

Abstract
Climate change models predict frequent and intense droughts in the world. Development of drought-tolerant species and cultivars is necessary to cope with such changes. Forage grass species are affected, especially in the Mediterranean region. The aim of the present study was to investigate the diversity for drought survival, summer dormancy, and productivity within a cocksfoot population.
The study was conducted in Morocco, under field conditions from 2011 to 2013. 283 genotypes of cocksfoot and parents were tested, characterized for dry matter yield, heading date, plant height, senescence, summer dormancy, and drought survival. Results exhibited a large variability between traits. 79% of the population had survived after severe drought summer while 57% yielded more than both parents. Also, 63% of the progeny had an intermediate score of summer dormancy estimated by senescence score. Large variability was also noticed for heading date and plant height. Several accessions combined a high yield and persistence under severe summer drought. Which explain the significant correlation (r = 0.18, P < 0.005) founded between total dry matter accumulated in 2013 and plant survival. Accordingly, our results showed that we can rise persistent and resilient genotypes among population with a good level of biomass.
Keywords: Cocksfoot, Drought resistance, Hybrids, Plants survival, Summer dormancy
Abbreviations: ADM, autumn recovery dry matter; ANOVA, one-way analysis of variance procedure; DS, drought survival; HD, heading date; PCA, Principal Component Analyses; PH, plant height; SD, summer dormancy; SDI, summer dormancy index; Senesc, senescence; SpB, spring biomass; S/Sp, Norton index; TDM, total dry matter
1. Introduction
Under global climate changes, precipitation is anticipated to vary greatly with increasing intra- and inter-annual variability (IPCC, 2007). These changes have influences on perennial grass growth and survival (Howden et al., 2008, Cullen et al., 2009). A positive correlation exists between plant productivity and annual precipitation in grassland ecosystems (Knapp and Smith, 2001, Dukes et al., 2005, Chimner et al., 2010). Under Mediterranean climate, the persistence of perennial herbaceous plants is mainly determined by plant survival over consecutive summer droughts (Volaire, 2008, Lelièvre and Volaire, 2009). Perennial grasses, such as cocksfoot and tall fescue, are required to persist over many years. These grasses are useful in many cases to improve the sustainability of agricultural systems (Bell et al., 2007). Moreover, perennial grasslands are expected to produce for many years and their sustainability is combined with yield stability and long-term resilience (Volaire et al., 2014). Recent studies approved that both cocksfoot and tall fescue are adapted to Mediterranean climates (Annicchiarico et al., 2011, Pecetti et al., 2011). These forage species are characterized by their winter production and tolerance (Ogle et al., 2011). Moroccan summer drought tolerance (Lelièvre and Volaire, 2009). Also, these species have a valuable trait to survive drought, which may be as important as productivity (Bolger et al., 2007).
Cocksfoot Dactylis glomerata L. is an autotetraploid perennial grass species, characterized by good productivity in pure or mixed crops and drought cocksfoot presents complete summer dormancy (Norton et al., 2006b, Norton et al., 2008), which ensures its superior survival under extreme drought. Moroccan populations were found more adapted to severe water deficit conditions since they maintained a longer and a higher photosynthetic rate even in the most severe drought treatments (Salis et al., 2006). Summer dormancy is defined as suppression of growth in the summer despite irrigation (Norton et al., 2009). Dormancy is more potent than drought survival traits while it improves persistence of pasture plants over severe drought (Norton, 2011). It confers to genotypes of some grass species the endogenous ability to cease aerial growth and senesce irrespective of the water supply in summer (Volaire and Norton, 2006). Besides, dormancy is associated with superior survival and autumn regrowth after severe and repeated summer drought (Volaire and Norton, 2006, Shaimi et al., 2009b) as in tall Fescue (Norton et al., 2006b) and Phalaris (Culvenor and Boschma, 2005). Moreover, the persistence of perennial herbaceous plants is mainly determined by plant survival over repeated summer droughts (Volaire, 2008). In fact, it is the main indicator of drought resistance for perennial grasses (Lelièvre et al., 2011). Drought survival is defined as valuable plant adaptation during part of the plant cycle. Which may enhance long term persistence under increasing drought (Lelièvre et al., 2011). Additionally, plant survival through summer droughts is one of the most important adaptive responses. It determines persistence, long-term production and water use efficiency during the next seasons (Kallida et al., 2010). Therefore the most important strategy is not conservation of production during drought, but the capacity to survive and recover rapidly after autumn rains (Kemp and Culvenor, 1990). When the photosynthetic leaf area is lost, carbohydrate reserves provide resources for the survival of cocksfoot (Volaire, 1991, Volaire, 1994, Volaire, 1995). Besides, plant survival relies on some physiological process such as dehydration tolerance in surviving tissues and the ability of roots to extract water at low soil water potentials (Volaire and Lelievre, 2001). It is associated to how long the surviving tissues can maintain cell integrity at a given moisture content, rather than on the actual minimum threshold of dehydration reached by the tissues (Volaire, 2002). Furthermore, survival is combined to summer dormancy in semi-arid environment (Volaire and Norton, 2006). Indeed, dormant genotypes recover after the drought for a longer time at lower soil water contents (Volaire et al., 2009). Other research confirmed that superior survival after severe and repeated summer droughts is generally associated with summer dormancy (Norton et al., 2006a, Norton et al., 2012).
The main objective of this study is to determine and analyze the persistence and resilience among genotypes from mapping population of cocksfoot with regard to their biomass productivity.
2. Materials and methods
2.1. Plant material and growth conditions
Two cultivars of cocksfoot and their hybrids were compared under field trial. The cultivar ‘Kasbah’ Dactylis glomerata ssp. hispanica exhibits a complete summer dormancy under irrigation very highly drought resistant (Norton et al., 2006a, Norton et al., 2006b, Norton et al., 2008), less productive than Medly cv and more tolerant to severe drought (Annicchiarico et al., 2011, Lelièvre et al., 2011), originated from southern of Morocco and was bred in Australia. The ‘Medly’ cultivar Dactylis glomerata ssp. glomerata is an early flowering variety, summer-active of Mediterranean origin, and bred in the south of France (Volaire, 2002). Parents are early flowering varieties (Shaimi et al., 2009a). All offspring and both parents were tetraploids (2n = 28).
The field experiment was conducted at the Guich experimental station of INRA, Rabat/Morocco (Latitude 34°03′ N, Longitude 06°46′ W, Elevation 10, 5 masl) on an alficxeropsamments soil, having a similar texture throughout with a pH of 6.8 and an organic matter content of 1.2%. An intraspecific crossing between two genotypes of cocksfoot was performed: a genotype of the summer dormant cultivar Kasbah and a genotype of the summer active Mediterranean cultivar Medly, under pollen-proof cages at INRA, Rabat. At maturity, seeds were harvested from parents and were pre-germinated in Petri dishes, transplanted into small pots filled with peat, and maintained under greenhouse conditions. 283 hybrids and parents were cloned and replicated three times. After two months, seedlings were transplanted in a completely randomized block design in a nursery at the Guich experimental station of INRA on February 23, 2011, at the two-tiller stage, with three replicates for each genotype. The space between plots and between rows was 1 m. The trial field was fertilized with 28, 56, 28 kg/ha of (N, P, K) respectively, before planting. In the second year, 2012, the soil was covered with a plastic mulch to prevent weed growth, except plants holes. A nitrogen fertilizer coverage (33% ammonium nitrate) was applied at a rate of 40 units of nitrogen/ha at tillering after each harvest. All plants were maintained well watered in this year (two times per week) to maintain soil moisture at field capacity. Forage was harvested manually at 5 cm, one homogenization harvest in 10 of May 2011, and four harvests in 2012: on 31st of January, 2nd of April, 15th of May and 11th of September.
During 2013, the trial was conducted under rain-fed conditions with minimal N fertilizer applied after cutting. Biomass was collected on 26th January, mid-June and on 9th December, to assess biomass of autumn recovery. Plants experienced a severe summer drought for more than 110 days. Visual plant survival scoring was recorded 14 days after the first significant autumn rain. A single plant was considered surviving when at least one or more green leaves started growing, while totally dark plants were considered as dead.
2.2. Plant measurements
The following traits were recorded in the field: Dry matter biomass of individual plants was weighed after each cut (g/plant), all cuts were made at 5 cm height. Fresh matter from each plant cut was dried in an oven at 70 °C for 72 h. In 2012, heading date was recorded in spring as the number of days from January 1 to the date that more of three tillers reach heading stage. Plant height was measured on 2 April using a graduate rule just before second cuts in 2012. After re-watering, we examined regeneration for new tillers. The percentage of plant senescence was visually scored on 16th of July, conferring to scale (0 = all tissues are green and 100 = no visible green tissues, and used to assess summer dormancy). The summer dormancy index (SDI) was calculated according to Norton index: S/Sp = (100 − [(summer yield/spring yield) × 100])/10, which ranged between 1 = summer active and 10 = fully summer dormant (Norton et al., 2008). The survival for each genotype was scored 14 days after the first significant rainfall in the autumn after the summer drought of 2013.
2.3. Climatic conditions during the experiment
Weather conditions during (2011/2013) are displayed in Fig. 1. The average annual rainfall is 263 mm and 533 mm, in 2012 and 2013 respectively.
Figure 1.

Distribution of monthly precipitation in (mm), temperature (mean, max, min) in (°C) for 2010/2011, 2011/2012 and 2012/2013 at the experimental station Guich/Rabet.
2.4. Statistical analysis
Population genotypes were tested in a randomized complete block design with three replications. Statistical analyses were performed using the SAS software, using the one-way analysis of variance (ANOVA) procedure. The significance of differences between means was evaluated by Duncan multiple range test. Principal component analysis (PCA), was performed using Genstat, 15th edition.
3. Results
The following results will show diversity within cocksfoot population related to dry matter yield, heading date, plant height, senescence, summer dormancy and plant survival. We consider P value (P < 0.005) for all measured traits.
3.1. Dry matter yield
Significant differences were found between hybrids and parents in the total dry matter for the three years of trial. Total dry matter yield means were 4.66 g/plant, 87.44 and 28.35 g/plant for 2011, 2012 and 2013, respectively.
In the third year, both winter and autumn biomasses showed significant differences among populations. Total dry matter yield varied between 4.44 and 108 g/plant. The two parents Kasbah and Medly cultivars yielded 5.34 and 24.57 g/plant, respectively. About 38.8% of the progeny yielded more than Kasbah, but less than the productive parent Medly (Fig. 2), and 57% of the progeny yielded more than both parents. Harvested biomass in late autumn varied between 0.5 and 17.56 g/plant. Indeed, the severe summer drought reduced the biomass of regrowth, many genotypes ceased or limited their growth during summer because of drought coupled with summer dormancy trait.
Figure 2.

Distribution of total dry matter accumulated in 2013 (g/plant) (first and second harvest) in cocksfoot population issued by crossing between Medly and Kasbah.
3.2. Heading date and plant height
Significant differences among genotypes were found for heading date and plant height. Heading date ranged from 58 to 97 days from the first of January in 2012. The average was 77 days 2012, the parent Medly was earlier than Kasbah. Plant height population average was 56.9 cm, the parent Kasbah was slightly shorter than Medly.
3.3. Senescence of aerial tissues
Large variability was shown between genotypes. Plant senescence score during the summer under irrigation, varied from 8.33% to 95% (Fig. 3) within the population. The genotype 91 achieved the highest score, which was not significantly different from that of parent Kasbah that maintained a higher level of 86.67%. The non-dormant parent Medly had slightly less senescence during summer and reached 38.33%. About 62.88% of hybrids had an intermediate score between those of the parents. Duncan rating showed that 4% of genotypes had higher senescence scores than Kasbah.
Figure 3.
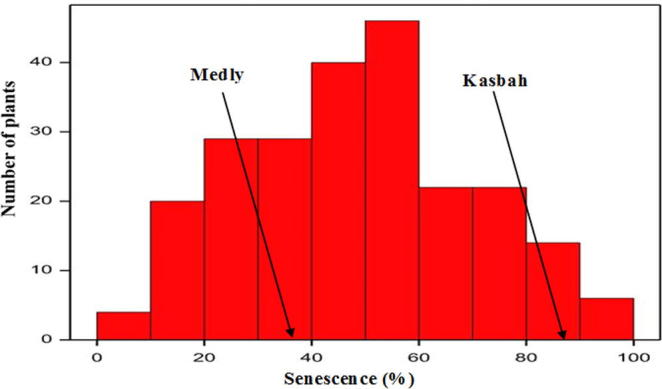
Distribution of senescence scoring in (%) noted under irrigation in July 2012 in cocksfoot population individuals from the summer dormant parent Kasbah and the summer active parent Medly.
Progression of plant senescence under summer drought was quite similar and started earlier. The summer dormant parent Kasbah became completely senescent at the onset of drought. However, about a third of the population had less complete senescence in early summer to reach the complete senescence at mid-summer.
3.4. Summer dormancy index (SDI)
Significant differences were shown between genotypes for the summer dormancy index (SDI) (P < 0.05), calculated according to Norton index (S/Sp) (Norton et al., 2008). SDI varied between 5.05 and 9.85 with an average value of 9.13. The dormant parent ‘Kasbah’ had an important level of dormancy 9.76. A majority of hybrids showed a high level of SDI including parents. The productive parent Medly showed a high SDI, while it is known by its productivity over a year. Moreover, Medly accumulated a great vegetative biomass in spring, but a low summer biomass under irrigation. It is the main reason of the high SDI. It is also related to high temperature during summer.
3.5. Drought survival
During summer 2013, plants experienced 110 days of severe drought before the first rain occurred. Differences were found between genotypes.
The severe drought affected single plant evolution, 78% of hybrids persisted to drastic summer drought, while 21% had moderate survival, varying between 34% and 67%. The decrease was more important for 1% of hybrids, which had disappeared completely by the end of the third year. The distribution of total dry matter accumulated in this year and drought survival score noted after first significant autumn rainfall showed five classes (Fig. 4). In the first class, we find genotypes that can’t persist after the severe drought and dead completely. These genotypes accumulated 10 g/plant. While the second class contains eight genotypes that persisted at 33% and accumulated an important total biomass about 20 g/plant. About 7% of hybrids achieved 22 g/plant and persisted incompletely to summer conditions. Besides, 9% of the progeny persisted highly to drought and marked a survival score of 67% accumulated 23 g/plant. The last class comprised the majority of genotypes (79%), including both parents. These genotypes presented an interesting material since they achieved a great drought survival scoring 100% and accumulated 30 g/plant on average.
Figure 4.
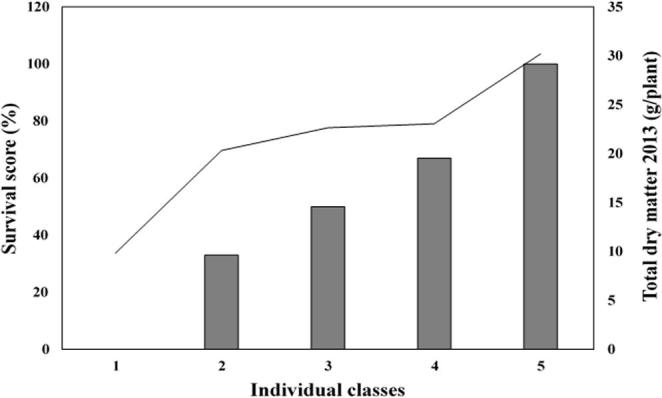
Distribution of total dry matter accumulated in 2013 (g/plant) (first and second harvest) and survival score (%) noted after first autumn significant rainfall in December 2013 after severe summer drought.
3.6. Principal Component Analyses (PCA)
PCA using summer dormancy scored under summer irrigation in 2012 and drought survival in 2013 and total dry matter accumulated between first and second harvest in 2013, and biomass recovery after autumn rainfall showed that two components explained 66.35% of total variability between four measured traits. Four groups were identified, and two independent genotypes were distinguished (Fig. 5).
Figure 5.

Distribution of 4 variables on the ordination produced by the two axes of PCA. SD2012-Summer dormancy under summer irrigation in 2012, DS2013-Drought survival in 2013, TDM2013-Total dry matter accumulated from the first and the second harvest in 2013 and ADM2013-Autum recovery dry matter recorded after significance autumn rainfall in 2013.
The genotypes of the first group, including Kasbah, are completely summer drought survivable and had an important level of summer dormancy. They achieved less total biomass production and persisted with less biomass in autumn recovery. The second group includes the productive parent Medly, that survived to summer drought. These genotypes had a complete summer drought survival, achieved a great level of total dry matter and autumn recovery biomass more important than the first group. The third group had a good level of drought survival, contains the higher biomass production genotypes from the whole progeny which had the lower summer dormancy whereas they persisted and recovered well after the severe summer drought. The genotypes of the fourth group were less drought surviving moreover, they had a moderate summer dormancy. They yielded a less autumn recovery and total biomass. Two distinguished genotypes were independent of other groups: 211 died in all repetitions, and 285: a control for a temperate individual, was non-dormant.
4. Discussion
The correlations between traits are presented in Table 1.
Table 1.
Pearson’s correlation coefficients among measured traits in the establishment, under irrigation (2011, 2012), under rainfed conditions (2013). We consider (P < 0.005) for all measured traits.
| Traits | HD2012 | Senesc2012 | SD2012 | DS2013 | ADM2013 |
|---|---|---|---|---|---|
| TDM2011 | −0.16 | ||||
| TDM2012 | −0.52 | ||||
| PH2012 | −0.58 | ||||
| SpB2012 | −0.21 | 0.28 | |||
| SD2012 | 0.40 | −0.16 | |||
| TDM2013 | −0.20 | 0.18 | |||
| DS2013 | −0.27 |
TDM: Total dry matter; ADM: Autumn recovery dry matter; SpB: Spring biomass; PH: Plant height; HD: Heading date; SD: Summer dormancy; Senesc: Senescence; DS: Drought survival.
4.1. Dry matter yield
The obtained results showed a large variability between hybrids for the total dry matter accumulated in the second and the third years. In the third year, the summer dormant variety exhibited a higher yield than the others due to their lack of persistency under severe droughts. Similar to the finding of Norton (2014) in southern Australia. A significant positive correlation was observed between drought survival and total dry matter accumulated in 2013 (r = 0.18, P < 0.005), Shaimi et al. (2009a) reported a similar correlation.
4.2. Heading date and plant height
Results showed that genotypes with an earlier heading date can pass through the different stages of development earlier than varieties with a later heading date (Feuerstein and Swieter, 2014). The analysis showed that Heading date was negatively correlated with total dry matter even in the first year (r = −0.16, P < 0.005) or in the second year (r = −0.52, P < 0.005). Similar correlation was found between heading date and plant height (r = −0.58, P < 0.005) with a large variability between genotypes, consistent with the finding of Hazard et al. (2006) in ryegrass populations.
4.3. Senescence of aerial tissues
Senescence, as the last stage of leaf development under summer irrigation manifested by a decrease in the length of green leaf (Duru and Ducrocq, 2000). Under summer irrigation, senescence was inversely correlated with the spring biomass (r = −0.21, P < 0.005), similar to our previous study concerning the assessment of the same plant material for some phenotypical parameters (Zhouri et al., 2016). Despite this correlation, some hybrids were good senescent and productive. Leaf senescence was inversely correlated to plant survival (r = −0.27, P < 0.005). A similar correlation was found on perennial species under drought (Zwicke et al., 2015). Inversely, gradual foliage senescence was positively correlated with plant survival in a range of Mediterranean grasses (Volaire et al., 1998, Volaire and Lelievre, 2001, Pérez-Ramos et al., 2013).
4.4. Summer dormancy index (SDI)
A significant negative correlation was found between total dry matter accumulated in the third year and summer dormancy (r = −0.20, P < 0.005). Shaimi et al. (2009a) found a similar correlation between summer dormancy and annual forage production. Whereas, a positive correlation was found between spring biomass accumulated in the second year and summer dormancy (r = 0.28, P < 0.005). Summer dormancy seems to be positively correlated to leaf senescence (r = 0.40, P < 0.005). Earlier research realized by Norton et al. (2006b) on Festuca arundinacea Schreb suggested that leaf senescence could be used to identify summer dormant plants. A significant negative correlation was achieved between summer dormancy and autumn recovery yield (r = −0.16, P < 0.005). Probably, summer dormancy is associated with reduced water consumption (Lolicato, 2000) and generally promote survival in drier, hotter areas (Culvenor and Boschma, 2005).
4.5. Drought survival
This distribution showed that more a plant had an important total dry matter more it can persist to several conditions. No correlation was found between autumn recovery and drought survival. Although, autumn recovery biomass gives evidence of hybrids and parents perenniality. After first significant autumn rainfall, 58% of hybrids persisted completely and accumulated an autumn biomass more important than both parents. These hybrids presented a powerful material to persist to summer drought. The biomass of recovery informs about hybrid persistence through summer drought. Indeed, the summer survival of Moroccan cocksfoot ecotypes grown in population was higher of that of European lines and reached 97% versus 59%, respectively (Shaimi et al., 2009a).
4.6. Principal Component Analyses (PCA)
This distribution showed that complete drought surviving genotypes had the best summer dormancy, but accumulated an intermediate total dry matter and autumn recovery. Nevertheless, summer dormancy is one of the main traits conferring drought survival and autumn recovery of perennial forage species in Mediterranean areas (Shaimi et al., 2009a). Also, Medly survived under summer drought. It’s due to its higher dehydration avoidance correlated to its Mediterranean origin (Volaire et al., 1998). An earlier research on several grass species showed that superior survival after severe and repeated summer droughts is generally correlated with summer dormancy (Norton et al., 2006a, Norton et al., 2012). Moreover, summer dormancy is a powerful trait for survival under severe drought (Norton et al., 2007). Mainly, we can explain these obtained results by a photosynthetic mechanism. Under severe drought, a reduction of photosynthesis, stomatal conduction, and biochemical limitation is manifested. Moreover, a strong contribution of carbohydrate to drought survival was shown (Xu et al., 2010, Xu and Zhou, 2011). In fact, the accumulation of soluble carbohydrate is necessary for osmotic adjustment (Chaves, 1991). Other traits such as water-soluble carbohydrate (WSC) concentrations in tiller bases may also be useful in developing drought tolerance (Volaire and Lelievre, 1997). Consequently, the total WSC reserves in leaf bases of cocksfoot plants increased by 35% on average during drought (Volaire and Lelievre, 1997). Especially, fructans are the most abundant WSC in some perennial grasses such as cocksfoot and tall fescue. Generally, species that accumulate fructans were correlated with environments where water availability was limited (Hendry, 1993). Moreover, WSC was slightly associated with plant survival after autumn rains (Volaire, 1991). So we can suggest that our surviving genotypes concentrated carbohydrate to the escape drought period. Immediately, after rewatering, production and accumulation of green herbage started the recoveries of plant growth and photosynthesis appears over growing new plant (Xu et al., 2010). A clear association appeared between summer dormancy and drought survival, the same results have been reviewed by (Volaire and Norton, 2006). We can conclude that dormancy and biomass production over the year are powerful traits to survive the severe drought.
4.7. Conclusion
The results, according to the measured parameters for hybrids issued from crossing between two contrasting varieties of cocksfoot tested under Moroccan climate conditions, suggested a considerable variability for all measured traits, such as dry matter yield, heading date, senescence score, summer dormancy index, and drought survival.
These results show that some hybrids in progeny were able to regrow after severe drought and produce better than parents in spring under rain-fed conditions. These genotypes can regrow and stay alive in all repetitions. They constitute a promising material to climate change and especially to summer drought conditions, they even present a good perenniality which can be used for a further breeding program. Indeed, summer drought survival genotypes had a significant autumn recovery after the severe summer drought and accumulated a good level of total dry matter in spring, having an important summer dormancy index.
Hybrids and parent cultivars tested under Moroccan climate were able to stand severe drought stress levels in the stressful conditions. Many hybrids tended to outperform parental cultivars for yield and persistence. Probably this material will have the interest to enhance both yield and persistence when aridity increases.
Funding
This work has been funded by the Partenariat Hubert Curien PRAD France-Morocco and by the Institut National de la Recherche Agronomique (INRA) metaprogram ACCAF ‘Climagie’.
Acknowledgements
This work has been founded by a Partenariat Hubert Curien PRAD France-Maroc and by the INRA metaprogram ACCAF (Climagie). The authors are grateful to P. Dorchies, F. Prévot and the team of «Agence française pour la promotion de l’enseignement supérieur, l’accueil et la mobilité internationale» for their help in the organisation of exchanges between France and Morocco. We thank all persons who participated in the execution of the field work and in shaping up the manuscript. Special appreciation is due to Ms. Chawki Al Faiz who played a part in the building of this project also to Ms. El Housni Abdallah, Ms. Douaik Ahmed and Ms. Saadi Abdenbi for their help.
Footnotes
Peer review under responsibility of King Saud University.
References
- Annicchiarico P., Pecetti L., Abdelguerfi A., Bouizgaren A., Carroni A., Hayek T., Bouzina M.H., Mezni M. Adaptation of landrace and variety germplasm and selection strategies for lucerne in the Mediterranean basin. Field Crops Res. 2011;120:283–291. [Google Scholar]
- Bell L.W., Williams A.H., Ryan M.H., Ewing M.A. Water relations and adaptations to increasing water deficit in three perennial legumes, Medicago sativa, Dorycnium hirsutum and Dorycnium rectum. Plant Soil. 2007;290:231–243. [Google Scholar]
- Bolger, T., Garden, D., Rivelli, A.R., 2007. Drought resistance of perennial grasses. In: Proceedings of the 22nd Annual Conference of the Grassland Society of NSW. Pasture Systems. Managing for a Variable Climate, Queanbeyan, ACT. pp. 60–61.
- Chaves M. Effects of water deficits on carbon assimilation. J. Exp. Bot. 1991;42:1–16. [Google Scholar]
- Chimner R., Welker J., Morgan J., LeCain D., Reeder J. Experimental manipulations of winter snow and summer rain influence ecosystem carbon cycling in a mixed-grass prairie, Wyoming, USA. Ecohydrology. 2010;3:284–293. [Google Scholar]
- Cullen B., Johnson I., Eckard R., Lodge G., Walker R., Rawnsley R., McCaskill M. Climate change effects on pasture systems in south-eastern Australia. Crop Pasture Sci. 2009;60:933–942. [Google Scholar]
- Culvenor R., Boschma S. Evaluation of phalaris (Phalaris aquatica L.) germplasm for persistence under grazing on the North-West Slopes, New South Wales. Aust. J. Agric. Res. 2005;56:731–741. [Google Scholar]
- Dukes J.S., Chiariello N.R., Cleland E.E., Moore L.A., Shaw M.R., Thayer S., Tobeck T., Mooney H.A., Field C.B. Responses of grassland production to single and multiple global environmental changes. PLoS Biol. 2005;3:1829–1837. doi: 10.1371/journal.pbio.0030319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duru M., Ducrocq H. Growth and senescence of the successive leaves on a Cocksfoot tiller. Effect of nitrogen and cutting regime. Ann. Bot. 2000;85:645–653. [Google Scholar]
- Feuerstein U., Swieter A. Fair evaluation of yield potential in forage species evaluation. In: Sokolović D., Huyghe C., Radović J., editors. Quantitative Traits Breeding for Multifunctional Grasslands and Turf. Springer; 2014. pp. 131–135. [Google Scholar]
- Hazard L., Betin M., Molinari N. Correlated response in plant height and heading date to selection in perennial ryegrass populations. Agron. J. 2006;98:1384–1391. [Google Scholar]
- Hendry G.A. Evolutionary origins and natural functions of fructans-a climatological, biogeographic and mechanistic appraisal. New Phytol. 1993;123:3–14. [Google Scholar]
- Howden S., Crimp S., Stokes C. Climate change and Australian livestock systems: impacts, research and policy issues. Anim. Prod. Sci. 2008;48:780–788. [Google Scholar]
- IPCC . Cambridge University Press; Cambridge, UK: 2007. Climate Change 2007: Impacts, Adaptation and Vulnerability: Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; p. 938. [Google Scholar]
- Kallida R., Shaimi N., Al Faiz C. Drought survival of some perennial grasses in Moroccan rainfed conditions: agronomic traits. In: Huyghe C., editor. Sustainable Use of Genetic Diversity in Forage and Turf Breeding. Springer; 2010. pp. 113–118. [Google Scholar]
- Kemp D., Culvenor R. Improving the grazing and drought tolerance of temperate perennial grasses. N.Z. J. Agric. Res. 1990;37:365–378. [Google Scholar]
- Knapp A.K., Smith M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science. 2001;291:481–484. doi: 10.1126/science.291.5503.481. [DOI] [PubMed] [Google Scholar]
- Lelièvre F., Volaire F. Current and potential development of perennial grasses in rainfed Mediterranean farming systems. Crop Sci. 2009;49:2371–2378. [Google Scholar]
- Lelièvre F., Seddaiu G., Ledda L., Porqueddu C., Volaire F. Water use efficiency and drought survival in Mediterranean perennial forage grasses. Field Crops Res. 2011;121:333–342. [Google Scholar]
- Lolicato S. Soil water dynamics and growth of perennial pasture species for dryland salinity control. Anim. Prod. Sci. 2000;40:37–45. [Google Scholar]
- Norton M.R. Proceedings of the 52nd Annual Conference of the Grassland Society of Southern Australia. Grassland Society of Southern Australia Inc.; Echuca, Victoria: 2011. Improving the persistence of temperate perennial pastures with the summer dormancy trait; pp. 57–63. [Google Scholar]
- Norton M.R. Farming Ahead; Perth, WA: 2014. Summer Dormant Cocksfoot A Great Pasture Option; p. 266. [Google Scholar]
- Norton M.R., Lelievre F., Volaire F. Summer dormancy in Dactylis glomerata L., the influence of season of sowing and a simulated mid-summer storm on two contrasting cultivars. Aust. J. Agric. Res. 2006;57:565–575. [Google Scholar]
- Norton M.R., Volaire F., Lelievre F. Summer dormancy in Festuca arundinacea Schreb; the influence of season of sowing and a simulated mid–summer storm on two contrasting cultivars. Aust. J. Agric. Res. 2006;57:1267–1277. [Google Scholar]
- Norton, M.R., Volaire, F., Fukai, S.L., Lelièvre, F., 2007. Summer dormancy-a drought resistance strategy in perennial grasses. In: Proceedings of the 22nd Annual Conference of the Grassland Society of NSW. Pasture Systems. Managing for a Variable Climate, Queanbeyan, ACT, pp. 58–59.
- Norton M.R., Lelievre F., Fukai S., Volaire F. Measurement of summer dormancy in temperate perennial pasture grasses. Aust. J. Agric. Res. 2008;59:498–509. [Google Scholar]
- Norton M.R., Volaire F., Lelièvre F., Fukai S. Identification and measurement of summer dormancy in temperate perennial grasses. Crop Sci. 2009;49:2347–2352. [Google Scholar]
- Norton M.R., Lelièvre F., Volaire F. Summer dormancy in Phalaris aquatica L., the influence of season of sowing and summer moisture regime on two contrasting cultivars. J. Agron. Crop Sci. 2012;198:1–13. [Google Scholar]
- Ogle D., John L.S., Stannard M., Holzworth L. USDA-Natural Resources Conservation Service; Boise, ID: 2011. Conservation Plant Materials for the Intermountain West; p. 57. [Google Scholar]
- Pecetti L., Annicchiarico P., Abdelguerfi A., Kallida R., Mefti M., Porqueddu C., Simões N.M., Volaire F., Lelièvre F. Response of mediterranean tall fescue cultivars to contrasting agricultural environments and implications for selection. J. Agron. Crop Sci. 2011;197:12–20. [Google Scholar]
- Pérez-Ramos I.M., Rodríguez-Calcerrada J., Ourcival J.M., Rambal S. Quercus ilex recruitment in a drier world: a multi-stage demographic approach. Perspect. Plant Ecol. Evol. Syst. 2013;15:106–117. [Google Scholar]
- Salis, M., Seddaiu, G., Ledda, L., Lloveras, J., González-Rodríguez, A., Vázquez-Yañez, O., Piñeiro, J., Santamaría, O., Olea, L., Poblaciones, M., 2006. Drought resistance evaluation of perennial grasses. In: Proceedings of the 21st General Meeting of the European Grassland Federation, Badajoz (Spain) 3–6 April 2006. pp. 60–70.
- Shaimi N., Kallida R., Volaire F., Faiz C.A. Summer dormancy in orchard grass: evaluation and characterization through ecophysiological and genetic studies. Crop Sci. 2009;49:2353–2358. [Google Scholar]
- Shaimi N., Kallida R., Volaire F., Saidi N., Faiz C.A. Summer dormancy and drought survival of Moroccan ecotypes of orchard grass. Crop Sci. 2009;49:1416–1424. [Google Scholar]
- Volaire F. Agronomical evaluation of local populations of coocksfoot (Dactylis glomerata L.) in Corsica. Agric. Medit. 1991;121:263–271. [Google Scholar]
- Volaire F. Effects of summer drought and spring defoliation on carbohydrate reserves, persistence and recovery of two populations of cocksfoot (Dactylis glomerata) in a Mediterranean environment. J. Agric. Sci. 1994;122:207–215. [Google Scholar]
- Volaire F. Growth, carbohydrate reserves and drought survival strategies of contrasting Dactylis glomerata populations in a Mediterranean environment. J. Appl. Ecol. 1995;32:56–66. [Google Scholar]
- Volaire F. Drought survival, summer dormancy and dehydrin accumulation in contrasting cultivars of Dactylis glomerata. Physiol. Plant. 2002;116:42–51. doi: 10.1034/j.1399-3054.2002.1160106.x. [DOI] [PubMed] [Google Scholar]
- Volaire F. Plant traits and functional types to characterise drought survival of pluri-specific perennial herbaceous swards in Mediterranean areas. Eur. J. Agron. 2008;29:116–124. [Google Scholar]
- Volaire F., Lelievre F. Production, persistence, and water-soluble carbohydrate accumulation in 21 contrasting populations of Dactylis glomerata L. subjected to severe drought in the south of France. Aust. J. Agric. Res. 1997;48:933–944. [Google Scholar]
- Volaire F., Lelievre F. Drought survival in Dactylis glomerata and Festuca arundinacea under similar rooting conditions in tubes. Plant Soil. 2001;229:225–234. [Google Scholar]
- Volaire F., Norton M.R. Summer dormancy in perennial temperate grasses. Ann. Bot. 2006;98:927–933. doi: 10.1093/aob/mcl195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volaire F., Thomas H., Lelievre F. Survival and recovery of perennial forage grasses under prolonged Mediterranean drought: I. Growth, death, water relations and solute content in herbage and stubble. New Phytol. 1998;140:439–449. doi: 10.1111/j.1469-8137.1998.00288.x. [DOI] [PubMed] [Google Scholar]
- Volaire F., Norton M.R., Lelièvre F. Summer drought survival strategies and sustainability of perennial temperate forage grasses in Mediterranean areas. Crop Sci. 2009;49:2386–2392. [Google Scholar]
- Volaire F., Barkaoui K., Norton M. Designing resilient and sustainable grasslands for a drier future: adaptive strategies, functional traits and biotic interactions. Eur. J. Agron. 2014;52:81–89. [Google Scholar]
- Xu Z., Zhou G. Responses of photosynthetic capacity to soil moisture gradient in perennial rhizome grass and perennial bunchgrass. BMC Plant Biol. 2011;11:1. doi: 10.1186/1471-2229-11-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Z., Zhou G., Shimizu H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010;5:649–654. doi: 10.4161/psb.5.6.11398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhouri L., Kallida R., Shaimi N., Barre P., Volaire F., Fakiri M. Analysis of progeny for productivity traits and summer dormancy of Dactylis glomerata L. Ekin J. 2016;2:36–39. [Google Scholar]
- Zwicke M., Picon-Cochard C., Morvan-Bertrand A., Prud'homme M.-P., Volaire F. What functional strategies drive drought survival and recovery of perennial species from upland grassland? Ann. Bot. 2015;116:1001–1015. doi: 10.1093/aob/mcv037. [DOI] [PMC free article] [PubMed] [Google Scholar]