
FIGURE 1.
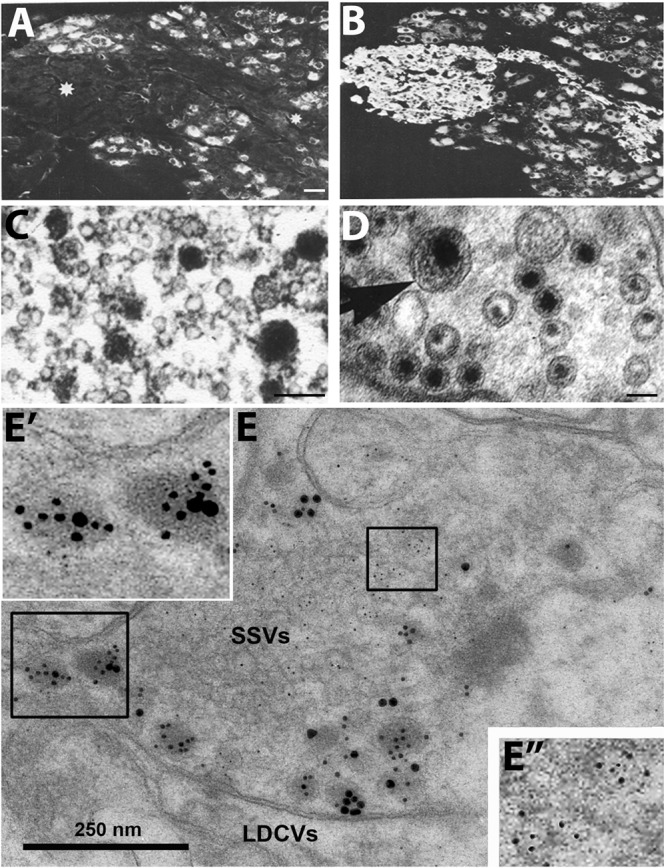
Immunofluorescence micrographs of the guinea-pig inferior mesenteric ganglion (A,B) and electron micrographs from different types of nerve endings (C–E). (A,B) Two adjacent sections incubated with antibodies to somatostatin (A) and the noradrenaline (NA) synthesizing enzyme dopamine ß-hydroxylase (DBH) (B). The majority of the principal ganglion cells are somatostatin-positive, whereas the small intensely fluorescent (SIF) cells (asterisk) lack the peptide. Virtually all ganglion cells and the SIF cells are DBH-positive, i.e., are noradrenergic. (C–E) Examples of transmitter storage in nerve endings based on or immunohistochemistry (C,E) or potassium permanganate fixation (D). (D) In sympathetic nerve endings NA (black precipitate) is stored in both (small) synaptic vesicles and large dense core vesicle (LDCVs) (arrow). Note that content varies between vesicles, both in the synaptic and LDCVs. (C) Substance P, a neuropeptide (black precipitate), in a sensory nerve ending in the monkey dorsal horn, is stored exclusively in LDCVs, all synaptic vesicles are empty. (E) Peptide and glutamate co-storage and coexistence in the dorsal horn of the rat spinal cord based on immunogold immunohistochemistry. Substance P/CGRP is detected with 10/20 nm gold particles and glutamate with 5 nm gold particles. Note that substance P and CGRP can be stored within the same LDCV (left box, magnified in E’). Staining for glutamate is restricted to synaptic vesicles (right box, magnified in E”). The results suggest that glutamate, a small molecule transmitter, is not stored in LDCVs in sensory nerve endings, and release of peptide and amino acid may be separate events. This contrasts NA (see D). Bars: 40 μm, for (A,B); 100 nm for (C,D); 250 nm for (E). (A,B) From Hokfelt et al. (1977a). (C) From DiFiglia et al. (1982), with permission. (D,E) Courtesy of Dr. A. Merighi (cf., Merighi, 2002).