

Abstract
Liver cancer, which is the second leading cause of tumor-associated mortality, is of great concern worldwide due to its resistance to chemotherapeutic drugs. Transcatheter arterial chemoembolization (TACE) has previously been used as a treatment for unresectable liver tumors in China; however, the response to TACE treatment differs between patients. It has been reported that hepatitis B virus (HBV)-as sociated tumors are less sensitive to TACE treatment compared with non-HBV-associated liver cancer. Previous studies have demonstrated that exosomes serve a crucial role in hepatic carcinoma chemoresistance. We therefore hypothesized that HBV may modulate chemosensitivity via exosomes. The aim of the present study was to investigate how exosomes affect chemoresistance by assessing their role in chaperone-mediated autophagy (CMA)-dependent chemoresistance in HBV-associated liver cancer. Iconography data from HBV-positive and HBV-negative patients with hepatic carcinoma receiving TACE treatment were assessed, and it was revealed that the tumor volume was decreased in the patients with non-HBV-associated liver cancer compared with that in the patients with HBV-associated tumors following TACE therapy. Furthermore, it was revealed that exosomes from HBV-infected liver cancer cells were able to downregulate cell apoptosis when treated with oxaliplatin compared with exosomes from normal HepG2 cells. Furthermore, the results demonstrated that HBV-associated exosomes modulate cell death via activating the CMA pathway, and its key molecule, lysosome-associated membrane protein (Lamp2a), was also upregulated. Lamp2a-knockdown was also found to reverse anti-apoptotic effects in liver cancer. Taken together, the results of the present study suggest that chemoresistance in patients with HBV-associated hepatic tumors may be mediated by exosomes, and thus may provide a basis for the development of novel treatment strategies for chemoresistant liver cancer.
Keywords: exosomes, hepatitis B virus-associated liver cancer, chemoresistance, chaperone-mediated autophagy
Introduction
In 2012, liver cancer was reported as a significant cause of tumor-associated mortality worldwide with 746,000 cases and a mortality rate of 9.1% (1). Various treatments for liver cancer are available, including surgery, radiotherapy, systemic chemotherapy and transcatheter arterial chemoembolization (TACE). However, liver cancer is resistant to a number of chemotherapy and radiotherapy regimes (2). TACE, combined arterial embolization with chemotherapy for liver cancer treatment, is typically used as a treatment for advanced liver tumor (3). However, patient responses to TACE therapy are varied. It is known that hepatitis B virus (HBV) is a risk factor for hepatic carcinoma and that it is associated with liver cirrhosis and liver cancer development (4). It has been reported that HBV may affect the efficacy of chemotherapy by mediating gene mutation and intratumor heterogeneity in tumor livers (5). Furthermore, it has been reported that the apoptosis-inducing effects of chemotherapy are different in HBV-associated liver cancer cells compared with that in HBV-negative hepatic tumor cells (6). The aim of the present study was to use clinical data to assess differences in treatment response to TACE in patients with HBV-associated liver cancer and those with non-HBV liver cancer, and to elucidate the possible mechanisms.
Exosomes are membrane-bound microvesicles (50–150-nm) that are released by various cells under normal and pathological conditions (7,8). Exosomes are established as participating in a number of biological functions and contain various cargos, including proteins and RNAs (9). It has been demonstrated that exosomes serve an important role in the formation and progression of tumors, with anti-apoptotic effects in malignant cancer (10–12). For instance, extracellular vesicles released from ovarian cancer cells stimulated by cisplatin treatment may induce invasiveness and bystander cell drug resistance via p38 and c-Jun N-terminal kinase signaling (13). In addition, it has been reported that exosomes are able to induce chemoresistance in liver cancer treated with sorafenib (14).
Chaperone-mediated autophagy (CMA) is a selective form of autophagy in which cytosolic proteins bearing a pentapeptide motif biochemically associated with the KFERQ sequence are recognized by a cytosolic heat shock cognate protein, delivered to the lysosomal membrane and directly translocated across it by a protein complex containing lysosome-associated membrane protein 2a (Lamp2a) (15). CMA is associated with tumor growth, metastasis (16) and resistance to anticancer therapy (17). Lamp2a overexpression in breast tumors increases overall cell survival via the CMA pathway, while Lamp2a inhibition causes glyceraldehyde 3-phosphate dehydrogenase (GAPDH) accumulation, Protein kinase B (AKT1) phosphorylation, reactive oxygen species generation and increased cellular apoptosis in breast cancer cells (18). A previous study revealed that autophagy also serves an essential role in neuroblastoma cells drug resistance and cell survival (19). We therefore speculated that CMA may be able to induce chemoresistance in HBV-associated liver cancer.
In the present study, in order to investigate the underlying mechanisms of different responses to TACE in HBV and non-HBV patients with liver cancer, exosomes derived from HBV-associated liver cancer cells were isolated and the crucial roles of exosomes in chemoresistance were further investigated. The research of the present study would provide novel insight in the underlying mechanisms in liver tumors diagnostics and therapeutics.
Materials and methods
Patient data and samples
Data was collected from patients with hepatic carcinoma who were treated with TACE therapy (oxaliplatin 130 mg/m2) at the Department of Hepatobiliary Surgery between January 2014 and June 2017 in North China University of Science and Technology Affiliated Hospital (Tangshan, China). Inclusion criteria were as follows: Based Specification for Diagnosis and Treatment of Primary Liver Cancer (20); liver function A-B level (Child-Pugh Classification) (21,22); Barcelona Clinical Liver Cancer Staging System (BCLC) B stage (23); patients with no surgical history and other therapy treatments, and patients with no other primary tumor. The exclusion criteria were as follows: Patients suffering from other severe diseases, including heart and renal dysfunction, diffuse liver cancer, coagulation disorders, and patients infected with hepatitis C. Patients were sorted into HBV-associated liver cancer and non-HBV-associated liver cancer groups depending on whether they were positive for hepatitis B surface antigen, hepatitis B e-antigen (HBeAg) and hepatitis B core antibody. In the HBV-associated liver cancer group, there were 3 female and 15 male patients, with a sex ratio (male:female) of 15:3 and a mean age of 54.63±6.21 years (range, 39–68 years). In the non HBV-associated liver cancer group, there were 4 female and 11 male patients, with a sex ratio (male:female) of 11:4 and a mean age of 56.25±7.57 years (range, 38–74 years). Data collection from the patients was approved by the Institutional Review Board of North China University of Science and Technology Affiliated Hospital (Tangshan, Hebei, China). Serum samples were collected from HBV-DNA-positive and healthy individuals at the Clinical Laboratory of North China University of Science and Technology Affiliated Hospital. All specimens were flash-frozen upon collection and stored at −80°C until further use. The present study was approved by the Ethics Review Committee of North China University of Science and Technology Affiliated Hospital (approval no. 17014).
Cell lines, reagents and antibodies
The hepatoblastoma HepG2 cell line (24) (supplied by Cell Resource Center, Shanghai Institute of Life Sciences, Chinese Academy of Sciences, Shanghai, China) was cultured at 37°C in a humidified atmosphere containing 5% CO2 in high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% heat-inactivated FBS (both Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and 100 mg/ml streptomycin (Beijing Solarbio Technology Co., Ltd., Beijing, China). The primary antibody against cleaved caspase-3 (dilution, 1:500; catalog no. PB0183) was obtained from Wuhan Boster Biological Technology, Ltd. (Wuhan, China), the anti-B-cell lymphoma-2 (Bcl-2) antibody (dilution, 1:500; catalog no. WL01556) was obtained from Wanleibio Co., Ltd. (Shanghai, China) and the antibody against GAPDH (dilution, 1:5,000; catalog no. 5174S) was obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA). The antibody against Lamp2a (dilution, 1:1,000; catalog no. ab125068) was obtained from Abcam (Cambridge, MA, USA). The Goat anti-rabbit IgG secondary antibody with the (dilution, 1:4,000; catalog no. BA1039) was obtained from Wuhan Boster Biological Technology, Ltd. (Wuhan, China).
HBV virus infection
Cells were seeded at a concentration of 1×105 cells/dish. At 2 days post-plating, the cells were incubated at 4°C for 2 h, then at 37°C for 6 h in an atmosphere containing 5% CO2. The HepG2 cells were infected with HBV-positive serum (HBV-particles 1×1010 copies/ml) and cocultured with DMEM for 48 h. The HBV-positive serum was then removed, the cells were washed with phosphate-buffered saline (PBS) 8 times and, in order to ensure no residual HBV virus was present in the supernatants, the last washing PBS was restored for polymerase chain reaction analysis to detect HBV virus DNA replication. The cells were subsequently incubated with pure DMEM for 24, 48, 72 and 96 h. Culture supernatants were collected after these different incubation durations and stored at −80°C for later use.
PCR analysis
HBV DNA copies of cell supernatant were quantified by a SLAN-96P Real-time PCR system (Shanghai Hongshi Medical Technology, Co., Ltd., Shanghai, China), according to the Hepatitis B Viral DNA Quantitative Fluorescence Diagnostic Kit (PCR-Fluorescence Probing kit; cat. no., 20153400083; Sansure Biotech Inc., Hunan, China.) instructions. The thermocycling conditions were as follows: 50°C for 2 min, 94°C for 5 min, 45 cycles of 94°C for 15 sec and 57°C for 30 sec, and a final cooling at 25°C for 10 sec.
Isolation and identification of exosomes
HBV-associated exosomes derived from HepG2 cells infected with HBV serum and non-HBV exosomes derived from HepG2 cells incubated with HBV-negative serum were collected from the supernatant. Exosomes were isolated according to Exosome Isolation Reagent protocols (GS™ Exosome Isolation Reagent, cat. no., E1002; Geneseed Biotech, Co., Ltd, Guangzhou, China). Exosomes Vesicles were resuspended in 100–200 µl PBS and stored at −80°C for further use. The biomarkers of exosomes, including cluster of differentiation (CD)9 and CD63, were identified using western blotting and the protein in HepG2 cells was used as a positive control. Exosome pellets were resuspended in PBS and placed onto Formvor carbon-coated electron microscope grids (Electron Microscope Sciences, Hatfield, PA, USA). Following incubation for 5 min at room temperature, exosomes were fixed in 2% paraformaldehyde at room temperature for 4 h, and washed twice with water. The grids were then negatively stained with 10% uranyl acetate for 10 min. The preparations were examined and images were captured using transmission electron microscopy (TEM; JEM-2100; JEOL, Ltd., Tokyo, Japan). Each isolation was verified by nanoparticle tracking analysis using a Nanosight N-300 (Nanosight Ltd., Amesbury, UK) to determine the size and quantity of EVs extracted.
Silencing
HepG2 cells were seeded in 6-well plates with complete medium for 24 h, following which they were transfected with short hairpin RNA (shRNA) targeting Lamp2a (targeted sequence 5′-TCTTATGCATTGGAACTTAATTTGACATCT0-3′; LAMP2 siRNA/shRNA/RNAi lentivirus Human Target A; cat. no., iV012052a; ABM lnc, China) and negative control (NC) shRNA (targeted sequence 5′-GGGTGAACTCACGTCAGAA-3′; Scrambled shRNA GFP lentivirus; cat. no., LVP015-G; ABM lnc.). The concentration of lentivirus was >107 IU/ml. The transfection reagent EndoFectin™-Max (iGeneBio, Guangzhou, China) was used to transfect HepG2 cells with Lamp2a shRNA lentivirus. At 96 h post-transfection, the cells were washed twice and treated with 0.1 µg/µl puromycin for screening. Transfection efficiency was determined immediately using florescence microscopy and the effects of transfection were assessed using western blotting.
Western blotting
HepG2 cells and cells treated with exosomes or PBS were harvested and were lysed for 50 min on ice in 100 ml of lysis buffer (containing phenylmethanesulfonyl fluoride and a phosphatase inhibitor). Lysates were centrifuged at 12,000 × g for 15 min at 4°C. Proteins were quantified by using the bincinchoninic acid protein assay kit (Thermo Fisher Scientific, Inc.), according to the manufacturer's protocols. Equal amounts (60 µg) of cell lysates were loaded and separated by 10% SDS-PAGE, following which proteins were electrotransferred to polyvinylidene difluoride membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The membranes were blocked for 60 min at 37°C with 5% skimmed milk in the 1×TBST (Beijing Solarbio Technology Co., Ltd., Beijing, China) suspension medium and incubated with the aforementioned primary antibodies at 4°C overnight. After three washes with TBST, the membranes were incubated with secondary antibodies for 60 min at room temperature and washed again. Proteins were detected using an enhanced chemiluminescence system (Pierce; Thermo Fisher Scientific, Inc.). Each experiment was repeated three times and similar results were obtained.
Flow cytometry
Cell samples were analyzed using flow cytometry analysis. Cells were stained with Annexin-V and propidium iodide reagents (Annexin V-FITC/PI Apoptosis Detection kit; catalog no. BB-4101-1; Bestbio, Co., Shanghai, China) to assess apoptosis. Data were analyzed using a FACSCalibur flow cytometer and BD CellQuest Pro software 5.1 (BD Biosciences, Franklin Lakes, NJ, USA). The analysis was performed three times.
Statistical analysis
Graphs were created using Image Lab system (v4.1; Bio-Rad Laboratories, Inc.). Student's t-test or one-way analysis of variance with Scheffe's F post hoc test were performed using SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA). Data were presented as mean ± standard deviation. P<0.05 was considered to indicate a statistically significant difference.
Results
Tumor volume reduction is lower in HBV-associated hepatic carcinoma patients compared with that in non-HBV-associated liver cancer patients following TACE treatment
To determine the effects of HBV on the therapeutic efficacy of TACE, pre- and post-treatment tumor volume data were collected for HBV-positive and -negative liver cancer patients treated with TACE. In addition, a comparison of general patient characteristics was performed, as shown in Table I. The data results revealed that the reduction in tumor volume following TACE was significantly smaller in the HBV-associated liver cancer group compared with the HBV-negative liver cancer group (Fig. 1). These results indicated that HBV infection was associated with a reduced response to TACE therapy.
Table I.
Comparison of patient characteristics between the experimental and control groups.
| Clinical features | HBV-associated liver cancer | Non-HBV-associated liver cancer |
|---|---|---|
| Sex (male/female) | 15/3 | 11/4 |
| Mean age, years (mean ± standard deviation) | 54.63±6.21 | 56.25±7.57 |
| Liver function (Child-Pugh classification) | A-B | A-B |
| BCLC staging system (23) | B | B |
BCLC, Barcelona Clinic Liver Cancer staging system; HBV, hepatitis B virus.
Figure 1.
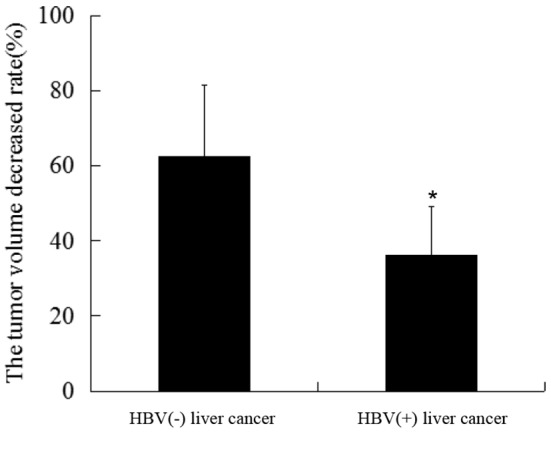
Tumor volume reduction in patients with HBV-associated hepatic carcinoma and non-HBV-associated liver cancer following transcatheter arterial chemoembolization. *P<0.05 vs. HBV(−) liver cancer. HBV, hepatitis B virus.
Exosomes derived from HBV-associated HepG2 cells modulate chemoresistance in liver cancer cells
A cell model infected with HBV serum was established as previously described (25). PCR analysis was used to confirm that the last PBS wash was negative for HBV-DNA. Therefore, no residual HBV virus existed in the washing supernatants. As indicated in Fig. 2A, the copies of HBV DNA in the 96 h duration was the highest compared with 12, 24 and 48 h durations. Cell supernatant containing pure DMEM for 96 h were selected for use in further experiments.
Figure 2.

Exosomes derived from HBV-positive liver cancer cells promote cell chemoresistance. (A) HBV-DNA contents in culture supernatant reached a peaked at 96 h. Exosomes were isolated according to manufacturer's instructions (47). HepG2 cells were treated with HBV-positive serum (HBV-DNA 1×1010 copies/ml) and HBV-DNA copy of the cell supernatant containing pure DMEM was detected at different times following removal of serum; experiments demonstrated that HBV-DNA copies reached a peaked (16.89±5.02×103 copies/ml) at 96 h comparing with HBV-DNA copies in 12, 24, 48 h, which provided the basis for subsequent experiments. (B) Exosome markers CD63 and CD9 were assessed by western blotting. GAPDH was used as an internal reference. (C) Exosomes were characterized as round vesicles when examined using transmission electron microscopy (magnification, ×40,000). (D) Detection of cell apoptosis on treatment with 1×1010 particles of HBV-associated exosomes in the experimental group and an equal amount of particles of non-HBV-associated exosomes in the negative group. (E) Flow cytometry analysis of cell apoptosis. (F) Analysis of size distribution and exosome concentration in purified exosomes using Nanosight technology. No statistically significant differences were indicated between HBV-associated and non-HBV-associated liver cancer cells concerning exosomes concentration. #P>0.05 vs. exosomes from non-HBV-associated liver cancer cells. HBV, hepatitis B virus; Bcl-2, B-cell lymphoma; CD, cluster of differentiation; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Exo, exosome; Oxa, oxaliplatin; UR, upper right; LR, lower right.
To confirm that the pellets were exosomes, their characteristics were determined via various methods, including western blotting, TEM and Nanosight tracking analysis. Exosome markers CD63 and CD9 were identified using western blotting (Fig. 2B). Exosomes were also confirmed to have round vesicular morphology, as observed in the pellets (Fig. 2C). It has been reported that exosomes released from liver cancer cells induce cell resistance to sorafenib in vivo and in vitro (14). However, the effects of exosomes derived from HBV-associated hepatic tumors on liver cell chemoresistance remain unknown. In the present study, it was demonstrated that different apoptotic effects were achieved when cells were treated with equivalent HBV-positive or -negative exosomes (1×1010 particles). Cleaved caspase-3 expression was decreased and Bcl-2 expression was markedly increased in HepG2 cells treated with HBV-associated exosomes compared with that in the negative and blank control groups (Fig. 2D and E). Exosome concentration and distribution were analyzed using Nanosight tracking analysis following their isolation from the supernatant. The results revealed that there was no significant difference in exosome concentration between cells treated with HBV-positive and HBV-negative serum (Fig. 2F). As such, the possibility that exosomes released from liver cancer cells infected with HBV induce chemoresistance due to an increase in exosome secretion was eliminated. It has previously been demonstrated in certain tumors that specific exosomes containing RNAs can be transferred to specific target cells, in which shuttled RNA induces functional chemoresistance (12,13). We therefore concluded that HBV-associated exosomes significantly downregulate apoptosis in liver cancer cells by affecting the CMA pathway.
HBV-associated exosomes downregulate chemosensitivity in a concentration-dependent manner
To further identify the role of HBV-associated exosomes in liver tumor chemoresistance, apoptosis was assessed in cells transfected with HBV-associated exosomes at different concentrations (4×109, 8×109 and 1.6×1010 particles/ml). The results revealed that apoptosis was negatively associated with the concentration of HBV-associated exosomes (Fig. 3A and B). The data therefore confirmed that exosomes released from HBV-associated liver cancer cells could downregulate cell sensitivity to oxaliplatin therapy in a concentration-dependent manner. Previous studies have reported that CMA serves a role in tumor anti-apoptosis (17). Based on this, the following experiments were performed.
Figure 3.

Cell apoptosis was decreased by HBV-associated exosomes in a concentration-dependent manner. (A) Cells were treated with 1×1010, 2×1010 or 4×1010 HBV-associated exosomes, and the expression of cleaved caspase-3 and Bcl-2 was assessed by western blotting. (B) Flow cytometry results indicated that cell apoptosis was negatively associated with the concentration of HBV-associated exosomes. HBV, hepatitis B virus; Bcl-2, B-cell lymphoma; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Exo, exosome; Oxa, oxaliplatin; UR, upper right; LR, lower right; Ctr, control; PBS, phosphate-buffered saline.
CMA activation decreases chemosensitivity in cells treated with HBV-associated exosomes
CMA serves important roles in tumor progression and inhibiting apoptosis (16,17). Autophagy has previously been reported to be involved in cancer drug resistance (19). The aim of the present study was to investigate whether the interaction between exosomes and CMA influenced HBV-associated liver cancer chemoresistance. The expression of Lamp2a, a key molecule in the CMA pathway, was therefore investigated. Lamp2a expression was negatively associated with cell apoptosis. In HepG2 cells treated with HBV-associated exosomes and oxaliplatin, Lamp2a expression was upregulated in a concentration-dependent manner (Fig. 4A and B). These results revealed that HBV-associated exosomes decreased cell chemosensitivity by activating the CMA pathway. Furthermore, in order to investigate whether the apoptotic rate of cells treated with HBV-associated exosomes was associated with CMA activation, Lamp2a was knocked down in the present study. There was no indication of Lamp2a protein expression on sh-Lamp2a HepG2 cells. (Fig. 4C and D). When cells were treated with HBV-associated exosomes, apoptosis was increased in HepG2-Lamp2a shRNA cells compared with in HepG2-control shRNA cells (Fig. 4E). Therefore, HBV-associated exosomes could influence cell viability via regulating the CMA pathway in liver cancer chemotherapy.
Figure 4.

HBV-associated exosomes modulate drug resistance via CMA pathway activation. (A) Western blotting revealed that Lamp2a was highly expressed following treatment with oxaliplatin in the HBV-associated exosome group compared with in the non-HBV exosome and blank control groups, and (B) that Lamp2a was upregulated by HBV-associated exosomes in a concentration-dependent manner. (C) Transfection efficiency of shRNA as assessed using fluorescence microscopy. (D) Interference efficiency of shRNA in HepG2 cells as assessed by western blotting. (E) The anti-apoptotic effects of HBV-associated exosomes in HepG2 cells were reversed by Lamp2a silencing. Lamp2a, lysosome-associated membrane protein; HBV, hepatitis B virus; oxa, oxaliplatin; CMA, chaperone-mediated autophagy; shRNA, short hairpin RNA; UR, upper right; LR, lower right; Ctr, control; PBS, phosphate-buffered saline; NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Discussion
The present study demonstrated the effect of exosomes derived from HBV-associated HepG2 cells on liver cancer chemoresistance. Hepatoblastoma cell HepG2 cells has been reported could generate resistance to chemotherapeutic drugs in vivo and in vitro (26–28). Previous studies reported that exosomes derived from hepatic carcinoma cells may generate resistance to sorafenib in mice and induce sorafenib resistance via the HGF/c-Met/Akt pathway (14). It has been reported that HBV infection increases the risk of hepatic carcinoma development and progression (29,30). Furthermore, HBV-associated liver cancer cells and non-HBV-associated liver cancer cells respond differently to chemotherapy (6). The focus of the current study was the effects of exosomes derived from HepG2 cells infected with HBV-positive serum on liver cancer chemoresistance. It has been reported that the HBV virus affects tumor cell growth and associated microRNA (miR) production (31), and it has been revealed that HBeAg-induced miR-106b expression contributes to the pathogenesis of HBV-associated liver cancer by downregulating the retinoblastoma gene (32). Furthermore, various long non-coding RNAs serve functional roles in HBV-associated hepatic carcinoma by regulating biological processes (33,34).
Exosomes are 50- to 150-nm extracellular vesicles released from cells that deliver cell-to-cell communications in diverse conditions (7,35). It has been indicated that miR-21 in exosomes derived from neuroblastoma (NBL) is transferred to human monocytes, while miR-155 in exosomes released from human monocytes is transferred to NBL cells, and that their interaction may mediate cisplatin resistance (36). miR-155 released by exosomes derived from cancer stem cells mediates chemoresistance and migration in breast cancer cells (37). Indeed, in various tumors, the cargo transferred by exosomes clearly serves a role in chemoresistance (38). CMA has been reported to serve a role in the development of various cancer types, including breast cancer (16,39) and gastric cancer (40). CMA activation has been assessed by measuring the expression of Lamp2a, which is required for the growth of breast tumors (18). We previously demonstrated that CMA pathway activation serves an important role in liver cancer chemoresistance and radioresistance (41). However, to the best of our knowledge, no evidence exists regarding the combined effect of exosomes and the CMA pathway on liver cancer chemoresistance. In the present study, in order to specify the effects of CMA activation on liver cancer chemoresistance, the expression of Lamp2a was investigated using western blotting following incubation with HBV-associated exosomes and oxaliplatin. The results revealed that Lamp2a was highly expressed in cells treated with HBV-associated exosomes, and that it was negatively associated with apoptosis. These results indicate that exosomes derived from HBV-associated liver cancer cells promote chemoresistance by modulating the CMA pathway. These findings may have clinical relevance for HBV-associated liver cancer resistance to TACE treatment.
However, the mechanisms by which HBV-associated exosomes regulate the CMA pathway to induce chemoresistance in liver cancer cells were not directly addressed in the current study. Exosomes serve a role in a range of biological processes and it has been revealed that miR-1246 carried by exosomes may induce breast cancer progression and chemoresistance via targeting of cyclin-G2 (42). More notably, it has been reported that HBV-encoded X protein induces miR-21 expression in hepatic tumor cells, and upregulates levels of exosomal miR-21 in hepatic carcinoma cells (43,44). miR-21 upregulation may repress the tumor-suppressor function of programmed cell death protein 4, leading to the proliferation of hepatic carcinoma cells (45). In addition, miR-21 has been indicated to modulate systemic therapy resistance via autophagy in breast cancer cells (46). It was therefore assumed that exosomal miR-21 may modulate HBV-associated liver cancer chemoresistance via CMA activation. Further studies are required to elucidate the exact role of exosomes released from HBV-associated HepG2 cells on the efficacy of chemotherapy in liver cancer, and to identify novel treatment targets for liver cancer patients with HBV-infection.
In summary, the results of the present study revealed that tumor sensitivity to TACE differed greatly between HBV and non-HBV liver cancer patients. To the best of our knowledge, the present study is the first to investigate the interaction between exosomes and CMA pathway activation in liver cancer chemoresistance, and may provide a basis for targeting exosomes to increase chemosensitivity in patients with liver cancer and HBV infection.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural Science Foundation of Hebei Province (grant no. H2016209007), the Project for Scientific Research Program of Health and Family Planning Commission of Hebei (grant no. 20180754), the State Administration of Traditional Chinese Medicine of Hebei (grant no. 2018182) and the Innovation Project for Postgraduate of North China University of Science and Technology, Hebei Province, China (grant no. 2018S39).
Availability of data and material
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Authors' contributions
DXL and PPL participated in the literature search, study design, data collection, data analysis and data interpretation, and wrote the manuscript. JPG, LLL and BG performed data collection and analysis, and provided critical revision. HBJ conceived the study and participated in its design and coordination. JHW and JMC participated in study design and provided the critical revision.
Ethics approval and consent to participate
The present study was approved by the Institutional Review Board of North China University of Science and Technology (approval no. 17014). Written consent for enrollment was provided by all patients.
Patient consent for publication
Not applicable.
Competing interests
All authors declare that there they have no competing interests.
References
- 1.Wallace MC, Preen D, Jeffrey GP, Adams LA. The evolving epidemiology of hepatocellular carcinoma: A global perspective. Expert Rev Gastroenterol Hepatol. 2015;9:765–779. doi: 10.1586/17474124.2015.1028363. [DOI] [PubMed] [Google Scholar]
- 2.Wahid B, Ali A, Rafique S, Idrees M. New insights into the epigenetics of hepatocellular carcinoma. Biomed Res Int. 2017;2017:1609575. doi: 10.1155/2017/1609575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang Y, Ma L, Sheng S, Yuan Z, Zheng J, Li W. Combination therapy of TACE and CT-guided partial hepatic segment ablation for liver cancer. Minim Invasive Ther Allied Technol. 2018:1–10. doi: 10.1080/13645706.2018.1462833. (Epub ahead of print) [DOI] [PubMed] [Google Scholar]
- 4.Feng GX, Li J, Yang Z, Zhang SQ, Liu YX, Zhang WY, Ye LH, Zhang XD. Hepatitis B virus X protein promotes the development of liver fibrosis and hepatoma through downregulation miR-30e targeting P4HA2 mRNA. Oncogene. 2017;36:6895–6905. doi: 10.1038/onc.2017.291. [DOI] [PubMed] [Google Scholar]
- 5.Castelli G, Pelosi E, Testa U. Liver cancer: Molecular characterization, clonal evolution and cancer stem cells. Cancers (Basel) 2017;9(pii):E127. doi: 10.3390/cancers9090127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li XY, Wen JY, Jia CC, Wang TT, Li X, Dong M, Lin QU, Chen ZH, Ma XK, Wei LI, et al. MicroRNA-34a-5p enhances sensitivity to chemotherapy by targeting AXL in hepatocellular carcinoma MHCC-97L cells. Oncol Lett. 2015;10:2691–2698. doi: 10.3892/ol.2015.3654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Raposo G, Stoorvogel W. Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol. 2013;200:373–383. doi: 10.1083/jcb.201211138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pugholm LH, Revenfeld AL, Søndergaard EK, Jørgensen MM. Antibody-based assays for phenotyping of extracellular vesicles. Biomed Res Int. 2015;2015:524817. doi: 10.1155/2015/524817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Geis-Asteggiante L, Belew AT, Clements VK, Edwards NJ, Ostrand-Rosenberg S, El-Sayed NM, Fenselau C. Differential content of proteins, mRNAs, and miRNAs suggests that MDSC and their exosomes may mediate distinct immune suppressive functions. J Proteome Res. 2018;17:486–498. doi: 10.1021/acs.jproteome.7b00646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Oushy S, Hellwinkel JE, Wang M, Nguyen GJ, Gunaydin D, Harland TA, Anchordoquy TJ, Graner MW. Glioblastoma multiforme-derived extracellular vesicles drive normal astrocytes towards a tumour-enhancing phenotype. Philos Trans R Soc Lond B Biol Sci. 2018;373(pii):20160477. doi: 10.1098/rstb.2016.0477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kawakubo-Yasukochi T, Morioka M, Hazekawa M, Yasukochi A, Nishinakagawa T, Ono K, Kawano S, Nakamura S, Nakashima M. miR-200c-3p spreads invasive capacity in human oral squamous cell carcinoma microenvironment. Mol Carcinog. 2018;57:295–302. doi: 10.1002/mc.22744. [DOI] [PubMed] [Google Scholar]
- 12.Min QH, Wang XZ, Zhang J, Chen QG, Li SQ, Liu XQ, Li J, Liu J, Yang WM, Jiang YH, et al. Exosomes derived from imatinib-resistant chronic myeloid leukemia cells mediate a horizontal transfer of drug-resistant trait by delivering miR-365. Exp Cell Res. 2018;362:386–393. doi: 10.1016/j.yexcr.2017.12.001. [DOI] [PubMed] [Google Scholar]
- 13.Samuel P, Mulcahy LA, Furlong F, McCarthy HO, Brooks SA, Fabbri M, Pink RC, Carter DRF. Cisplatin induces the release of extracellular vesicles from ovarian cancer cells that can induce invasiveness and drug resistance in bystander cells. Philos Trans R Soc Lond B Biol Sci. 2018;373(pii):20170065. doi: 10.1098/rstb.2017.0065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Qu Z, Wu J, Wu J, Luo D, Jiang C, Ding Y. Exosomes derived from HCC cells induce sorafenib resistance in hepatocellular carcinoma both in vivo and in vitro. J Exp Clin Cancer Res. 2016;35:159. doi: 10.1186/s13046-016-0430-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Catarino S, Pereira P, Girão H. Molecular control of chaperone-mediated autophagy. Essays Biochem. 2017;61:663–674. doi: 10.1042/EBC20170057. [DOI] [PubMed] [Google Scholar]
- 16.Han Q, Deng Y, Chen S, Chen R, Yng M, Zhang Z, Sun X, Wang W, He Y, Wang F, et al. Downregulation of ATG5-dependent macroautophagy by chaperone-mediated autophagy promotes breast cancer cell metastasis. Sci Rep. 2017;7:4759. doi: 10.1038/s41598-017-04994-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dewaele M, Martinet W, Rubio N, Verfaillie T, de Witte PA, Piette J, Agostinis P. Autophagy pathways activated in response to PDT contribute to cell resistance against ROS damage. J Cell Mol Med. 2011;15:1402–1414. doi: 10.1111/j.1582-4934.2010.01118.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Saha T. LAMP2A overexpression in breast tumors promotes cancer cell survival via chaperone-mediated autophagy. Autophagy. 2012;8:1643–1656. doi: 10.4161/auto.21654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Oehme I, Linke JP, Böck BC, Milde T, Lodrini M, Hartenstein B, Wiegand I, Eckert C, Roth W, Kool M, et al. Histone deacetylase 10 promotes autophagy-mediated cell survival. Proc Natl Acad Sci USA. 2013;110:E2592–E2601. doi: 10.1073/pnas.1300113110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Qin S Primary Liver Cancer Diagnosis and Treatment Expert Panel of the Chinese Ministry of Health, corp-author. Guidelines on the diagnosis and treatment of primary liver cancer (2011 edition) Chin Clin Oncol. 2012;1:10. doi: 10.3978/j.issn.2304-3865.2012.07.01. [DOI] [PubMed] [Google Scholar]
- 21.Lee TY, Lin CC, Chen CY, Wang TE, Lo GH, Chang CS, Chao Y Combination of transcatheter arterial chemoembolization and interrupted dosing sorafenib improves patient survival in early-intermediate stage hepatocellular carcinoma, corp-author. A post hoc analysis of the START trial. Medicine (Baltimore) 2017;96:e7655. doi: 10.1097/MD.0000000000007655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yamakado K, Miyayama S, Hirota S, Mizunuma K, Nakamura K, Inaba Y, Maeda H, Matsuo K, Nishida N, Aramaki T, et al. Subgrouping of intermediate-stage (BCLC stage B) hepatocellular carcinoma based on tumor number and size and Child-Pugh grade correlated with prognosis after transarterial chemoembolization. Jpn J Radiol. 2014;32:260–265. doi: 10.1007/s11604-014-0358-1. [DOI] [PubMed] [Google Scholar]
- 23.European Association For The Study Of The Liver1; European Organisation For Research And Treatment Of Cancer. EASL-EORTC clinical practice guidelines: Management of hepatocellular carcinoma. J Hepatol. 2012;56:908–943. doi: 10.1016/j.jhep.2011.12.001. [DOI] [PubMed] [Google Scholar]
- 24.López-Terrada D, Cheung SW, Finegold MJ, Knowles BB. Hep G2 is a hepatoblastoma-derived cell line. Hum Pathol. 2009;40:1512–1515. doi: 10.1016/j.humpath.2009.07.003. [DOI] [PubMed] [Google Scholar]
- 25.Paran N, Geiger B, Shaul Y. HBV infection of cell culture: Evidence for multivalent and cooperative attachment. EMBO J. 2001;20:4443–4453. doi: 10.1093/emboj/20.16.4443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Luo D, Cheng SC, Xie H, Xie Y. Effects of Bcl-2 and Bcl-XL protein levels on chemoresistance of hepatoblastoma HepG2 cell line. Biochem Cell Biol. 2000;78:119–126. doi: 10.1139/bcb-78-2-119. [DOI] [PubMed] [Google Scholar]
- 27.Hsiao CC, Chen PH, Cheng CI, Tsai MS, Chang CY, Lu SC, Hsieh MC, Lin YC, Lee PH, Kao YH. Toll-like receptor-4 is a target for suppression of proliferation and chemoresistance in HepG2 hepatoblastoma cells. Cancer Lett. 2015;368:144–152. doi: 10.1016/j.canlet.2015.08.004. [DOI] [PubMed] [Google Scholar]
- 28.Vander Borght S, van Pelt J, van Malenstein H, Cassiman D, Renard M, Verslype C, Libbrecht L, Roskams TA. Up-regulation of breast cancer resistance protein expression in hepatoblastoma following chemotherapy: A study in patients and in vitro. Hepatol Res. 2008;38:1112–1121. doi: 10.1111/j.1872-034X.2008.00381.x. [DOI] [PubMed] [Google Scholar]
- 29.Rapti I, Hadziyannis S. Risk for hepatocellular carcinoma in the course of chronic hepatitis B virus infection and the protective effect of therapy with nucleos(t)ide analogues. World J Hepatol. 2015;7:1064–1073. doi: 10.4254/wjh.v7.i8.1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Shirvani-Dastgerdi E, Winer BY, Celià-Terrassa T, Kang Y, Tabernero D, Yagmur E, Rodríguez-Frías F, Gregori J, Luedde T, Trautwein C, et al. Selection of the highly replicative and partially multidrug resistant rtS78T HBV polymerase mutation during TDF-ETV combination therapy. J Hepatol. 2017;67:246–254. doi: 10.1016/j.jhep.2017.03.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lamontagne J, Steel LF, Bouchard MJ. Hepatitis B virus and microRNAs: Complex interactions affecting hepatitis B virus replication and hepatitis B virus-associated diseasses. World J Gastroenterol. 2015;21:7375–7399. doi: 10.3748/wjg.v21.i24.7375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Samal J, Kandpal M, Vivekanandan P. HBeAg-induced miR-106b promotes cell growth by targeting the retinoblastoma gene. Sci Rep. 2017;7:14371. doi: 10.1038/s41598-017-14652-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fan H, Zhang Q, Zhao X, Lv P, Liu M, Tang H. Transcriptomic profiling of long non-coding RNAs in hepatitis B virus-related hepatocellular carcinoma. Oncotarget. 2017;8:65421–65434. doi: 10.18632/oncotarget.18897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Moyo B, Nicholson SA, Arbuthnot PB. The role of long non-coding RNAs in hepatitis B virus-related hepatocellular carcinoma. Virus Res. 2016;212:103–113. doi: 10.1016/j.virusres.2015.07.025. [DOI] [PubMed] [Google Scholar]
- 35.Bayraktar R, Van Roosbroeck K, Calin GA. Cell to cell communication: microRNAs as hormones. Mol Oncol. 2017;11:1673–1686. doi: 10.1002/1878-0261.12144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Challagundla KB, Wise PM, Neviani P, Chava H, Murtadha M, Xu T, Kennedy R, Ivan C, Zhang X, Vannini I, et al. Exosome-mediated transfer of microRNAs within the tumor microenvironment and neuroblastoma resistance to chemotherapy. J Natl Cancer Inst. 2015;107(pii):djv135. doi: 10.1093/jnci/djv135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Santos JC, Lima NDS, Sarian LO, Matheu A, Ribeiro ML, Derchain SFM. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci Rep. 2018;8:829. doi: 10.1038/s41598-018-19339-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Boelens MC, Wu TJ, Nabet BY, Xu B, Qiu Y, Yoon T, Azzam DJ, Twyman-Saint Victor C, Wiemann BZ, Ishwaran H, et al. Exosome transfer from stromal to breast cancer cells regulates therapy resistance pathways. Cell. 2014;159:499–513. doi: 10.1016/j.cell.2014.09.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li L, Fang R, Liu B, Shi H, Wang Y, Zhang W, Zhang X, Ye L. Deacetylation of tumor-suppressor MST1 in Hippo pathway induces its degradation through HBXIP-elevated HDAC6 in promotion of breast cancer growth. Oncogene. 2016;35:4048–4057. doi: 10.1038/onc.2015.476. [DOI] [PubMed] [Google Scholar]
- 40.Melo SA, Luecke LB, Kahlert C, Fernandez AF, Gammon ST, Kaye J, LeBleu VS, Mittendorf EA, Weitz J5, Rahbari N, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature. 2015;523:177–182. doi: 10.1038/nature14581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wu JH, Guo JP, Shi J, Wang H, Li LL, Guo B, Liu DX, Cao Q, Yuan ZY. CMA down-regulates p53 expression through degradation of HMGB1 protein to inhibit irradiation-triggered apoptosis in hepatocellular carcinoma. World J Gastroenterol. 2017;23:2308–2317. doi: 10.3748/wjg.v23.i13.2308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Li XJ, Ren ZJ, Tang JH, Yu Q. Exosomal microRNA MiR-1246 promotes cell proliferation, invasion and drug resistance by targeting CCNG2 in breast cancer. Cell Physiol Biochem. 2017;44:1741–1748. doi: 10.1159/000485780. [DOI] [PubMed] [Google Scholar]
- 43.Qiu X, Dong S, Qiao F, Lu S, Song Y, Lao Y, Li Y, Zeng T, Hu J, Zhang L, et al. HBx-mediated miR-21 upregulation represses tumor-suppressor function of PDCD4 in hepatocellular carcinoma. Oncogene. 2013;32:3296–3305. doi: 10.1038/onc.2013.150. [DOI] [PubMed] [Google Scholar]
- 44.Kouwaki M, Fukushima Y, Daito T, Sanada T, Yamamoto N, Mifsud EJ, Leong CR, Tsukiyama-Kohara K, Kohara M, Matsumoto M, et al. Extracellular vesicles including exosomes regulate innate immune responses to hepatitis B virus infection. Front Immunol. 2016;7:335. doi: 10.3389/fimmu.2016.00335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Damania P, Sen B, Dar SB, Kumar S, Kumari A, Gupta E, Sarin SK, Venugopal SK. Hepatitis B virus induces cell proliferation via HBx-induced microRNA-21 in hepatocellular carcinoma by targeting programmed cell death protein4 (PDCD4) and phosphatase and tensin homologue (PTEN) PLoS One. 2014;9:e91745. doi: 10.1371/journal.pone.0091745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Campos-Parra AD, Mitznahuat1 GC, Pedroza-Torres A, Romo RV, Reyes FIP, López-Urrutia E, Pérez-Plasencia C. Micro-RNAs as potential predictors of response to breast cancer systemic therapy: Future clinical implications. Int J Mol Sci. 2017;18(pii):E1182. doi: 10.3390/ijms18061182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Malla B, Aebersold DM, Dal Pra A. Protocol for serum exosomal miRNAs analysis in prostate cancer patients treated with radiotherapy. J Transl Med. 2018;16:223. doi: 10.1186/s12967-018-1592-6. [DOI] [PMC free article] [PubMed] [Google Scholar]