the GEO database (accession number GSE118371). All other relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was funded by the National Science Foundation, Division of Molecular and Cellular Biosciences (https://www.nsf.gov/div/index.jsp?div=MCB) CAREER grant 1453459 to MG. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Gehring M, Huh JH, Hsieh T-F, Penterman J, Choi Y, Harada JJ, et al. DEMETER DNA glycosylase establishes MEDEA polycomb gene self-imprinting by allele-specific demethylation. Cell. 2006; 124(3):495–506. 10.1016/j.cell.2005.12.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gehring M, Bubb KL, Henikoff S. Extensive demethylation of repetitive elements during seed development underlies gene imprinting. Science. 2009; 324(5933):1447–51. 10.1126/science.1171609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hsieh T-F, Ibarra CA, Silva P, Zemach A, Eshed-Williams L, Fischer RL, et al. Genome-wide demethylation of Arabidopsis endosperm. Science. 2009; 324(5933):1451–4. 10.1126/science.1172417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Satyaki PRV, Gehring M. DNA methylation and imprinting in plants: machinery and mechanisms. Crit Rev Biochem Mol Biol. 2017; 52(2):163–75. 10.1080/10409238.2017.1279119 [DOI] [PubMed] [Google Scholar]
- 5.Haig D. The kinship theory of genomic imprinting. Annu Rev Ecol Syst. 2000; 31: 9–32. [Google Scholar]
- 6.Haig D. Kin conflict in seed development: an interdependent but fractious collective. Annu Rev Cell Dev Biol. 2013; 29:189–211. 10.1146/annurev-cellbio-101512-122324 [DOI] [PubMed] [Google Scholar]
- 7.Patten MM, Ross L, Curley JP, Queller DC, Bonduriansky R, Wolf JB. The evolution of genomic imprinting: theories, predictions and empirical tests. Heredity. 2014; 113(2):119–28. 10.1038/hdy.2014.29 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Klosinska M, Picard CL, Gehring M. Conserved imprinting associated with unique epigenetic signatures in the Arabidopsis genus. Nat Plants. 2016; 2(10):16145 10.1038/nplants.216.145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Patten MM. Epigenetics: Imprinting evolution in Arabidopsis. Nat Plants. 2016; 2(10):16152 10.1038/nplants.2016.152 [DOI] [PubMed] [Google Scholar]
- 10.Schmitz RJ, Schultz MD, Urich MA, Nery JR, Pelizzola M, Libiger O, et al. Patterns of population epigenomic diversity. Nature. 2013; 495(7440):193–8. 10.1038/nature11968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kawakatsu T, Huang S-SC, Jupe F, Sasaki E, Schmitz RJ, Urich MA, et al. Epigenomic Diversity in a Global Collection of Arabidopsis thaliana Accessions. Cell. 2016; 166(2):492–505. 10.1016/j.cell.2016.06.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Weigel D, Colot V. Epialleles in plant evolution. Genome Biol. 2012; 13(10):249 10.1186/gb-2012-13-10-249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Aller EST, Jagd LM, Kliebenstein DJ, Burow M. Comparison of the Relative Potential for Epigenetic and Genetic Variation To Contribute to Trait Stability. G3 (Bethesda). 2018; 8(5):1733–46. 10.1534/g3.118.200127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Durand S, Bouché N, Perez Strand E, Loudet O, Camilleri C. Rapid Establishment of Genetic Incompatibility through Natural Epigenetic Variation. Current Biology. 2012; 22(4):326–31. 10.1016/j.cub.2011.12.054 [DOI] [PubMed] [Google Scholar]
- 15.Cubas P, Vincent C, Coen E. An epigenetic mutation responsible for natural variation in floral symmetry. Nature. 1999; 401(6749):157–61. 10.1038/43657 [DOI] [PubMed] [Google Scholar]
- 16.He L, Wu W, Zinta G, Yang L, Wang D, Liu R, et al. A naturally occurring epiallele associates with leaf senescence and local climate adaptation in Arabidopsis accessions. Nat Commun. 2018; 9(1):460 10.1038/s41467-018-02839-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Martin A, Troadec C, Boualem A, Rajab M, Fernandez R, Morin H, et al. A transposon-induced epigenetic change leads to sex determination in melon. Nature. 2009; 461(7267):1135–8. 10.1038/nature08498 [DOI] [PubMed] [Google Scholar]
- 18.Quadrana L, Almeida J, Asís R, Duffy T, Dominguez PG, Bermúdez L, et al. Natural occurring epialleles determine vitamin E accumulation in tomato fruits. Nat Commun. 2014; 5:3027–7. 10.1038/ncomms5027 [DOI] [PubMed] [Google Scholar]
- 19.Manning K, Tör M, Poole M, Hong Y, Thompson AJ, King GJ, et al. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat Genet. 2006; 38(8):948–52. 10.1038/ng1841 [DOI] [PubMed] [Google Scholar]
- 20.Pignatta D, Erdmann RM, Scheer E, Picard CL, Bell GW, Gehring M. Natural epigenetic polymorphisms lead to intraspecific variation in Arabidopsis gene imprinting. Elife. 2014; 3:e03198 10.7554/eLife.03198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Waters AJ, Bilinski P, Eichten SR, Vaughn MW, Ross-Ibarra J, Gehring M, et al. Comprehensive analysis of imprinted genes in maize reveals allelic variation for imprinting and limited conservation with other species. Proc Natl Acad Sci USA. 2013; 110(48):19639–44. 10.1073/pnas.1309182110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Alonso-Blanco C, Blankestijn-De Vries H, Hanhart CJ, Koornneef M. Natural allelic variation at seed size loci in relation to other life history traits of Arabidopsis thaliana. Proc Natl Acad Sci USA. 1999; 96(8):4710–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nakamura M, Katsumata H, Abe M, Yabe N, Komeda Y, Yamamoto KT, et al. Characterization of the class IV homeodomain-Leucine Zipper gene family in Arabidopsis. Plant Physiol. 2006; 141(4):1363–75. 10.1104/pp.106.077388 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Robinson DO, Roeder AHK. Themes and variations in cell type patterning in the plant epidermis. Current Opinion in Genetics & Development. 2015; 32:55–65. doi: 10.106/j.gde/2015.01.008 [DOI] [PubMed] [Google Scholar]
- 25.Abe M, Katsumata H, Komeda Y, Takahashi T. Regulation of shoot epidermal cell differentiation by a pair of homeodomain proteins in Arabidopsis. Development. 2003; 130(4):635–43. [DOI] [PubMed] [Google Scholar]
- 26.Roeder AHK, Cunha A, Ohno CK, Meyerowitz EM. Cell cycle regulates cell type in the Arabidopsis sepal. Development. 2012; 139(23):4416–27. 10.1242/dev.082925 [DOI] [PubMed] [Google Scholar]
- 27.Horstman A, Fukuoka H, Muino JM, Nitsch L, Guo C, Passarinho P, et al. AIL and HDG proteins act antagonistically to control cell proliferation. Development. 2015; 142(3):454–64. 10.1242/dev.117168 [DOI] [PubMed] [Google Scholar]
- 28.Kinoshita T, Miura A, Choi Y, Kinoshita Y, Cao X, Jacobsen SE, et al. One-Way Control of FWA Imprinting in Arabidopsis Endosperm by DNA Methylation. Science. 2004; 303(5657):521–3. 10.1126/science.1089835 [DOI] [PubMed] [Google Scholar]
- 29.Hsieh T-F, Shin J, Uzawa R, Silva P, Cohen S, Bauer MJ, et al. Regulation of imprinted gene expression in Arabidopsis endosperm. Proc Natl Acad Sci USA. 2011; 108(5):1755–62. 10.1073/pnas.1019273108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wolff P, Jiang H, Wang G, Santos-González J, Köhler C. Paternally expressed imprinted genes establish postzygotic hybridization barriers in Arabidopsis thaliana. Elife. 2015; 4:e10074 10.7554/eLife.10074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Erdmann RM, Satyaki PRV, Klosinska M, Gehring M. A Small RNA Pathway Mediates Allelic Dosage in Endosperm. Cell Rep. 2017; 21(12):3364–72. 10.1016/j.celrep.2017.11.078 [DOI] [PubMed] [Google Scholar]
- 32.Singh DP, Jermakow AM, Swain SM. Gibberellins are required for seed development and pollen tube growth in Arabidopsis. Plant Cell. 2002; 14(12):3133–47. 10.1105/tpc.003046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Herridge RP, Day RC, Baldwin S, Macknight RC. Rapid analysis of seed size in Arabidopsis for mutant and QTL discovery. Plant Methods. 2011; 7(1):3 10.1186/1746-4811-7-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Boisnard-Lorig C, Colon-Carmona A, Bauch M, Hodge S, Doerner P, Bancharel E, et al. Dynamic analyses of the expression of the HISTONE::YFP fusion protein in arabidopsis show that syncytial endosperm is divided in mitotic domains. Plant Cell. 2001; 13(3):495–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Scott RJ, Spielman M, Bailey J, Dickinson HG. Parent-of-origin effects on seed development in Arabidopsis thaliana. Development. 1998; 125(17):3329–41. [DOI] [PubMed] [Google Scholar]
- 36.Garcia D, Saingery V, Chambrier P, Mayer U, Jürgens G, Berger F. Arabidopsis haiku mutants reveal new controls of seed size by endosperm. Plant Physiol. 2003; 131(4):1661–70. 10.1104/pp.102.018762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hehenberger E, Kradolfer D, Kohler C. Endosperm cellularization defines an important developmental transition for embryo development. Development. 2012; 139(11):2031–9. 10.1242/dev.077057 [DOI] [PubMed] [Google Scholar]
- 38.Gehring M, Satyaki PR. Endosperm and Imprinting, Inextricably Linked. Plant Physiol. 2017; 173(1):143–54. 10.1104/pp.16.01353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science. 2003; 301(5633):653–7. 10.1126/science.1086391 [DOI] [PubMed] [Google Scholar]
- 40.Kunieda T, Mitsuda N, Ohme-Takagi M, Takeda S, Aida M, Tasaka M, et al. NAC family proteins NARS1/NAC2 and NARS2/NAM in the outer integument regulate embryogenesis in Arabidopsis. Plant Cell. 2008; 20(10):2631–42. 10.1105/tpc.108.060160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jackson D. In situ hybridization in plants Molecular plant pathology: a practical approach. Oxford University Press; 1991. [Google Scholar]
- 42.Bortiri E, Chuck G, Vollbrecht E, Rocheford T, Martienssen R, Hake S. ramosa2 encodes a LATERAL ORGAN BOUNDARY domain protein that determines the fate of stem cells in branch meristems of maize. Plant Cell. 2006; 18(3):574–85. 10.1105/tpc.105.039032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tanaka H, Watanabe M, Sasabe M, Hiroe T, Tanaka T, Tsukaya H, et al. Novel receptor-like kinase ALE2 controls shoot development by specifying epidermis in Arabidopsis. Development. 2007; 134(9):1643–52. 10.1242/dev.003533 [DOI] [PubMed] [Google Scholar]
- 44.Berger F. Endosperm: the crossroad of seed development. Current Opinion in Plant Biology. 2003; 6(1):42–50. [DOI] [PubMed] [Google Scholar]
- 45.Hothorn T, Hornik K, van de Wiel MA, Zeileis A. A Lego System for Conditional Inference. The American Statistician. 2012; 60(3):257–63. [Google Scholar]
- 46.Zhang C, Galbraith DW. RNA interference-mediated gene knockdown within specific cell types. Plant Molecular Biology. 2012; 80(2):169–76. 10.1007/s11103-012-9937-7 [DOI] [PubMed] [Google Scholar]
- 47.Gehring M, Missirian V, Henikoff S. Genomic Analysis of Parent-of-Origin Allelic Expression in Arabidopsis thaliana Seeds. PLoS ONE. 2011; 6(8):e23687 10.1371/journal.pone.0023687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001; 25(4):402–8. 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- 49.Czechowski T, Stitt M, Altmann T, Udvardi MK, Scheible W-R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005; 139(1):5–17. 10.1104/pp.105.063743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hetzl J, Foerster AM, Raidl G, Mittelsten Scheid O. CyMATE: a new tool for methylation analysis of plant genomic DNA after bisulphite sequencing. Plant J. 2007; 51(3):526–36. 10.1111/j.1365-313X.2007.03152.x [DOI] [PubMed] [Google Scholar]
- 51.Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013; 14(4):R36 10.1186/gb-2013-14-4-r36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Trapnell C, Hendrickson DG, Sauvageau M, Goff L, Rinn JL, Pachter L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat Biotechnol. 2013; 31(1):46–53. 10.1038/nbt.2450 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(PDF)
(XLSX)
(PDF)
(PDF)
(XLSX)
(PDF)
Data Availability Statement
All mRNA-seq files are available from the GEO database (accession number GSE118371). All other relevant data are within the manuscript and its Supporting Information files.