

Abstract
The effect of [10]-gingerol on cytosol free Ca2+ concentration ([Ca2+]i) and viability is large unknown. This study examines the early signaling effects of [10]-gingerol on human colorectal cancer cells. It was found that this compound caused a slow and sustained rise of [Ca2+]i in a concentration-dependent manner. [10]-Gingerol also induced a [Ca2+]i rise when extracellular Ca2+ was removed, but the magnitude was reduced by 38%. In a Ca2+-free medium, the [10]-gingerol-induced [Ca2+]i rise was partially abolished by depleting stored Ca2+ with thapsigargin (an endoplasmic reticulum Ca2+ pump inhibitor). The elevation of [10]-gingerol-caused [Ca2+]i in a Ca2+-containing medium was not affected by modulation of protein kinase C activity. The [10]-gingerol-induced Ca2+ influx was insensitive to L-type Ca2+ channel blockers. At concentrations of 10-100 μM, [10]-gingerol killed cells in a concentration-dependent manner. These findings suggest that [10]-gingerol induces [Ca2+]i rise by causing Ca2+ release from the endoplasmic reticulum and Ca2+ influx from non-L-type Ca2+ channels in SW480 cancer cells.
Keywords: Ca2+, [10]-Gingerol, L-type Ca2+ channel blockers, SW480 cells, Thapsigargin
Introduction
Ginger (Zingiber officinale L. Zingiberaceae) is a common condiment for various foods and beverages. Ginger also has a long history of use in traditional medicine. The underground stems or rhizomes of this plant have been used as a medicine in East Asian, Indian, and Arabic herbal traditions since ancient times [1]. In mainland China, the rhizomes of ginger have been used in oriental medicine for the treatment of the common cold, disorders of the gastrointestinal tract, neuralgia, rheumatism, colic, and motion discomfort [2,3]. The non-volatile pungent ingredients from ginger include gingerol, shogaol and zingerone. Recently, several population-based studies have shown that persons in Southeast Asian countries have a much lower risk of colon, gastrointestinal, prostate, breast, and other cancers than those in European and American countries [4]. It is believed that constituents of their diet may play important roles in cancer prevention. Indeed, some phenolic substances present in fruit and vegetables, and in medicinal plants, have potential cancer chemopreventive activities, as supported by both in vitro and in vivo in experiments [2,5,6,7]. These agents are known to have the ability to suppress the transformative, hyperproliferative, and inflammatory processes of carcinogenesis.
The phenolic compounds derived from ginger possess many interesting pharmacological and physiological activities. For example, [6]-gingerol [1-(4′-hydroxy-3′-methoxyphenyl)-5-hydroxy-3- decanone], the major pungent principle of ginger, has potential anti-inflammatory, antioxidant, anticarcinogenic, and antimutagenic activities [8,9,10]. Evidence indicates that [6]-gingerol exerts an inhibitory effect on DNA synthesis, also causes apoptosis in human promyelocytic leukemia (HL-60) cells [11]. In vitro, [6]-gingerol inhibited both the VEGF- and bFGF-induced proliferation of human endothelial cells and caused cell-cycle arrest in the G1 phase [12]. This compound also induced [Ca2+]i elevation and was cytotoxic to canine renal cells [13]. In the case of [10]-gingerol (Figure 1), its effect on human promyelocytic leukemia (HL-60) cells is better than [6]-gingerol’s [11] and the activity of sarcoplasmic reticulum of Ca2+-ATPase could be stimulated by [10]-gingerol [14]. However, the detailed mechanism of [10]-gingerol’s anticarcinogenic effects is still unclear.
Figure 1.

Structure of [10]-gingerol.
The effect of [10]-gingerol on Ca2+ signaling and cytotoxicity in human colon cancer SW480 cells has not been explored. Colorectal cancer is the third most frequent and second most lethal in the United States [15]. Therefore, there is a need to search more effective chemotherapeutic agents that can be used to remedy the patients who have failed to respond under traditional chemotherapy. This study was performed to elucidate whether [10]-gingerol affects human colorectal tumorigenesis. Using fura-2 as a fluorescent Ca2+ indicator, we report for the first time that [10]-gingerol induced a significant and prolonged [Ca2+]i increase and cytotoxicity in human colorectal cancer cells. The concentration-response relationship, the Ca2+ sources of the Ca2+ signal, and the role of protein kinase C in the signal have been investigated.
Results and Discussion
Effect of [10]-gingerol on [Ca2+]i
[10]-Gingerol at concentrations between 5-25 μM increased [Ca2+]i in a concentration- dependent manner in the presence of extracellular Ca2+. Figure 2A shows typical recordings of the [Ca 2+]i rise induced by 5-25 μM [10]-gingerol.
Figure 2.
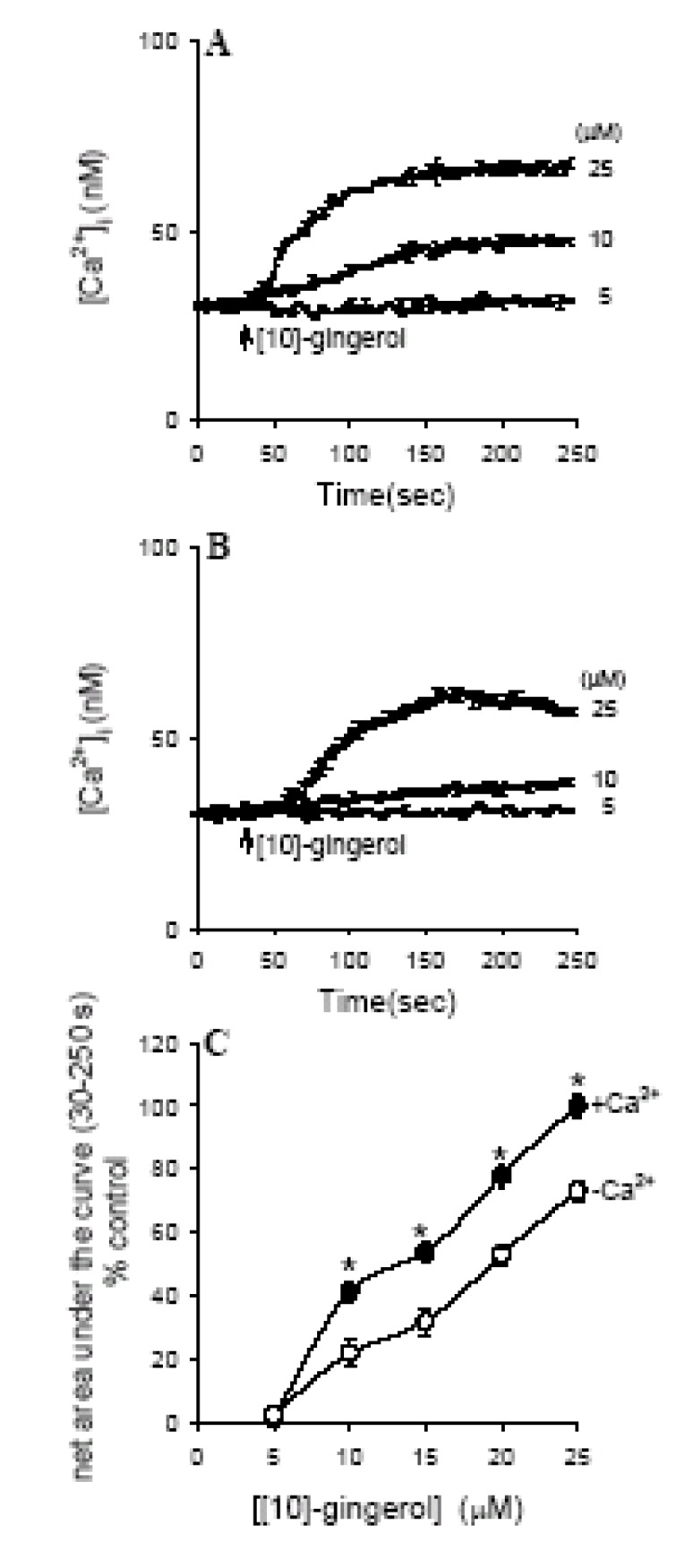
Effects of [10]-gingerol on [Ca2+]i in SW480 cells. (A) Concentration-dependent effects of [10]-gingerol, with the concentration of the reagent indicated. Experiments were performed in Ca2+-containing medium. [10]-Gingerol was added at 30 sec and was present throughout the measurements for 250 sec. (B) Effect of extracellular Ca2+ removal on [10]-gingerol-induced [Ca2+]i elevation. The concentration of [10]-Gingerol is indicated. (C) Concentration-response plots of [10]-gingerol-induced [Ca2+]i rises in Ca2+-containing medium (filled circles) and Ca2+-free medium (open circles). The data are presented as the percentage of control, which is the net [Ca2+]i rise induced by 25 μM [10]-gingerol in Ca2+-containing medium. Data are mean SEM of five experiments (*p < 0.05 compared to open circles).
At a concentration of 0.1 μM, [10]-gingerol had no effect on [Ca2+]i (i.e., equivalent to baseline, 0 μM). The [Ca2+]i rise induced by 5-25 μM [10]-gingerol comprised an immediate rise and a sustained phase in 250 sec. At a concentration of 25 μM, the [Ca2+]i rise had a net value of 752nM at 250 sec. Figure 2C the [10]-gingerol-induced response.
Effect of removing extracellular Ca2+ on [10]-gingerol-induced [Ca2+]i signals
Experiments were performed to evaluate the relative contribution of extracellular Ca2+ entry and Ca2+ release from stores in the [10]-gingerol response. Figure 2B shows that removal of extracellular Ca2+ largely suppressed the [10]-gingerol-induced [Ca2+]i elevation. The concentration-response relationship of [10]-gingerol-induced [Ca2+]i rise in the presence and absence of extracellular Ca2+ is shown in Figure 2C. Ca 2+ removal inhibited the [Ca2+]i rise caused by 25 μM [10]-gingerol by 38% as the maximum value (n = 5; p <0.05).
Internal Ca2+ stores for [10]-gingerol-induced [Ca2+]i Rises
Previous reports have shown that the endoplasmic reticulum is a major Ca2+ store in the majority of cells [16]. Figure 3A shows that in Ca2+-free medium, application of 1 μM thapsigargin caused a [Ca2+]i rise that comprised an initial increase and a gradual decay toward baseline. The net maximum [Ca2+]i value was 96±4 nM (n = 5). After depleting the endoplasmic reticulum Ca2+ store with thapsigargin, addition of 25 μM [10]-gingerol did not induce a [Ca2+]i rise, as shown in Figure 2B. Conversely, Figure 3B shows that after preincubation with [10]-gingerol (25 μM) for 250 sec, subsequent addition of 1 μM thapsigargin induced a [Ca2+]i rise with a net value of 72±3 nM (n = 5), which was 25% (p < 0.05), smaller than the control thapsigargin response shown in Figure 3A.
Figure 3.

Intracellular sources of [10]-gingerol-induced [Ca2+]i elevation. Experiments were performed in Ca2+-free medium. Reagents were applied at the times indicated by arrows. (A) Thapsigargin (1 μM) and [10]-gingerol (25 μM) were added at 30 s and 250 s, respectively. (B) [10]-gingerol and thapsigargin were added at 30 s and 250 s, respectively. Data are means SEM of five experiments.
Lack of inhibitory effect of L-type Ca2+ entry blockers on Ca2+ release by [10]-gingerol
To explore the pathways underlying [10]-gingerol-induced Ca2+ entry, the effects of several L-type Ca2+ entry blockers on [10]-gingerol-induced [Ca2+]i rise were evaluated. Figure 4 shows that in Ca2+-containing medium, pretreatment with 1 μM nicardipine did not inhibited 25 μM [10]-gingerol-induced [Ca2+]i elevation (n = 5; p < 0.05). Similarly, the Ca2+ influx was not affected by 1 μM diltiazem, nifedipine, or verapamil (n = 5).
Figure 4.

Effect of Ca2+ channel blockers on [10]-gingerol-induced [Ca2+]i elevation. All experiments were performed in Ca2+-containing medium. The data are presented as the percentage of the control, which is the net area under the curve (30-250 sec) of the [Ca2+]i rise induced by 25 μM [10]-gingerol. Data are means SEM of five experiments.
Regulation of [10]-gingerol-induced Ca2+ influx by Protein Kinase C
A Ca2+ signal could be modulated by the activity of protein kinase C [17,18]. The role of protein kinase C in [10]-gingerol-induced [Ca2+]i elevation was investigated. Figure 5 shows that the 25 μM [10]-gingerol-induced [Ca2+]i elevation was not altered by pretreatment with 10 nM phorbol myristate acetate (PMA, a protein kinase C activator) or 2 μM GF109203 X (a protein kinase C inhibitor) (n = 5).
Figure 5.

Effect of protein kinase C modulator on [10]-gingerol-induced [Ca2+]i elevation. Experiments were performed in Ca2+-containing medium. PMA(10 nM) or GF 109203X (2 μM) was added 1 min prior to 25 μM 10-gingerol. Data are expressed as the percentage of control that is the net area under the curve of 25μM-[10]-gingerol-induced [Ca2+]i rise (30-250 sec interval), and are means SEM of five experiments.
Effect of [10]-gingerol on Cell Viability
Given the acute incubation with [10]-gingerol caused a substantial and lasting (Ca2+)i response, and unregulated (Ca2+)i rises can be linked to cytotoxicity [16,19], experiments were performed to examine the effect of 24 h incubation with [10]-gingerol on the viability of SW480 cells. Colon cancer cells were treated with 0-100 μM 24 h, and the trypan blue method was performed. In the exposure of 10-25 μM [10]-gingerol, cell viability was unchanged. However, at concentrations of 50-100 μM. [10]-gingerol decreased cell viability in a concentration-dependent manner (Figure 6; n=5).
Figure 6.

Cytotoxic effect of [10]-gingerol on human colorectal cancer SW480 cells. The cell viability was determined by the trypan blue exclusion (TBE) method. Data are expressed as the percentage of control ([10]-gingerol was absent).
This study is the first to investigate the effects of [10]-gingerol on [Ca2+]i in colon cancer cells. The results obtained suggest that [10]-gingerol causes a significant concentration-dependent, sustained [Ca2+]i rise in human colorectal SW480 cancer cells. In Ca2+-containing medium, the [Ca2+]i rise induced by [10]-gingerol was sustained without a decay during the 5 min. of measurements. Sustained [Ca2+]i elevations are thought to alter many cellular functions [20]. [10]-Gingerol may affect cell physiology significantly by changing Ca2+ signaling and stimulating Ca2+-coupled bioactive molecules. The results show that the [Ca2+]i rise was contributed to by both intracellular Ca2+ release and extracellular Ca2+ influx, because the signal was suppressed by removal of extracellular Ca2+.
Regarding the Ca2+ depositories of the [10]-gingerol response, the present data show that thapsigargin-sensitive endoplasmic reticulum store appears to play a crucial role because the [10]-gingerol-induced Ca2+ release was partly abolished by depletion of the endoplasmic reticulumic Ca2+ store with thapsigargin, and, conversely, pretreatment with [10]-gingerol also inhibited thapsigargin-induced Ca2+ release. The endoplasmic reticulum is one of the major Ca2+ stores where various proteins and lipids are synthesized and modified [21,22]. Perturbation of endoplasmic reticulum Ca2+ homeostasis, protein misfolding, or oxidative stress can lead to cell death [22,23]. Evidence reveals that reactive oxygen species (ROS) and the oxidation-reduction (redox) state play significant roles in many cytotoxic pathways caused by frequently used antitumor drugs or environmental toxicants [24]. A rise in [Ca2+]i induced by oxidants may activate Ca2+-dependent enzymes such as proteases, nucleases, and phospholipases to facilitate mitochondrial oxidative stress leading to cytotoxicity [25,26]. Exactly how [10]-gingerol releases Ca2+ stored in the endoplasmic reticulum is unclear, but the process seems to be independent on protein kiase C activity because suppression of this protein did not affect [10]-gingerol-induced Ca2+ release. Because [10]-gingerol and thapsigargin share the same Ca2+ stores, [10]-gingerol may very likely release Ca2+ in a manner similarly to thapsigargin by inhibiting endoplasmic reticulum Ca2+ pump.
In Ca2+-free medium, the [10]-gingerol-induced [Ca2+]i elevation displayed a smaller [Ca2+]i increase throughout the measurement period of 250 sec. This suggests that Ca2+ influx contributed not only to the initial increase, but also to the prolonged phase of the [10]-gingerol-induced [Ca2+]i signal in the Ca2+-containing medium. In non-excitable cells, a possible Ca2+ influx pathway is a store-operated Ca2+ entry, a process triggered by depletion of Ca2+ stores [27]. This possibility was not explored due to the lack of selective pharmacological inhibitors for this Ca2+ influx [28]. Thus, it remains possible that Ca2+ entry mechanisms other than depletion-activated channels may be important in Ca2+ influx in non-excitable cells. [10-]-gingerol is cytotoxic in several cell types including human A549, SK-OV-3, SK-MEL-2, and HCT15 tumor cells [34]. The present data indicate that [10]-gingerol at 50 μM or higher concentrations caused concentration-dependent cell death; whereas at lower concentrations it had no cytotoxic effects.
We are the first to demonstrate that in the presence of 10-100 μM [10]-gingerol, the viability of SW480 cells decreased in a concentration-dependent manner. Ca2+ overloading is known to initiate processes leading to cell death [20]. Moreover, Cell death is induced in a Ca2+-dependent or -independent manner, depending on the stimulating agent and cell type [32,33]. Collectively, this study shows that in SW480 cells, [10]-gingerol caused [Ca2+]i elevations in a concentration-dependent manner by evoking protein kiase C-independent Ca2+ release from the endoplasmic reticulum pathway. These effects may play a crucial role in the physiological action of [10]-gingerol.
Experimental
General
Optical rotations were measured with a JASCO DIP-370 digital polarimeter. UV spectra were obtained in MeCN using a JASCO V-530 spectrophotometer. The IR spectra were measured on a Hitachi 260-30 spectrophotometer. 1H (400 MHz, using CDCl3 as solvent for measurement), 13C (100 MHz), DEPT, HETCOR, COSY, NOESY, and HMBC NMR spectra were obtained on a Unity Plus Varian NMR spectrometer. LRFABMS and LREIMS were obtained with a JEOL JMS-SX/SX 102A mass spectrometer or a Quattro GC-MS spectrometer with a direct inlet system. Silica gel 60 (Merck, 230-400 mesh) was used for column chromatography. Precoated silica gel plates (Merck, Kieselgel 60 F-254, 0.20 mm) were used for analytical TLC, and precoated silica gel plates (Merck, Kieselgel 60 F-254, 0.50 mm) were used for preparative TLC. Spots were detected by spraying with 50% H2SO4 and then heating on a hot plate.
Plant Material
The roots of Zingiber officinale (ginger) were purchased from a local market of Kaohsiung in Taiwan in July 2006, which was identified by Dr. Yen-Ray Hsui of the Division of Silviculture, Taiwan Forestry Research Institute, Taipei, Taiwan. A voucher specimen (Hsui-Zo-1) was deposited at Fooyin University.
Extraction and Isolation
The roots (25.6 kg) of Z. officinale were chipped and air-dried and extracted repeatedly with CHCl3 at room temperature. The combined CHCl3 extracts were then evaporated further separated into 20 fractions by column chromatography on silica gel with gradients of n-hexane/CHCl3. Fr. 9 eluted with CHCl3-MeOH (50:1) was next repeatedly subjected to silica gel CC and yielded [10]-gingerol (210 mg) [29].
Cell Culture and Test Compound Treatment
The SW480 cells were obtained from the American Type Culture Collection. Cells were cultured in Dulbecco's modified Eagle's medium. The media were supplemented with 10% heat-inactivated fetal calf serum, 100 units/mL penicillin and 100 μg/mL streptomycin. Cells were kept at 37 °C in 5% CO2-containing humidified air.
Measurement of [Ca2+]i
Trypsinized cells (106/mL) were allowed to recover in the culture medium for 1 h before being loaded with 2 μM fura-2/AM for 30 min at 25 °C in the same medium. The cells were washed once with serum-free DMEM medium and resuspended in Ca2+-containing medium (pH 7.4) containing (mM): NaCl, 140; KCl, 5; MgCl2, 1; CaCl2, 2; HEPES, 5; d-glucose, 5. Fura-2 fluorescence measurements were performed in a water-jacketed cuvette (25 °C) with continuous stirring; the cuvette contained 1 mL of medium and 0.5 million cells. Fluorescence was monitored with a Shimadzu RF-5301PC spectrofluorophotometer (Kyoto, Japan) by recording the excitation signals at 340 and 380 nm and the emission signal at 510 nm at 1-sec intervals. Maximum and minimum fluorescence values were obtained by adding 0.1% Triton X-100 and 10 mM EGTA sequentially at the end of each experiment. [Ca2+]i was calculated as described previously assuming a Kd of 155 nM [30]. In experiments that were performed in the absence of extracellular Ca2+, cells were bathed in Ca2+-free medium in which CaCl2 (2 mM) was substituted with 0.1 mM EGTA.
Cell viability assay
Cell viability was determined by the trypan blue exclusion (TBE) method [31]. Briefly, 10 μL of 4% trypan blue was added to 90 μL of cell suspension, followed by examination with a hemocytometer under an inverted microscope. Cells which excluded the dye were considered viable and the data were expressed as a percentage of that of the corresponding control group.
Data analysis
Data is presented as means ± standard deviation (SD) and analyzed using one-way ANOVA with Scheffe′s test. A p value of less than 0.05 was considered as statistically significant.
Acknowledgements
This investigation was partly supported by grants from National Science Council (NSC 97-2320-B-242-002-MY3) to C.-Y. C. This work was also supported by grants from National Science Council (NSC93-2311-B-242-002) to S.-Y. K.
Footnotes
Sample Availability: Not available.
References
- 1.Surh Y. Molecular mechanisms of chemopreventive effects of selected dietary and medicinal phenolic substances. Muta. Res. 1999;428:305–327. doi: 10.1016/S1383-5742(99)00057-5. [DOI] [PubMed] [Google Scholar]
- 2.Altman R.D., Marcussen K.C. Effects of a ginger extract on knee pain in patients with osteoarthritis. Arth. Rheum. 2001;44:2531–2538. doi: 10.1002/1529-0131(200111)44:11<2531::AID-ART433>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 3.Grant K.L., Lutz R.B. Alternative therapies: ginger. Am. J. Health-Syst. Pharm. 2000;57:945–947. doi: 10.1093/ajhp/57.10.945. [DOI] [PubMed] [Google Scholar]
- 4.Dorai T., Aggarwal B.B. Role of chemopreventive agents in cancer therapy. Cancer Lett. 2004;215:129–140. doi: 10.1016/j.canlet.2004.07.013. [DOI] [PubMed] [Google Scholar]
- 5.Mahmoud N.N., Carothers A.M., Grunberger D., Bilinski R.T., Churchill M.R., Martucci C., Newmark H.L., Bertagnolli M.M. Plant phenolics decrease intestinal tumors in an animal model of familial adenomatous polyposis. Carcinogenesis. 2000;21:921–927. doi: 10.1093/carcin/21.5.921. [DOI] [PubMed] [Google Scholar]
- 6.Kim S.O., Chun K.S., Kundu J.K., Surh Y.J. Inhibitory effects of [6]-gingerol on PMA-induced COX-2 expression and activation of NFκB and p38 MAPK in mouse skin. Biofactors. 2004;21:27–31. doi: 10.1002/biof.552210107. [DOI] [PubMed] [Google Scholar]
- 7.Murakami A., Tanaka T., Lee J.Y., Surh Y.J., Kim H.W., Kawabata K., Nakamura Y., Jiwajinda S., Ohigashi H. Zerumbone, a sesquiterpene in subtropical ginger, suppresses skin tumor initiation and promotion stages in ICR mice. Inter. J. Cancer. 2004;110:481–490. doi: 10.1002/ijc.20175. [DOI] [PubMed] [Google Scholar]
- 8.Surh Y.J. Anti-tumor promoting potential of selected spice ingredients with antioxidative and anti-inflammatory activities: a short review. Food Chem. Toxicol. 2002;40:1091–1097. doi: 10.1016/S0278-6915(02)00037-6. [DOI] [PubMed] [Google Scholar]
- 9.Surh Y.J., Lee E., Lee J.M. Chemoprotective properties of some pungent ingredients present in red pepper and ginger. Mut. Res. 1998;402:259–267. doi: 10.1016/S0027-5107(97)00305-9. [DOI] [PubMed] [Google Scholar]
- 10.Surh Y.J., Park K.K., Chun K.S., Lee L.J., Lee E., Lee S.S. Anti-tumor- promoting activities of selected pungent phenolic substances present in ginger. J. Environ. Path. Toxicol. Oncol. 1999;18:131–139. [PubMed] [Google Scholar]
- 11.Lee E., Surh Y.J. Induction of apoptosis in HL-60 cells by pungent vanilloids, [6]-gingerol and [6]-paradol. Cancer Lett. 1998;134:163–168. doi: 10.1016/S0304-3835(98)00253-5. [DOI] [PubMed] [Google Scholar]
- 12.Kim E.C., Min J.K., Kim T.Y., Lee S.J., Yang H.O., Han S., Kim Y.M., Kwon Y.G. [6]-Gingerol, a pungent ingredient of ginger, inhibits angiogenesis in vitro and in vivo. Biochem. Biophys. Res. Commun. 2005;335:300–308. doi: 10.1016/j.bbrc.2005.07.076. [DOI] [PubMed] [Google Scholar]
- 13.Chen C.Y., Chen C.H., Kung C.H., Kuo S.H., Kuo S.Y. [6]-Gingerol Induces Ca2+ Mobilization in Madin-Darby Canine Kidney Cells. J. Nat. Prod. 2008;71:137–140. doi: 10.1021/np070279y. [DOI] [PubMed] [Google Scholar]
- 14.Ohizumi Y.,, Sasaki S., Shibusawa K., Ishikawa K., Ikemoto F. Stimulation of sarcoplasmic reticulum Ca(2+)-ATPase by gingerol analogues. Biol. Pharm. Bull. 1996;19:1377–1379. doi: 10.1248/bpb.19.1377. [DOI] [PubMed] [Google Scholar]
- 15.Jemal A., Siegel R., Ward E., Murray T., Xu J., Thun M.J. Cancer statistics. CA Cancer J. Clin. 2007;57:43–66. doi: 10.3322/canjclin.57.1.43. [DOI] [PubMed] [Google Scholar]
- 16.Berridge M.J. Inositol trisphosphate and calcium signaling. Nature. 1993;361:315–325. doi: 10.1038/361315a0. [DOI] [PubMed] [Google Scholar]
- 17.Liantonio A., Giannuzzi V., Picollo A., Babini E., Pusch M., Conte Camerino D. Niflumic acid inhibits chloride conductance of rat skeletal muscle by directly inhibiting the CLC-1 channel and by increasing intracellular calcium. Br. J. Pharmacol. 2007;150:235–247. doi: 10.1038/sj.bjp.0706954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang F., Wen Q., Mergler S., Yang H., Wang Z., Bildin V.N., Reinach P.S. PKC isoform-specific enhancement of capacitative calcium entry in human corneal epithelial cells. Invest. Ophthalmol. Vis. Sci. 2006;47:3989–4000. doi: 10.1167/iovs.06-0253. [DOI] [PubMed] [Google Scholar]
- 19.Berridge M.J. Elementary and global aspects of calcium signaling. J. Physiol. 1997;499:291–306. doi: 10.1113/jphysiol.1997.sp021927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Annunziato L., Amoroso S., Pannaccione A., Cataldi M., Pignataro G., D’Alessio A., Sirabella R., Secondo A., Sibaud L., Di Renzo G.F. Apoptosis induced in neuronal cells by oxidative stress: role played by caspases and intracellular calcium ions. Toxicol. Lett. 2003;139:125–133. doi: 10.1016/S0378-4274(02)00427-7. [DOI] [PubMed] [Google Scholar]
- 21.Ma Y., Hendershot L.M. The role of the unfolded protein response in tumour development: friend or foe? Nat. Rev. Cancer. 2004;4:966–977. doi: 10.1038/nrc1505. [DOI] [PubMed] [Google Scholar]
- 22.Orrenius S., Zhivotovsky B., Nicotera P. Regulation of cell death: the calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003;4:552–565. doi: 10.1038/nrm1150. [DOI] [PubMed] [Google Scholar]
- 23.Soboloff J., Berger S.A. Sustained ER Ca2+ depletion suppresses protein synthesis and induces activation-enhanced cell death in mast cells. J. Biol. Chem. 2002;277:13812–13820. doi: 10.1074/jbc.M112129200. [DOI] [PubMed] [Google Scholar]
- 24.Orrenius S. Oxidative stress studied in intact mammalian cells. Phil. Trans R. Soc. Lond., B Biol. Sci. 1985;311:673–677. doi: 10.1098/rstb.1985.0172. [DOI] [PubMed] [Google Scholar]
- 25.Goldhaber J.I., Qayyum M.S. Oxygen free radicals and excitation-contraction coupling. Antioxi. Redox. Signal. 2000;2:55–64. doi: 10.1089/ars.2000.2.1-55. [DOI] [PubMed] [Google Scholar]
- 26.Chakraborti T., Das S., Mondal M., Roychoudhury S., Chakraborti S. Oxidant, mitochondria and calcium: an overview. Cell. Signal. 1999;11:77–85. doi: 10.1016/S0898-6568(98)00025-4. [DOI] [PubMed] [Google Scholar]
- 27.Putney J.W. A model for receptor-regulated calcium entry. Cell Calcium. 1986;7:1–12. doi: 10.1016/0143-4160(86)90026-6. [DOI] [PubMed] [Google Scholar]
- 28.McFadzean I., Gibson A. The developing relationship between receptor-operated and store-operated calcium channels in smooth muscle. Br. J. Pharmacol. 2002;135:1–13. doi: 10.1038/sj.bjp.0704468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Shoji N., Iwasa A., Takemoto T., Ishida Y. Cardiotonic principles of ginger (Zingiber officinale Roscoe) J. Pharm. Sci. 1982;71:1174–1175. doi: 10.1002/jps.2600711025. [DOI] [PubMed] [Google Scholar]
- 30.Grynkiewicz G., Poenie M., Tsien R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985;260:3440–3450. [PubMed] [Google Scholar]
- 31.Baliga R., Zhang Z., Shah S.V. Role of cytochrome P-450 in hydrogen peroxide-induced cytotoxicity to LLC-PK cells. Kidney Int. 1996;50:1118–1124. doi: 10.1038/ki.1996.418. [DOI] [PubMed] [Google Scholar]
- 32.Andersson D.A., Zygmunt P.M., Movahed P., Andersson T.L., Hogestatt E.D. Effects of inhibitors of small- and intermediate-conductance calcium-activated potassium channels, inwardly-rectifying potassium channels and Na+/K+ ATPase on EDHF relaxations in the rat hepatic artery. Br. J. Pharmacol. 2000;129:1490–1496. doi: 10.1038/sj.bjp.0703226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Das S., Bhattacharyya S., Ghosh S., Majumdar S. TNF-alpha induced altered signaling mechanism in human neutrophil. Mol. Cell. Biochem. 1999;197:97–108. doi: 10.1023/A:1006935114624. [DOI] [PubMed] [Google Scholar]
- 34.Kim J.S., Lee S.I., Park H.W., Yang J.H., Shin T.Y., Kim Y.C., Baek N.I., Kim S.H., Choi S.U., Kwon B.M., Leem K.H., Jung M.Y., Kim D.K. Cytotoxic components from the dried rhizomes of Zingiber officinale Roscoe. Arch Pharm Res. 2008;31:415–418. doi: 10.1007/s12272-001-1172-y. [DOI] [PubMed] [Google Scholar]