

Abstract
Background
Regular loading of tendons may counteract the negative effects of aging. However, the influence of strength training loading magnitude on tendon mechanical properties and its relation to matrix collagen content and collagen cross-linking is sparsely described in older adults. The purpose of the present study was to compare the effects of moderate or high load resistance training on tendon matrix and its mechanical properties.
Methods
Seventeen women and 19 men, age 62–70 years, were recruited and randomly allocated to 12 months of heavy load resistance training (HRT), moderate load resistance training (MRT) or control (CON). Pre- and post-intervention testing comprised isometric quadriceps strength test (IsoMVC), ultrasound based testing of in vivo patellar tendon (PT) mechanical properties, MRI-based measurement of PT cross-sectional area (CSA), PT biopsies for assessment of fibril morphology, collagen content, enzymatic cross-links, and tendon fluorescence as a measure of advanced glycation end-products (AGEs).
Results
Thirty three participants completed the intervention and were included in the data analysis. IsoMVC increased more after HRT (+ 21%) than MRT (+ 8%) and CON (+ 7%) (p < 0.05). Tendon stiffness (p < 0.05) and Young’s modulus (p = 0.05) were also differently affected by training load with a reduction in CON and MRT but not in HRT. PT-CSA increased equally after both MRT and HRT. Collagen content, fibril morphology, enzymatic cross-links, and tendon fluorescence were unaffected by training.
Conclusion
Despite equal improvements in tendon size after moderate and heavy load resistance training, only heavy. load training seemed to maintain tendon mechanical properties in old age. The effect of load magnitude on tendon biomechanics was unrelated to changes of major load bearing matrix components in the tendon core.
The study is a sub-study of the LISA study, which was registered at http://clinicaltrials.gov (NCT02123641) April 25th 2014.
Keywords: Tendon biomechanics, Collagen cross-links., Strength training., Aging.
Background
Aging negatively affects structure and mechanical function of collagen-rich tissues [1–3]. The skin loses elasticity, blood vessels become stiffer, and tendons may become more compliant [4]. The possible age-related decline in tendon stiffness may in turn reduce maximal skeletal muscle performance [5] and postural balance [6], which negatively affects mobility in old age. Regular loading of tendons seems to protect the tissue from the negative effects of aging [7, 8] by increasing tensile stiffness [9–11]. However, the knowledge about training duration and the magnitude of load needed to induce favorable adaptations in tendons of older adults is limited. Moreover, the relation between tendon mechanical properties and molecular changes in the matrix is largely unresolved.
Training with heavy load seems to elicit a more pronounced increase in tendon stiffness than training with moderate loads in middle aged [12] as well as older adults [11], although low load training has also been demonstrated to affect tendon mechanical properties [4]. Short term (12 weeks) resistance training has effectively increased tendon stiffness in older adults in most [9–11] but not all [13] studies. Only few studies have investigated the effect of very long training duration and found no additional effect on tendon stiffness after either 18 months (older adults) [10] or 4 years (young adults) [14] compared to only three months of training. However, young athletes engaging in sports where one leg is more loaded than the other over several years display pronounced differences in tendon stiffness and cross-sectional area, suggesting a continued adaptation to loading beyond the first three months [15]. Thus, more studies are warranted to investigate the influence of training load over a long duration of training on tendon size and mechanical properties, especially in older adults.
The main contributor to tendon tensile stiffness is type I collagen, which makes up most of the tendon matrix. Acute exercise increases tendon collagen expression in animals [16] and peritendionus collagen synthesis in humans [17], but the overall collagen content and the collagen fibril volume fraction as measured by transmission electron microscopy (TEM) does not seem to be responsive to training [8, 18]. Moreover, recent evidence suggest that the vast majority of core tendon collagen molecules are not renewed after late teenage years [19], suggesting that the aging and loading induced adaptations in tendon mechanical properties are mediated by other molecular components in the matrix.
Another important matrix component that could influence tendon mechanical properties are collagen cross-links [20], which are essentially either enzymatic or non-enzymatic. Enzymatic cross-links (i.e. lysylpyrridinoline (LP) and hydroxylysylpyrridinoline (HP)) are tightly regulated in time and space by the family of lysyl oxidases (LOX) and are essential for normal tendon development. Tendon LOX expression is unaffected by regular training [21] but induced by acute exercise [16] indicating a stimulatory effect of exercise on formation of cross links. Life-long endurance training does not seem to affect HP or LP [8], but it remains unknown if long-term resistance training increases enzymatic cross-link density in older adults and if this is related to increased tendon mechanical properties.
Non-enzymatic cross-links form sporadically when reducing sugars react with amino-groups and mature into advanced glycation end-products (AGEs) [1, 2]. AGEs accumulate in collagen rich tissues with age [22], not least in tissues with slow turnover rate like tendon [23, 24], where they may eventually impair material properties [3, 25, 26]. Despite the slow turnover rate of tendon, regular training seems to reduce [21] or at least be associated with an attenuated AGE accumulation in older adults [8], and heavy resistance training reduces patellar tendon pentosidine, which is a cross-linking AGE, in young men [27]. However, it is still unknown how regular heavy and light load resistance training affects tendon AGEs in older adults. Moreover, although cross-linking AGEs increase tendon mechanical properties in animals [26, 28], they may also have the opposite effect by inducing fibril damage accumulation [3, 25] or by taking up space in the matrix [29], and the relation between tendon AGEs and mechanical properties in old humans needs to be clarified [30].
The present investigation aimed to compare the effects of long-term (12 months) heavy or light load resistance training on patellar tendon mechanical properties, morphology and collagen cross-links in older adults as well as the relation between changes in mechanical properties and tendon matrix. Our hypothesis was that compared to a control group, both training loads would increase patellar tendon stiffness. Moreover, we hypothesized that the increased tendon stiffness would primarily be related to greater density of enzymatic cross-links, whereas AGE cross-link accumulation would be attenuated and collagen content and fibril morphology would remain unchanged.
Methods
Study design
The present investigation was a sub-study of a randomized trial started in 2014 at the Institute of Sports Medicine Copenhagen [31]. The study primarily investigated the effect of 12 months resistance training on muscle strength and physical function in 450 men and women, 62–70 years old. The 36 participants recruited for the present sub-study gave consent to undergo additional tendon specific tests. They were allocated to 12 months of moderate load resistance training (MRT), heavy load resistance training (HRT), or habitual physical activity level (< 1 h strenuous exercise/week) (CON).
Participants
Based on expected changes in tendon fluorescence of approximately 20%, which is what has been observed previously for pentosidine (a marker of AGEs) [27], and using the observed pooled SD of approximately 200 found in the present investigation, we needed 10 participants in each group to reach 80% power. The study included 17 women and 19 men, age 62–70 years. The participants were all home-dwelling, independent, and untrained. Exclusion criteria were more than 1 h of systematic strenuous exercise per week, systemic hormonal−/anti-hormonal therapy, anticoagulant therapy, a history of patellar tendinopathy within the past 12 months, chronic knee pain or other musculoskeletal problems impeding participation in resistance training. For the full list of exclusion criteria we refer to the original study protocol [31].
Interventions
The interventions are described in detail in a previous publication [31]. Briefly, HRT was a supervised, whole body, progressive, heavy resistance training program. In the first 6–8 weeks of the intervention the participants performed 3 sets of 15 repetitions at a moderate load (~ 50–60% of one repetition maximum (1RM)) to accustom them to the exercises. After this, the intervention consisted of periods of 8 weeks in which the number of repetitions was gradually reduced from 12 to 6 while the load was accordingly increased from 70 to 85% of 1RM. The periods of 8 weeks were repeated throughout the 12 months intervention, and one week of restitution (no training) was interspersed between each period. The leg exercises targeting the patellar tendon were leg press and knee-extension performed in Technogym fitness machines (TechnoGym, Gambettola, Cesena, Italy).
MRT was a home-based, whole body, progressive, moderate load resistance training program performed as circuit training and using TheraBand® elastic rubber bands (Hygenic Corp., Akron, OH, USA) and the participants own body-weight. The number of repetitions was gradually increased from 3 sets of 12 to 3 sets of 18 repetitions. Load was increased whenever the participants could perform 3 sets of 18 repetitions without feeling exhausted, at which point the number of repetitions were once again reduced to 3 sets of 12. The exercises targeting the patellar tendon were squat and knee-extension. Training frequency was three times per week, and the participants were supervised once a week. Participants recorded training activity (load, number of sets and repetitions) in training diaries.
CON was encouraged not to change their habitual physical activity level, which was < 1 h of regular strenuous physical activity per week, and were offered participation in social/cultural activities approximately once a month.
Muscle strength assessment
An investigator blinded to group allocation used a Good Strength dynamometer (V.3.14 Bluetooth; Metitur, Finland) to assess isometric knee extensor strength (IsoMVC) at 700 knee flexion. The participants performed one familiarization trial before recording at least three verbally encouraged maximal isometric knee-extensions. Measurements were continued until no further improvement occurred [31].
Muscle size
An experienced radiographer blinded to group allocation acquired MRI of the thighs using a 3.0 T TX Philips Achieva scanner (Philips Healthcare, Amsterdam, The Netherlands) with a 32-channel body array coil. The scanning was a 2D axial T1-weighted sequence (TR/TE = 666/20 ms, FA = 90, 672 × 672 matrix, 3 stacks with 3 slices, gap 1.91 mm, in-plane resolution 0.8 mm, slice thickness 4 mm) acquired at 10, 20 and 30 cm above tibia plateau, with three slices at each level for delineation of vastus lateralis of the quadriceps muscle and calculation of cross-sectional area. Image analysis was conducted by blinded assessors using the medical imaging software package Jim version 6.0 (Xynapse Systems, UK). The reported values for vastus lateralis cross-sectional area (VL-CSA) are from one measurement 20 cm proximal to the tibia plateau. In a few cases, the slice at the same location at follow-up was not visually consistent with that at the baseline measurement, due to small variations in the location of the slice matrix. Instead the most consistent neighboring slice (closer to 19.4 or 20.6 cm proximal to the tibia plateau) was used. We report VL-CSA of the dominant leg only [31].
Blood measurements
A general health screening was performed at baseline including several analyses on blood samples. Blood samples were obtained by medical doctors or medical students. Blood analysis was performed with standard assays and kits at the clinical biochemistry department at Bispebjerg Hospital (Copenhagen, Denmark). We report here values for HbA1c and total cholesterol since they may affect tendon health and mechanics [32–35].
Physical activity level
Physical activity level was measured with a previously validated [36] accelerometer and inclinometer (activPal micro, PAL technologies, Glasgow, Scotland), which was mounted on the thigh of the participants for five consecutive days, always including the weekend. Data were extracted with activPal software (Research edition, V.7.2.32, PAL Technologies, 2013). Here we report daily step count as a measure of physical activity level.
Patellar tendon mechanical properties
The procedure for testing patellar tendon mechanical properties is a validated method, which has been used previously in our own and other laboratories [11, 13, 37]. The participants initially performed 5 min warm-up on a cycle ergometer (Monark, Sweden) at a low resistance to precondition the patellar tendon. The participants were then seated in a custom-made rigid chair with the hip- and knee-joints fixed in a 900 angle. They then performed 8 s ramped isometric contractions with simultaneous recording of knee extensor force using a dynamometer (Noraxon Telemyo 2400 T G2, USA) and ultrasound (US) videos of the patellar tendon with a 10 MHz, 100 mm linear array transducer (Hitachi Hi Vision, Ascendus, Tokyo, Japan) to assess tendon elongation. We recorded 4–6 ramps to ensure at least 2–3 satisfactorily completed ramped contractions with a steadily increasing force production and ultrasound videos with a good contrast. US probe positioning at POST measurement was reproduced by comparison to the PRE measurement recordings.
Patellar tendon elongation was defined as the change in distance between the patella and tibia insertions of the tendon during the ramped contractions, and analyzed with a previously validated custom made semi-automated software [38]. Patella and tibia movements were always tracked within the area of patellar tendon insertion to reduce error due to rotation of tibia and patella during the isometric contraction. Patellar tendon force was calculated by multiplying dynamometer force with the external moment arm (distance from the lateral center of the leg cuff to the lateral collateral ligament) and dividing by the internal moment arm estimated as a percentage (6.5%) of femur length [39]. Femoral length was estimated as the distance from the most lateral point of trochanter major to the lateral collateral ligament. The measurement of external moment arm showed good reproducibility from PRE to POST with a coefficient of variation (CV) of 1,3%, indicating reproducible positioning of the leg-cuff. Second order polynomials were fitted to the force-deformation data points in Sigma Plot (Version 10.0, Systat Software, Germany) and used to estimate maximal values for tendon stiffness at the top 10% of the force-deformation relationship. Tendon deformation was normalized to strain by dividing with initial tendon length, and force was normalized to stress by dividing with average (total) tendon cross sectional area (CSA). Young’s modulus was calculated as the stress to strain ratio also in the top 10% of the stress-strain curve.
The investigators performing data analysis selected two trials for further analysis based on the following selection criteria: Visual consistency between bone movement and tracking points, good synchronization between force and deformation, a smooth inclining force curve, return to baseline after relaxation, and all other things equal, the trials with highest force and/or deformation were chosen. Because the force-deformation relationship of tendon is nonlinear, the raw data in the selected curves was cut off at the highest common force across repeated measurements for each individual.
US videos from 8 participants were analyzed using a custom implementation of cross-correlation tracking in Matlab (R2015b, MathWorks Inc., USA) due to technical difficulties with the video analysis in the program otherwise used. In all instances, we made sure to reanalyze both 0 and 12 months measurements so repeated measures on the same subject were analyzed with the same method.
Patellar tendon dimensions
Patellar tendon length and CSA were assessed with a different MR-scanner than that used for muscle size determination. MRI is a reliable and accurate method for determining patellar tendon dimensions [40]. Trained radiographers performed the scans on a Signa Horizon LX 1.5 T MRI scanner (General Electic, Milwaukee, WI, USA) with an axial and sagittal T1-weighted turbo spin echo sequence (TE: 17; TR: 500; matrix: 512 × 512; FOV: 150 mm; Slice thickness: 3 mm), which has been used in previous human studies in our department [41]. The axial slices of the patellar tendon were positioned orthogonal to the length in the sagittal plane covering the distal patellar pole to the tibia insertion. A supportive pillow was placed in the knee coil to ensure slight stretch on the tendon by bending the knee, which made it easier to measure tendon dimensions. A phantom containing 1.0% CuSO4 was included in the image and subsequently used to adjust contrast settings.
All participants were scanned in their habitual state in the afternoon, and were instructed to avoid strenuous physical activity in the preceding 48 h to avoid the possible influence of training on tendon and muscle water content [42].
Patellar tendon dimensions were assessed using Osirix imaging software (version 2.7.5, Osirix Imaging Medical, Geneva, Switzerland) to manually outline patellar tendon length as well as CSA at three locations (proximal, mid, distal) along the length of the tendon [15, 40]. All images were adjusted according to the phantom and measured using NIH (National Institute of Health) color scale, because this method provides more accurate measurements of patellar tendon CSA [40]. The proximal CSA was measured just distal to the patellar insertion, the distal CSA was measured just proximal to the tibia insertion, and the mid CSA on the slice midway between the proximal and distal slices. Using this procedure, tendon CSA was assessed at the same location at PRE and POST measurements (Fig. 1). Patellar tendon length was measured as the distance from the most dorsal insertion on the patella apex to the most dorsal insertion on the tibia. Coefficient of variation corrected for small sample size on the triplicate measurements was on average 1.5% (range: 0.2–3.2%) for patellar tendon length, and 2.3% (range: 0.2–6.1%) for total patellar tendon CSA. Day to day variation in measurements yielded a CV of 2.4%.
Fig. 1.

Typical magnetic resonance images of the proximal patellar tendon from the same subject before (PRE) and after (POST) 12 months intervention. The two images are obtained at the same location
Tendon biopsies
The patellar tendon biopsy procedure is a sterile procedure, which has been performed previously in our lab [43]. Briefly, the biopsy was obtained in local anesthesia (1 ml lidocaine, Lidokain Mylan 1 mg/ml, Mylan, Oslo, Norway) through a medio-lateral skin incision just distal to the patella with a semi-automated biopsy instrument (Bard Magnum, Bard biopsy systems, USA) at an angle of 450 relative to the patellar tendon in the proximal to distal direction.
We obtained all biopsies at the same time of day before and after the intervention (± 1 h) to avoid the potential influence of circadian rhythm on tendon physiology [44] The non-dominant leg was biopsied before and the dominant leg after the intervention to avoid the influence of the first biopsy on the second [45]. The same trained physician obtained or supervised all biopsies except for four biopsies obtained by another experienced physician. Training was initiated between 5 and 42 days after the PRE biopsy with an average ± SD of 17 ± 9 days. Besides local soreness upon palpation, none of the participants reported any pain in the patellar tendon or other limitations when performing the prescribed exercises. Another investigator prepared the tissue for further analysis under light microscope. The tissue was kept moist in isotonic saline during the entire procedure. The biopsy was first dissected free from potential non-tendinous tissue (i.e. fat and subcutaneous tissue). Then, the investigator cut a small piece for electron microscopy with visible regular, longitudinal arrangement of the collagen fibers, which was immersed in glutaraldehyde and stored at 5 °C. The rest of the sample was immediately frozen in liquid nitrogen and subsequently stored at − 80 °C until further analysis.
Fibril morphology
Fibril morphology was analyzed with transmission electron microscopy (TEM). Preparation of the samples are described in detail in previous publications [43, 46]. Images were obtained with a Philips TM 100 transmission electron microscope at 80 kV equipped with Megaview 2 camera. A blinded (to group) technician was instructed to zoom to a magnification of 1050 (100x100μm), where the tissue microstructure was not visible, and identify two different areas with core fibrillary structure (fascicles). The technician then divided the screen visually in six fields, and imaged one area in each field at 24500 x magnification (4x4μm). In this way we obtained a total of 12 unbiased images per biopsy, which all contained core tendon tissue with clearly visible cross-sectioned collagen fibrils.
One blinded investigator performed all measurements of fibril diameters in the image analysis software ImageJ (Version 1.51n FIJI package; NIH, Bethesda, Maryland, USA). A macro was initially applied on each image to randomly generate a 300 × 300 nm2 unbiased counting frame with guard regions, and semi-automatically delineate fibril contours by best fitting ellipses. The investigator then confirmed the automated measurements within the counting frame, and manually corrected any erroneously measured fibrils. We used an elliptic fit because some fibrils were not perfectly round, and chose the minor axis diameter as the true fibril diameter to alleviate the influence of sectioning angle. The investigator counted on average 335 ± 131 fibrils for each tendon specimen (12 images). The data were reduced for each individual and time point to mean fibril diameter, volume fraction, and fibril density.
Biochemical analysis
The raw tendon samples were thawed and freeze-dried to give the tendon dry-weight. For biochemical analysis, we used on average 7.2 mg (1.9 to 17.8 mg) tissue with an average water content of 84.2%, giving 1.1 mg tissue dry weight for biochemical analysis. Afterwards, we performed gas-phase hydrolysis on the samples within sealed and evacuated tubes for 24 h at 110 °C followed by freeze-drying the samples again. The freeze-dried samples were dissolved in de-mineralized water to a concentration of 5 mg/ml.
Hydroxy-proline was determined on the hydrolyzed samples as a measure of collagen content as described in detail in a recent publication [47].
HP and LP were determined with two different ELISA kits, one made for measuring HP in serum (MicroVue Serum PYD, 8019, Quidel Corp.), and one for measuring LP in urine (MicroVue DPD, 8007, Quidel Corp.). Due to the much higher concentration of HP and LP in the tissue hydrolysates than in serum and urine, the samples (reconstituted in water at 5 mg/mL) were diluted 2000 fold for the HP analysis and 100 fold for LP, which reduces the risk of interference from compounds that would not normally be present in serum or urine. Both assays were competitive ELISAs based on a polyclonal rabbit antibody for HP and a monoclonal one for LP. The ELISAs were performed in duplicate with a CV of 6 and 14% for LP and HP, respectively. Aside from using a different type of sample the manufacturer’s instructions were followed without modification.
Total fluorescence was used as a marker of total AGE modification [48]. Fluorescence measurements were made on the hydrolysates reconstituted in water. To ensure a consistent pH and get enough volume for triplicate measurements, samples were diluted 6 fold into 0.12 M HCl (0.1 M final concentration). The diluted samples containing 83 μg tissue/ml were plated onto black 96 well plates and read on a Wallac1420 Victor microplate reader (Perkin Elmer) at 340/5 nm excitation and 460 nm (382–507 nm) emission. Since fluorescence correlated positively to collagen content (r2 = 0.52, p < 0.001), we chose to report fluorescence relative to collagen.
Statistics
Data were analyzed by repeated measures two-way ANOVA with baseline adjustment using time-point (0 mths vs 12 mths) and intervention-group (CON vs MRT vs HRT) as factors and Tukey-Kramer post-hoc tests. Normality was confirmed by visual inspection of residual plots. Signal intensity and fluorescence were not normally distributed and were log-transformed and analyzed again. All other variables were normally distributed. Outlier analysis was performed with an online Grubb’s test [49] before statistical testing (no outliers were detected). SAS statistical Software v. 9.4 (SAS Institute, USA) were used for statistical testing. Continuous variables are presented as arithmetic mean ± SE, and log-transformed values are presented as geometric mean [upper limit-lower limit]. Participant characteristics were summarized as mean ± SD.
Results
Participants
One woman dropped out of CON due to lack of time, and one woman and one man dropped out of MRT due to stroke and meningitis. The remaining 33 participants completed the study with an average training compliance of 86 ± 12%, which was not different between MRT and HRT. Their physiological characteristics are presented in Table 1.
Table 1.
Participant baseline characteristics
| Total | CON | MRT | HRT | |
|---|---|---|---|---|
| Participants | 33 | 10 | 13 | 10 |
| Sex (men/women) | 18/15 | 6/4 | 5/8 | 7/3 |
| Age (y) | 67 ± 2 | 68 ± 1.8 | 66 ± 2.4 | 67 ± 2.3 |
| Height (cm) | 173 ± 8 | 175 ± 8 | 173 ± 7 | 171 ± 8 |
| Weight (kg) | 78 ± 14 | 82 ± 16.9 | 73 ± 12 | 79 ± 14 |
| BMI (kg/m2) | 25.8 ± 3.7 | 26.6 ± 4.3 | 24.3 ± 3.5 | 26.9 ± 3.1 |
| HbA1c (mmol/l) | 36 ± 3.1 | 37 ± 3.5 | 35.2 ± 3.0 | 35.4 ± 2.9 |
| Total-C (mmol/l) | 5.9 ± 0.9 | 5.7 ± 0.8 | 6.0 ± 1.0 | 6.0 ± 0.9 |
| Training Compliance (%) | 86 ± 12 | 86 ± 16 | 86 ± 7 |
Values are mean ± SD. Baseline group-differences within studies evaluated with one-way ANOVA, chi-square test (sex distribution), and Welch’s unpaired t-test (compliance). No significant group differences
HRT heavy resistance training, MRT moderate load resistance training, CON control, BMI body mass index, Total-C total cholesterol
Muscle strength and size
Twelve months intervention significantly improved isometric quadriceps muscle strength in HRT (p < 0.01) and MRT (p < 0.05) but not in CON. There was a significant time x group interaction since HRT improved more than both CON and MRT (p < 0.05) (Fig. 2). VL-CSA was not significantly affected by training (Fig. 2).
Fig. 2.

Isometric quadriceps (Q)-strength and vastus lateralis cross-sectional area (CSA) before (0 months) and after 12 months heavy resistance training (HRT), moderate load resistance training (MRT) or no training (CON). *Significant time x group interaction based on 2-way repeated measures ANOVA (p < 0.05) a: Significantly different from 0 months (p < 0.05). b: Significantly different from CON12 (p < 0.05), and c: Significantly different from MRT12 (p < 0.05) based on Tukey-Kramer post-hoc test
Health related variables & physical activity level
Weight, BMI, total cholesterol and HbA1c displayed no time x group interactions. There was a significant main effect of time (p < 0.001) on HbA1c which was higher after the intervention in all three groups. All other health-related variables did not change significantly during the intervention. Physical activity level measured as daily step-count displayed no time x group interactions, but there was a trend towards a main effect of time with increasing step-count across groups from 8500 (0 months) to 9400 (12 months) (p = 0.07).
Patellar tendon mechanical properties
Average patellar tendon force/deformation and stress/strain curves are illustrated in Fig. 3. There were significant time x group interactions in maximal patellar tendon stiffness and Young’s modulus. Post hoc tests showed higher maximal stiffness and a tendency for higher maximal modulus (p = 0.09) with HRT compared to CON (Table 2), and maximal stiffness tended to be higher after HRT compared to MRT (p = 0.06). Maximal deformation, strain, force, and stress did not display any time x group interactions, but there was a significant main effect of time in all these variables, except for maximal stress (p = 0.07).
Fig. 3.

Illustration of average patellar tendon force/deformation (left) and stress/strain (right) curves before (PRE) and after (POST) 12 months heavy load resistance training (HRT), moderate load resistance training (MRT), or no training (CON)
Table 2.
Maximal patellar tendon mechanical properties
| CON (n = 10) | MRT (n = 13) | HRT (n = 10) | ||||
|---|---|---|---|---|---|---|
| 0 mths | 12 mths | 0 mths | 12 mths | 0 mths | 12 mths | |
| Internal moment arm (cm) | 2.75 ± 0.04 | 2.79 ± 0.04 | 2.73 ± 0.05 | |||
| Maximal deformation (mm) ‡ | 2.7 ± 0.3 | 2.9 ± 0.2 | 2.3 ± 0.2 | 2.7 ± 0.2 | 2.4 ± 0.3 | 2.5 ± 0.3 |
| Maximal force (N) ‡‡ | 4890 ± 390 | 5020 ± 330 | 4070 ± 480 | 4690 ± 490 | 5020 ± 530 | 5550 ± 470 |
| Maximal stiffness (N/mm)** | 3530 ± 490 | 3010 ± 440 | 3350 ± 300 | 3170 ± 210 | 4060 ± 430 | 4420 ± 340B |
| Maximal strain (%) ‡ | 6.0 ± 0.8 | 6.4 ± 0.5 | 5.3 ± 0.5 | 6.1 ± 0.6 | 5.5 ± 0.6 | 5.7 ± 0.5 |
| Maximal stress (MPa) | 43 ± 3 | 43 ± 2 | 33 ± 3 | 36 ± 3 | 43 ± 4 | 45 ± 3 |
| Maximal modulus(MPa)* | 1430 ± 2000 | 1200 ± 180 | 1180 ± 70 | 1090 ± 50 | 1510 ± 150 | 1560 ± 140 |
Values are means ±SE. Data analyzed by repeated measures two-way ANOVA with baseline adjustment and Tukey-Kramer post hoc tests
CON control, MRT moderate load resistance training, HRT heavy resistance training
Significant interaction denoted by *(p < 0.05) or **(p < 0.01). Main effect of time denoted by ‡(p < 0.05) or ‡‡(p < 0.01). Post hoc tests: B: significant difference from CON12 (p < 0.05)
Common force mechanical properties also displayed a significant time x group interaction in patellar tendon stiffness and a strong trend towards interaction in Young’s modulus (p = 0.05) (Fig. 4). Common force stiffness was significantly higher after HRT compared to both CON and MRT, which reduced this variable. Common force Young’s modulus further tended to be higher after HRT compared to MRT (p = 0.07), and MRT significantly reduced modulus (Fig. 4). Common force deformation and strain displayed no time x group interactions or main effects of time.
Fig. 4.

Common force patellar tendon (PT) stiffness and Young’s modulus before (0 months) and after 12 months heavy resistance training (HRT), moderate load resistance training (MRT) or no training (CON). Bars represent mean ± SE. Significant time x group interaction denoted by **(p < 0.01) or *(p = 0.05) based on repeated measures 2-way ANOVA. A: Significantly different from 0 months (p < 0.05). B: Significantly different from CON12 (p < 0.05), C: Significantly different from MRT12 (p < 0.05) based on Tukey-Kramer post-hoc test
Patellar tendon MRI
There was a trend towards a time x group interaction of total patellar tendon CSA (p = 0.07), which increased significantly over time in HRT and MRT but not in CON (Fig. 5). The proximal tendon region (CSA prox) showed a significant time x group interaction, and post-hoc test revealed increase over time in MRT and a similar, although not significant, increase in HRT (p = 0.25) (Fig. 5). The mid and distal tendon regions did not display any time x group interactions, but there was a main effect of time in both regions where CSA increased from 0 to 12 months across groups.
Fig. 5.

Changes in patellar tendon cross-sectional area (CSA) in three different regions (proximal, mid, distal) and in total after 12 months heavy resistance training (HRT), moderate load resistance training (MRT) or no training (CON). Bars represent mean ± SE. *Significant time x group interaction based on repeated measures ANOVA (pz0.05). ‡: Main effect of time (p < 0.05). a: Significant change from 0 to 12 months (p < 0.05), b: Significantly different from CON (p < 0.05), c: Significantly different from MRT (p < 0.05) based on Tukey-Kramer post-hoc test
Collagen fibril morphology
There were no significant time x group interactions in volume fraction, fibril diameter or fibril density (Table 3). There was a main effect of time in fibril diameter, which decreased over time, and fibril density which increased over time.
Table 3.
Collagen fibril morphology
| CON | MRT | HRT | ||||
|---|---|---|---|---|---|---|
| 0 mths (n = 9)a | 12 mths (n = 9)a | 0 mths (n = 12)a | 12 mths (n = 12)a | 0 mths (n = 9)a | 12 mths (n = 9)a | |
| Volume Fraction (%) | 63 ± 2 | 59 ± 1 | 59 ± 2 | 62 ± 1 | 59 ± 2 | 59 ± 2 |
| Mean fibril diameter (nm)‡ | 91 ± 5 | 83 ± 4 | 87 ± 3 | 86 ± 5 | 89 ± 4 | 78 ± 3 |
| Density (#/μm2)‡ | 27 ± 2 | 31 ± 3 | 28 ± 2 | 33 ± 3 | 28 ± 3 | 36 ± 3 |
Values are means ±SE. Data analyzed by repeated measures 2-way ANOVA with baseline adjustment and Tukey-Kramer post-hoc test
HRT heavy resistance training, MRT moderate load resistance training, CON no training
Main effect of time denoted by ‡(p < 0.05)
aOne value missing due to technical problems
Collagen and collagen x-links
There were no significant time x group interactions in tendon collagen content, enzymatic cross-links, or fluorescence (Table 4). Tendon fluorescence displayed a significant main effect of time (Fig. 6).
Table 4.
Collagen and collagen cross-links
| CON | MRT | HRT | ||||
|---|---|---|---|---|---|---|
| 0 mths (n = 10) | 12 mths (n = 10) | 0 mths (n = 13) | 12 mths (n = 13) | 0 mths (n = 9)a | 12 mths (n = 9)a | |
| Collagen (%) | 69 ± 3 | 62 ± 2 | 61 ± 4 | 59 ± 2 | 63 ± 3 | 61 ± 2 |
| HP (mmol/mol collagen)b | 298 ± 28 | 322 ± 25 | 314 ± 23 | 267 ± 21 | 254 ± 28 | 292 ± 22 |
| LP (mmol/mol collagen)b | 24.8 ± 5.0 | 26.5 ± 3.8 | 40.8 ± 4.4 | 28.8 ± 5.1 | 37.0 ± 5.9 | 34.3 ± 3.2 |
| Fluorescence (AU/mol collagen)‡ | 1139 [1096–1185] | 1235 [1176–1297] | 1155 [1124–1187] | 1196 [1161–1232] | 1263 [1191–1339] | 1328 [1285–1372] |
Values are means ±SE or geometric mean [lower limit – upper limit]. Data analyzed by repeated measures two-way ANOVA with baseline adjustment and Tukey-Kramer post hoc test
CON = Control, MRT = moderate load resistance training, HRT = Heavy load resistance training
No time x group interactions. ‡: Main effect of time (p < 0.05)
aOne missing sample due to anti-coagulant therapy. bFive missing pairs of samples due to logistics
CON: n = 9, MRT: n = 11, HRT: n = 8
Fig. 6.
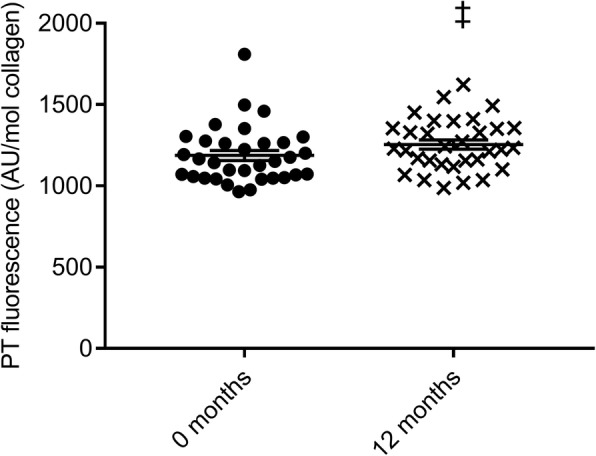
Fluorescence of patellar tendon (PT) biopsies as a marker of non-enzymatic (AGE) cross-links before (0 months) and after 12 months intervention. ‡: Main effect of time (p < 0.05) based on repeated measures two-way ANOVA with baseline adjustment. AU = arbitrary units
Discussion
The present investigation compared the effects of high or moderate load long-term resistance training on patellar tendon mechanical properties, macro- and microscopic morphology, and collagen cross-links in older adults. The main findings were that adaptation of patellar tendon mechanical properties over the 12 months intervention was dependent on magnitude of training load, whereas tendon CSA increased after both heavy and moderate load resistance training. Contrary to our hypothesis, the load-dependent changes in tendon mechanical properties were unrelated to any changes of enzymatic cross-links or advanced glycation end-products.
Strength training improved maximal isometric quadriceps strength (IsoMVC) with heavy load training (+ 21%) being superior to moderate load (+ 8%) or no training (7%) (Fig. 2). The superior effect of heavy compared to moderate load training on muscle strength is consistent with current knowledge [50] although the strength increase in the present study was slightly lower than previously reported in a meta-analysis of strength training studies [50]. Vastus lateralis CSA was surprisingly not affected by training, suggesting that either there was a quite large neural adaptation to training or that muscle hypertrophy mainly occurred in the other components of the quadriceps muscle. The relatively high training compliance of 86% in both groups could not explain the blunted muscular response to training. The training load and volume in the present investigation was comparable to previous investigations [50], but we cannot exclude that the prescribed training load and volume was compromised for some participants due to discomfort or lack of motivation. Another possibility is that the population of older adults in the present study had a higher fitness level to begin with than in previous investigations, which is supported by a relatively high habitual physical activity level of ∼8500 daily step-counts.
Patellar tendon mechanical properties
Tendon stiffness calculated at common force increased by 6% after HRT, and decreased by 10 and 7.5% after MRT and CON respectively. Young’s modulus remained unchanged after HRT but decreased by 14% after MRT and 9% after CON. The strong trend towards different tendon responses in the three groups (p = 0.05) may suggest an age-dependent decrease of tendon mechanical and material properties, which was ameliorated by HRT but not MRT (Fig. 4). The fact, that tendon modulus and not only tendon stiffness was affected by training suggest that there was a change in material quality and not only amount of tissue. Although reduced stiffness and modulus over time in the CON group may be plausible, unchanged stiffness after 12 months heavy load training combined with relatively small differences between the three groups is somewhat surprising since previous studies have reported ∼20–60% increases of patellar tendon modulus after only three months heavy load training in older adults [9, 11, 13, 51]. The modest tendon adaptation however fits well with the modest muscular adaptation to training, since we would expect the muscle-tendon unit to adapt synchronously [52]. Although HRT did not increase tendon modulus, our findings support previous studies that have found superior effects of heavy compared to light load training on tendon mechanical properties in young and middle aged [12, 53] as well as older adults [11]. The novelty of the present finding is that moderate load training with an approximately comparable volume as heavy load training was also insufficient to affect tendon mechanical properties. Our results thus confirm that a certain loading threshold needs to be surpassed in order for tendons to adapt [54, 55]. In fact, MRT seemed to reduce tendon stiffness, which contradicted our own hypothesis as well as a previous study showing decreased strain of the vastus lateralis aponeurosis after low load training [4]. As for the CON group, MRT may have provided an insufficient mechanical stimulus on the patellar tendon to counteract an age-related decrease of stiffness over the 12 months intervention.
Several previous studies have investigated the effects of short-term (3 months) resistance training on tendon mechanical properties and found either increased [9–11, 51] or unchanged [13] tendon stiffness. Only few studies have investigated the effect of long-term training and found no additional effects after 1.5 [10] or 4 years [14] compared to three months training. We cannot exclude that the improvements in tendon stiffness and modulus measured after 12 months training were already present after the first three months. In another recent randomized training study in our department (unpublished data) including older adults with an average age similar to the ones in the present study, and using the same test-protocol, we found no significant differences in tendon mechanical properties after 3 months heavy or light load resistance training. Although many previous studies have found significant increases of patellar tendon stiffness and modulus after short-term resistance training in older adults [9, 11, 51], our own data suggest that longer training periods are superior. A more protracted mechanical adaptation to training would also fit with the relatively slow turnover of tendon extracellular matrix.
Taken together, our results show a more blunted response of the muscle-tendon unit to training compared to previous studies but confirm that heavy load training is superior with regards to inducing favorable mechanical and material adaptations of the patellar tendon. Maintenance of tendon stiffness may be critical to optimal muscle function [5, 56] and postural balance [6] in old age, and our results therefore support that heavy load resistance training should be incorporated in training programs in this age group.
Patellar tendon morphology
Patellar tendon CSA (total) tended to be affected by intervention group (p = 0.07) with increases after both HRT (+ 6%) and MRT (+ 5%), but not CON (+ 1%) (Fig. 5). The possible increase in CSA makes the tendon able to support higher loads without imposing more stress on the tendon, and thus protects the tissue from damage. To our surprise, the effect of training on tendon CSA was not dependent on load magnitude since MRT in contrast to mechanical properties increased tendon CSA equally much as HRT (Fig. 5). The tendon tended to grow without any muscle hypertrophy, although myofibrillar protein synthesis is more sensitive to loading than tendon collagen synthesis [57]. Although somewhat surprising after a resistance training intervention, tendon growth is possible without a hypertrophic muscle stimulus as observed after life-long endurance running [8]. Since we only measured vastus lateralis CSA in the present investigation it cannot be ruled out that hypertrophy occurred in the remaining three muscle compartments of the quadriceps.
Although several short-term training studies have not been able to show any effect of heavy or light load on tendon CSA in older adults [9, 11, 51, 58], our longitudinal data supports some [8, 15], albeit not all [14] previous human cross-sectional data, which show that long-term (several years) habitual loading of both relatively low [8] and high load [15] is associated with higher patellar tendon CSA. One recent longitudinal study did, however, not find additional effect of 1.5 years compared to three months heavy resistance training on Achilles tendon CSA [10], and it is possible that specific tendons have unique time-courses of adaptation to loading. The increments of patellar tendon CSA tended to be region-specific with significant training effects in the proximal region, but not in the mid and distal regions (Fig. 5). A previous study in young men found similar region-specific increments of the patellar tendon CSA with 7% higher proximal CSA after light load training and 6 and 4% higher proximal and distal CSA respectively after high load training [59]. The increased CSA in that study was accompanied by increased tendon stiffness in the heavy load group only, which corroborates our own data. Local stress concentration in the patellar tendon during loading may explain the regional adaptations. Similar regional adaptations following strength training do not seem to take place in the Achilles tendon in older adults [10].
Increased CSA with training may be mediated by addition of new collagen to existing fibrils, new fibrils, increased intrafibrillary spacing between collagen molecules, or more interfibrillary material such as fat, water, or proteoglycans. In the present investigation we measured core tendon collagen content, fibril volume fraction, as well as fibril diameter and density but none of these variables were affected by training (Table 3). The lack of training effect on collagen content and fibril morphology was not surprising, since it has recently been demonstrated convincingly that no or very little renewal of collagen takes place in the core tendon after teenage years [19]. Moreover, it is also consistent with previous human cross-sectional data on master athletes compared to sedentary age-matched controls [8]. However, it is possible that adaptations at the fibrillary level have gone undetected in our core tendon biopsies, since new collagen may have been added to the peripheral region of the tendon [17] or to the most proximal region, which would be consistent with the region-specific adaptations in tendon CSA. Region-specificity could thus explain the disparity between adaptations at the fibrillary and whole tendon level. Training induced accumulation of water could also increase tendon CSA, but this explanation seems unlikely since water accumulation into the matrix would theoretically reduce fibril volume fraction which was unaffected by training.
In contrast to the changes in mechanical properties, we found that region specific patellar tendon hypertrophy occured independent of load magnitude. This could however not be explained by changes in the ultrastructural morphology of the tendon core which was unaffected by training.
Collagen cross-links
Contrary to our hypothesis, the enzymatic cross-links HP and LP were unaffected by 12 months resistance training (Table 4). Previous human data have also shown no difference in enzymatic cross-links between endurance trained and sedentary older adults [8], and no effect of short-term resistance training in young adults [60]. Our measurements of LP in tendon were similar in magnitude to that found by others [20, 27, 30], but it should be acknowledged that our HP results were somewhat lower than that found in other studies [20, 27, 30, 60]. It cannot be excluded that our measurements may have underestimated the HP content, but importantly there was no change in determined HP after exercise. Our results thus confirm that enzymatic cross-links do not mediate moderate or high load training induced changes in tendon stiffness or Young’s modulus despite long training duration. Since enzymatic cross-links are essential to development of normal force transmission within the tendon [3, 20, 61], we also analyzed the relationship between tendon stiffness or modulus and HP or LP, at baseline as well as in the changes over time. The analysis showed no correlations between enzymatic cross-links and tendon mechanical properties in our group of older adults, which corroborate previous human and animal studies [30, 62, 63] and suggests that enzymatic cross-links play a limited role in the adaptation of tendon biomechanics after maturity. It is also plausible that enzymatic cross-links after maturity are only formed in the peripheral regions of the tendon, where there is also evidence of collagen synthesis in response to exercise [17], and future research should investigate region-specific molecular adaptations to training.
Fluorescence which was used as a marker of AGE content increased by ~ 6% over the 12 months across intervention-groups (Fig. 6), but was unaffected by both moderate and heavy load resistance training. The time dependent increase of fluorescence was not surprising given the nature of the glycation process, which results in accumulation of AGEs over the life-course [1]. It was however somewhat surprising that 12 months training could not attenuate AGE accumulation since others have found decreased tendon AGE-levels after short-term endurance training in old mice [21], attenuated accumulation of the AGE-marker pentosidine after life-long endurance training [8], and reduced pentosidine after short-term heavy load resistance training in young men with patellar tendinopathy [27]. Even longer training duration may be necessary for healthy old humans to significantly attenuate AGE-accumulation.
To support that fluorescence measurement provided a truthful depiction of AGE-accumulation, we performed correlational analysis between age and fluorescence (Fig. 7). As expected, fluorescence correlated positively to age even within the relatively small age-range in our population (63–71 years). Interestingly, HbA1c also increased by ~ 3% or ~ 1 mmol/mol (p < 0.0001) without any difference between groups (Table 1). Increased HbA1c confirms the time-dependent accumulation of glycation products, but it may also be due to variation between PRE and POST measurements since the increase was way higher than what other large-scale studies on healthy individuals have found (~ 1 mmol/mol over 10 years) [64]. There was no relationship between tendon fluorescence and HbA1c either at baseline or in the changes over time, indicating different time-patterns of erythrocyte and tendon glycation.
Fig. 7.
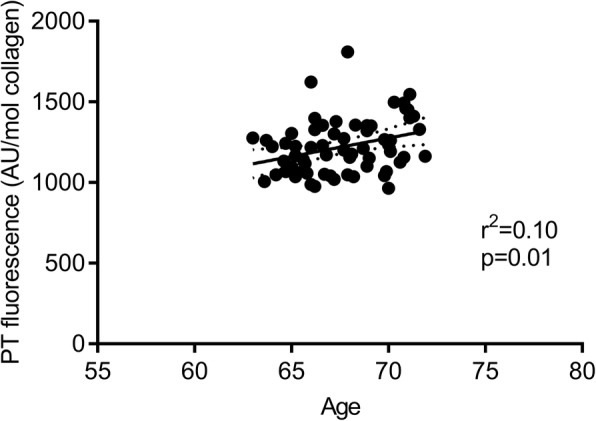
Correlation between age and patellar tendon (PT) fluorescence using bot PRE and POST values as determined by Pearson’s correlation coefficient. Dotted lines are 95% confidence bands of the best fit
We further examined the relationship between fluorescence and Young’s modulus since accumulation of AGEs may affect tendon material properties [3, 65], but there was no correlation between these variables either at baseline or in the changes over time. Previous in vitro studies have shown associations between AGEs and tendon mechanical properties [21, 26, 28], but human studies have in line with our results not been able to associate tendon AGEs and in vivo patellar tendon mechanical properties [30, 62], suggesting that other matrix components (i.e. proteoglycans, glycoproteins) contribute more to in vivo mechanical phenotype in old humans. AGEs comprise a large and heterogeneous family of chemical compounds, and it is possible that training induced changes of specific cross-linking AGEs have “drowned” in the measurement of total fluorescence. Further, region-specific changes in AGEs may have occurred in the outer layers or most proximal and distal tendon regions, without being detected in our core tendon samples. Finally, it is possible that AGEs mainly affect tissue mechanics in the failure region [47, 66, 67], whereas in vivo tendon testing only considers mechanical properties in the physiological range of the force-elongation relationship. It must be noted that recent studies have shown that the deformation mechanism and strength of the tendon are greatly affected by the presence of cross-links [68], and that multiscale mechanical analysis of in vitro glycated tendons strongly suggests that AGEs reduce tissue viscoelasticity by severely limiting fiber–fiber and fibril–fibril sliding [69].
Tendon fluorescence increased as expected over time and this may very well be due to age-related accumulation of AGEs. To prevent tendon AGE accumulation, training should be of longer duration than 12 months or probably initiated at younger age. The relation between patellar tendon AGEs and in vivo mechanical properties remains elusive.
Limitations
In contrast to young women [70, 71], older postmenopausal women seem to have indistinguishable tendon mechanical properties from men [72]. Both men and women were included in the present study to make the results more generally applicable. Although not statistically significant, more men were randomized to HRT and more women were randomized to MRT. To test the potential influence of sex on the training effect, we performed a secondary 2-way ANOVA on the changes over time with sex and intervention-group as factors. The analysis did not show any interactions between sex and intervention-group or main effects of sex in muscle or tendon variables, suggesting that older men and women respond similarly to resistance training. This makes it unlikely that the unequal sex-distribution explained the group-differences in training adaptations.
Despite a high average training compliance of 86% in both MRT and HRT, the range was quite large (51 to 100%). We therefore made a secondary per protocol statistical analysis excluding participants with a compliance < 80% and also excluding one CON who admitted to have initiated strenuous training activities during the intervention. The remaining 7 HRT (compliance = 88 ± 5), 9 MRT (compliance = 94 ± 5), and 9 CON did not display appreciably different results than the primary statistical analysis and did consequently not affect the reached conclusions.
The duration of the ramped contractions was fixed at 8 s in the present investigation, which means that individuals with increased maximal muscle strength had a higher rate of force development. Higher rate of force development has been associated with increased tendon stiffness but this has mainly been shown at even higher rates of force development (i.e. 50 Nm/s (~ 3 s.) to 110 Nm/s (~ 1.5 s) [73] than in the present investigation. We therefore find it less likely that the group differences in maximal strength could explain the observed group differences in stiffness after the interventions.
Antagonist (hamstring) co-contraction may amount to 10–30% of the resultant knee-joint moment during isometric knee-extensions as measured by electromyography (EMG) [74]. In the present investigation we however chose not to consider this potential contribution to the knee-joint moment because it would add an additional level of technical variation to the estimation of tendon force, which in our experience (unpublished data) is even larger in untrained individuals due to EMG cross-talk (overestimated co-contraction due to quadriceps signal).
The present investigation was a nested study where the training interventions and study population were originally determined to investigate the impact of different training regimens on muscle function and general health in a representative elderly population. Although the study design may not have been ideally suited to answer the question about the specific influence of training load on tendons (i.e. lack of training volume matching, heterogeneity of the participants), our results still support that a training program designed to generally enhance muscle function and health in an elderly population also affected connective tissue function.
Conclusion
The present study demonstrated load-dependent adaptations of patellar tendon mechanical properties with heavy load training being superior to moderate load or no training. However, patellar tendon cross-sectional area increased after training independent of load magnitude. The observed adaptations to resistance training could not be related to any changes in collagen content, fibril morphology, enzymatic cross-links, or advanced glycation end-products in the tendon core. We suggest that heavy load resistance training over longer periods is advisable for older adults to maintain tendon function with aging.
Acknowledgements
We appreciate the assistance of Christian Orhammer Josefsen and Mads Vang Rasmussen for assistance with data analysis. We thank PROcare and Theraband for kindly providing the elastic rubber bands used for MRT.
Funding
This work was supported by the Lundbeck Foundation, the Novo Nordisk Foundation, the Danish Council for Independent Research (Health and Disease), and the Nordea Foundation (Center for Healthy Ageing grant). PROcare ApS and Theraband® sponsored the elastic rubber bands. The funding bodies had no influence on the design of the study or in the collection, analysis, or interpretation of the data.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- 1RM
One repetition maximum
- AGE(s)
Advanced glycation end-product(s)
- ANOVA
Analysis of variance
- BMI
Body mass index
- CON
Control group
- CSA
Cross sectional area
- CV
Coefficient of variation
- EMG
Electromyography
- HP
Hydroxylysylpyrridinoline
- HRT
Heavy load resistance training
- IsoMVC
Maximal voluntary isometric knee-extensor strength
- LOX
Lysyl oxidase
- LP
Lysylpyrridinoline
- MRI
Magnetic resonance imaging
- MRT
Moderate load resistance training
- NIH
National Institute of Health
- POST
After intervention (12 months)
- PRE
Before intervention (0 months)
- TEM
Transmission Electron Microscopy
- Total-C
Total serum cholesterol
- US
Ultrasound
- VL-CSA
vastus lateralis cross-sectional area
Authors’ contributions
CSE was involved in designing, planning, and organizing the study, made significant contributions to data acquisition and analysis, and drafted the manuscript. AG and RBS made significant contributions to data acquisition and analysis and critically revised the manuscript for intellectual content. CC, MK, and SPM designed and planned the study and critically revised the manuscript for intellectual content. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The study complied with the Helsinki Declaration and all participants gave oral and written consent before enrollment. The study was registered at clinicaltrials.gov (NCT02123641, registered on April 25th 2014) and was approved by the Capital Region ethical committee (J-no. H-3- 2014-017) and the Danish Data Protection Agency (J-no. 2012-58-0004 - BFH-2016-025, I-suite 04545).
Consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Christian S. Eriksen, Email: Eriksen.christian@gmail.com
Rene B. Svensson, Email: svensson.nano@gmail.com
Anne T. Gylling, Email: anne.theil.gylling@regionh.dk
Christian Couppé, Email: ccouppe@gmail.com.
S. Peter Magnusson, Email: spmagnusson@gmail.com.
Michael Kjaer, Email: michaelkjaer@sund.ku.dk.
References
- 1.Bailey AJ, Paul RG, Knott L. Mechanisms of maturation and ageing of collagen. Mech Ageing Dev. 1998;106:1–2):1–56. doi: 10.1016/S0047-6374(98)00119-5. [DOI] [PubMed] [Google Scholar]
- 2.Brownlee M. Advanced protein glycosylation in diabetes and aging. Annu Rev Med. 1995;46:223–234. doi: 10.1146/annurev.med.46.1.223. [DOI] [PubMed] [Google Scholar]
- 3.Avery NC, Bailey AJ. The effects of the Maillard reaction on the physical properties and cell interactions of collagen. Pathol Biol [Internet] 2006;54(7):387–395. doi: 10.1016/j.patbio.2006.07.005. [DOI] [PubMed] [Google Scholar]
- 4.Kubo K, Kanehisa H, Miyatani M, Tachi M, Fukunaga T. Effect of low-load resistance training on the tendon properties in middle-aged and elderly women. Acta Physiol Scand. 2003;178(1):25–32. doi: 10.1046/j.1365-201X.2003.01097.x. [DOI] [PubMed] [Google Scholar]
- 5.Bojsen-Møller J, Magnusson SP, Rasmussen LR, Kjaer M, Aagaard P. Muscle Perfomance during maximal isometric and dynamic contractions is influenced by the stiffness of the tendinous structures. J Appl Physiol. 2005;99(3):986–994. doi: 10.1152/japplphysiol.01305.2004. [DOI] [PubMed] [Google Scholar]
- 6.Onambele GL, Narici MV, Maganaris CN. Calf muscle-tendon properties and postural balance in old age Calf muscle-tendon properties and postural balance in old age. J Appl Physiol. 2006:2048–56. [DOI] [PubMed]
- 7.Smith RKW, Birch HL, Goodman S, Heinegård D, Goodship AE. The influence of ageing and exercise on tendon growth and degeneration - hypotheses for the initiation and prevention of strain-induced tendinopathies. Comp Biochem Physiol - Part A. 2002;133(4):1039–1050. doi: 10.1016/S1095-6433(02)00148-4. [DOI] [PubMed] [Google Scholar]
- 8.Couppé C, Svensson RB, Grosset JF, Kovanen V, Nielsen RH, Olsen MR, et al. Life-long endurance running is associated with reduced glycation and mechanical stress in connective tissue. Age (Omaha). 2014;36(4). [DOI] [PMC free article] [PubMed]
- 9.Reeves ND, Maganaris CN, Narici M V. Effect of strength training on human patella tendon mechanical properties of older individuals. J Physiol [Internet]. 2003;548(Pt 3):971–81. [DOI] [PMC free article] [PubMed]
- 10.Epro G, Mierau A, Doerner J, Luetkens JA, Scheef L, Kukuk GM, et al. The Achilles tendon is mechanosensitive in older adults: adaptations following 14 weeks versus 1.5 years of cyclic strain exercise. J Exp Biol [Internet] 2017;220(6):1008–1018. doi: 10.1242/jeb.146407. [DOI] [PubMed] [Google Scholar]
- 11.Grosset JF, Breen L, Stewart CE, Burgess KE, Onambélé GL. Influence of exercise intensity on training-induced tendon mechanical properties changes in older individuals. Age (Omaha). 2014;36(3):9657. doi: 10.1007/s11357-014-9657-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bohm S, Mersmann F, Arampatzis A. Human tendon adaptation in response to mechanical loading: a systematic review and meta-analysis of exercise intervention studies on healthy adults. Sport Med - Open [Internet]. 2015;1(7). Available from: Missingcontent/1/1/7. [DOI] [PMC free article] [PubMed]
- 13.Carroll CC, Dickinson JM, LeMoine JK, Haus JM, Weinheimer EM, Hollon CJ, et al. Influence of acetaminophen and ibuprofen on in vivo patellar tendon adaptations to knee extensor resistance exercise in older adults. J Appl Physiol. 2011;111(2):508–515. doi: 10.1152/japplphysiol.01348.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Massey GJ, Balshaw TG, Maden-Wilkinson TM, Folland JP. Tendinous tissue properties after short- and long-term functional overload: Differences between controls, 12 weeks and 4 years of resistance training. Acta Physiol [Internet]. 2018;(July 2017):e13019. [DOI] [PubMed]
- 15.Couppe C, Kongsgaard M, Aagaard P, Hansen P, Bojsen-Moller J, Kjaer M, et al. Habitual loading results in tendon hypertrophy and increased stiffness of the human patellar tendon. J Appl Physiol [Internet] 2008;105(3):805–810. doi: 10.1152/japplphysiol.90361.2008. [DOI] [PubMed] [Google Scholar]
- 16.Heinemeier KM, Olesen JL, Haddad F, Langberg H, Kjaer M, Baldwin KM, et al. Expression of collagen and related growth factors in rat tendon and skeletal muscle in response to specific contraction types. J Physiol [Internet] 2007;582(Pt 3):1303–1316. doi: 10.1113/jphysiol.2007.127639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Langberg H, Skovgaard D, Petersen LJ, Bulow J, Kjaer M. Type I collagen synthesis and degradation in peritendinous tissue after exercise determined by microdialysis in humans. J Physiol [Internet]. 1999;521(1):299–306. [DOI] [PMC free article] [PubMed]
- 18.Lenskjold A, Kongsgaard M, Larsen JO, Nielsen RH, Kovanen V, Aagaard P, et al. The influence of physical activity during youth on structural and functional properties of the Achilles tendon. Scand J Med Sci Sport. 2015;25(1):25–31. doi: 10.1111/sms.12143. [DOI] [PubMed] [Google Scholar]
- 19.Heinemeier KM, Schjerling P, Heinemeier J, Magnusson SP, Kjaer M. Lack of tissue renewal in human adult Achilles tendon is revealed by nuclear bomb 14C. FASEB J. 2013;27(5):2074–2079. doi: 10.1096/fj.12-225599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Eyre DR, M a P, Gallop PM. Cross-linking in collagen and elastin. Annu Rev Biochem. 1984;53:717–748. doi: 10.1146/annurev.bi.53.070184.003441. [DOI] [PubMed] [Google Scholar]
- 21.Wood LK, Brooks SV. Ten weeks of treadmill running decreases stiffness and increases collagen turnover in tendons of old mice. J Orthop Res. 2016;34(2):346–353. doi: 10.1002/jor.22824. [DOI] [PubMed] [Google Scholar]
- 22.Bailey AJ. Molecular mechanisms of ageing in connective tissues. Mech Ageing Dev. 2001;122(7):735–755. doi: 10.1016/S0047-6374(01)00225-1. [DOI] [PubMed] [Google Scholar]
- 23.Bank RA. TeKoppele JM, Oostingh G, Hazleman BL, Riley GP. Lysylhydroxylation and non-reducible crosslinking of human supraspinatus tendon collagen: changes with age and in chronic rotator cuff tendinitis. Ann Rheum Dis [Internet] 1999;58(1):35–41. doi: 10.1136/ard.58.1.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Avery NC, Bailey AJ. Enzymic and non-enzymic cross-linking mechanisms in relation to turnover of collagen: relevance to aging and exercise. Scand J Med Sci Sport. 2005;15(4):231–240. doi: 10.1111/j.1600-0838.2005.00464.x. [DOI] [PubMed] [Google Scholar]
- 25.Fessel G, Li Y, Diederich V, Guizar-Sicairos M, Schneider P, Sell DR, et al. Advanced glycation end-products reduce collagen molecular sliding to affect collagen fibril damage mechanisms but not stiffness. PLoS One. 2014;9(11). [DOI] [PMC free article] [PubMed]
- 26.Sell DR, Monnier VM. Age-related association of tail tendon break time with tissue pentosidine in DBA/2 vs C57BL/6 mice: the effect of dietary restriction. J Gerontol A Biol Sci Med Sci. 1997;52(5):B277–B284. doi: 10.1093/gerona/52A.5.B277. [DOI] [PubMed] [Google Scholar]
- 27.Kongsgaard M, Kovanen V, Aagaard P, Doessing S, Hansen P, Laursen AH, et al. Corticosteroid injections, eccentric decline squat training and heavy slow resistance training in patellar tendinopathy. Scand J Med Sci Sport. 2009;19(6):790–802. doi: 10.1111/j.1600-0838.2009.00949.x. [DOI] [PubMed] [Google Scholar]
- 28.Reddy GK. Cross-linking in collagen by nonenzymatic glycation increases the matrix stiffness in rabbit Achilles tendon. Exp Diabesity Res. 2004;5(2):143–153. doi: 10.1080/15438600490277860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Naresh MD, Brodsky B. X-ray diffraction studies on human tendon show age-related changes in collagen packing. Biochim Biophys Acta [Internet] 1992;1122(2):161–166. doi: 10.1016/0167-4838(92)90319-9. [DOI] [PubMed] [Google Scholar]
- 30.Couppé C, Hansen P, Kongsgaard M, Kovanen V, Suetta C, Aagaard P, et al. Mechanical properties and collagen cross-linking of the patellar tendon in old and young men. J Appl Physiol 2009;107(June 2009):880–6. [DOI] [PubMed]
- 31.Eriksen CS, Garde E, Reislev NL, Wimmelmann CL, Bieler T, Ziegler AK, et al. Physical activity as intervention for age-related loss of muscle mass and function: protocol for a randomised controlled trial (the LISA study). BMJ Open [Internet]. 2016;6(12):e012951. [DOI] [PMC free article] [PubMed]
- 32.Volper BD, Huynh RT, Arthur KA, Noone J, Gordon BD, Zacherle EW, et al. Influence of acute and chronic streptozotocin-induced diabetes on the rat tendon extracellular matrix and mechanical properties. Am J Physiol Regul Integr Comp Physiol. 2015;309(9):R1135–R1143. doi: 10.1152/ajpregu.00189.2015. [DOI] [PubMed] [Google Scholar]
- 33.Couppé C, Svensson RB, Kongsgaard M, Kovanen V, Grosset J-F, Snorgaard O, et al. Human Achilles tendon glycation and function in diabetes. J Appl Physiol [Internet]. 2016;120(2):130–137. Available from: 10.1152/japplphysiol.00547.2015 [DOI] [PubMed]
- 34.Beason DP, Hsu JE, Marshall SM, McDaniel AL, Temel RE, Abboud JA, et al. Hypercholesterolemia increases supraspinatus tendon stiffness and elastic modulus across multiple species. J Shoulder Elb Surg [Internet]. 2013;22(5):681–686. Available from: 10.1016/j.jse.2012.07.008 [DOI] [PMC free article] [PubMed]
- 35.Hansen M, Couppe C, Hansen CSE, Skovgaard D, Kovanen V, Larsen JO, et al. Impact of oral contraceptive use and menstrual phases on patellar tendon morphology, biochemical composition, and biomechanical properties in female athletes. J Appl Physiol [Internet]. 2013;114(8):998–1008. [DOI] [PubMed]
- 36.Dahlgren G, Carlsson D, Moorhead A, Häger-Ross C, McDonough SM. Test-retest reliability of step counts with the ActivPAL device in common daily activities. Gait Posture. 2010;32(3):386–390. doi: 10.1016/j.gaitpost.2010.06.022. [DOI] [PubMed] [Google Scholar]
- 37.Hansen P, Bojsen-Moller J, Aagaard P, Kjaer M, Magnusson SP. Mechanical properties of the human patellar tendon, in vivo. Clin Biomech. 2006;21(1):54–58. doi: 10.1016/j.clinbiomech.2005.07.008. [DOI] [PubMed] [Google Scholar]
- 38.Magnusson SP, Hansen P, Aagaard P, Brønd J, Dyhre-Poulsen P, Bojsen-Moller J, et al. Differential strain patterns of the human gastrocnemius aponeurosis and free tendon, in vivo. Acta Physiol Scand. 2003;177(2):185–195. doi: 10.1046/j.1365-201X.2003.01048.x. [DOI] [PubMed] [Google Scholar]
- 39.Visser JJ, Hoogkamer JE, Bobbert MF, Huijing PA. Length and moment arm of human leg muscles as a function of knee and hip-joint angles. Eur J Appl Physiol. 1990;61(5–6):453–460. doi: 10.1007/BF00236067. [DOI] [PubMed] [Google Scholar]
- 40.Couppé C, Svensson RB, Sødring-Elbrønd V, Hansen P, Kjær M, Magnusson SP. Accuracy of MRI technique in measuring tendon cross-sectional area. Clin Physiol Funct Imaging. 2014;34(3):237–241. doi: 10.1111/cpf.12086. [DOI] [PubMed] [Google Scholar]
- 41.Boesen AP, Dideriksen K, Couppé C, Magnusson SP, Schjerling P, Boesen M, et al. Tendon and skeletal muscle matrix gene expression and functional responses to immobilisation and rehabilitation in young males: effect of growth hormone administration. J Physiol 2013;591.23(591):6039–6052. [DOI] [PMC free article] [PubMed]
- 42.Shalabi A, Kristoffersen-Wiberg M, Aspelin P, Movin T. Immediate achilles tendon response after strength training evaluated by MRI. Med Sci Sports Exerc. 2004;36(11):1841–1846. doi: 10.1249/01.MSS.0000145450.75035.61. [DOI] [PubMed] [Google Scholar]
- 43.Kongsgaard M, Qvortrup K, Larsen J, Aagaard P, Doessing S, Hansen P, et al. Fibril morphology and tendon mechanical properties in patellar tendinopathy: effects of heavy slow resistance training. Am J Sports Med. 2010;38(4):749–756. doi: 10.1177/0363546509350915. [DOI] [PubMed] [Google Scholar]
- 44.Yeung CYC, Gossan N, Lu Y, Hughes A, Hensman JJ, Bayer ML, et al. Gremlin-2 is a BMP antagonist that is regulated by the circadian clock. Sci Rep. 2014;4:1–8. doi: 10.1038/srep05183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Heinemeier KM, Lorentzen MP, Jensen JK, Schjerling P, Seynnes O, Narici M V, et alLocal trauma in human patellar tendon leads to widespread changes in the tendon gene expression. J Appl Physiol [Internet]. 2016;120:1000–10. [DOI] [PubMed]
- 46.Boesen AP, Dideriksen K, Couppe C, Magnusson SP, Schjerling P, Boesen M, et al. Effect of growth hormone on aging connective tissue in muscle and tendon: gene expression, morphology, and function following immobilization and rehabilitation. J Appl Physiol [Internet]. 2014;116(2):192–203. [DOI] [PubMed]
- 47.Svensson RB, Smith ST, Moyer PJ, Magnusson SP. Acta Biomater Effects of maturation and advanced glycation on tensile mechanics of collagen fibrils from rat tail and Achilles tendons. Acta Biomater [Internet]. 2018. Available from: 10.1016/j.actbio.2018.02.005. [DOI] [PubMed]
- 48.Monnier VM, Vishwanath V, KE F, CA E, P D RRK. Relation between complications of type I diabetes mellitus and collagen-linked fluorescence. N Engl J Med. 1986;314(7):403–408. doi: 10.1056/NEJM198602133140702. [DOI] [PubMed] [Google Scholar]
- 49.GraphPad Software I ©2017. Grubb’s test [Internet]. [cited 2017 Oct 11]. Available from: https://www.graphpad.com/quickcalcs/Grubbs1.cfm
- 50.Peterson MD, Rhea MR, Sen A, Gordon PM. Resistance exercise for muscular strength in older adults: a meta-analysis. Ageing Res Rev [Internet] 2010;9(3):226–237. Available from: 10.1016/j.arr.2010.03.004 [DOI] [PMC free article] [PubMed]
- 51.Onambele-Pearson GL, Pearson SJ. The magnitude and character of resistance-training-induced increase in tendon stiffness at old age is gender specific. Age (Omaha) 2012;34(2):427–438. doi: 10.1007/s11357-011-9248-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Seynnes OR, Erskine RM, Maganaris CN, Longo S, Simoneau EM, Grosset JF, et al. Training-induced changes in structural and mechanical properties of the patellar tendon are related to muscle hypertrophy but not to strength gains. J Appl Physiol. 2009;107(2):523–530. doi: 10.1152/japplphysiol.00213.2009. [DOI] [PubMed] [Google Scholar]
- 53.Malliaras P, Kamal B, Nowell A, Farley T, Dhamu H, Simpson V, et al. Patellar tendon adaptation in relation to load-intensity and contraction type. J Biomech. 2013;46(11):1893–1899. doi: 10.1016/j.jbiomech.2013.04.022. [DOI] [PubMed] [Google Scholar]
- 54.Arampatzis A, Karamanidis K, Albracht K. Adaptational responses of the human Achilles tendon by modulation of the applied cyclic strain magnitude. J Exp Biol 2007;210(Pt 15):2743–2753. [DOI] [PubMed]
- 55.Lavagnino M, Arnoczky SP, Tian T, Vaupel Z. Effect of amplitude and frequency of cyclic tensile strain on the inhibition of MMP-1 mRNA expression in tendon cells: an in vitro study. Connect Tissue Res. 2003;44(3–4):181–187. doi: 10.1080/03008200390215881. [DOI] [PubMed] [Google Scholar]
- 56.Voigt M, Bojsen-Møller F, Simonsen EB, Dyhre-Poulsen P. The influence of tendon youngs modulus, dimensions and instantaneous moment arms on the efficiency of human movement. J Biomech. 1995;28(3):281–291. doi: 10.1016/0021-9290(94)00071-B. [DOI] [PubMed] [Google Scholar]
- 57.Holm L, van Hall G, Rose AJ, Miller BF, Doessing S, E a R, et al. Contraction intensity and feeding affect collagen and myofibrillar protein synthesis rates differently in human skeletal muscle. Am J Physiol Endocrinol Metab. 2010;298(23):E257–E269. doi: 10.1152/ajpendo.00609.2009. [DOI] [PubMed] [Google Scholar]
- 58.Standley RA, Harber MP, Lee JD, Konopka AR, Trappe SW, Trappe TA. Influence of aerobic cycle exercise training on patellar tendon cross-sectional area in older women. Scand J Med Sci Sport. 2013;23(3):367–373. doi: 10.1111/j.1600-0838.2011.01396.x. [DOI] [PubMed] [Google Scholar]
- 59.Kongsgaard M, Reitelseder S, Pedersen TG, Holm L, Aagaard P, Kjaer M, et al. Region specific patellar tendon hypertrophy in humans following resistance training. Acta Physiol. 2007;191(2):111–121. doi: 10.1111/j.1748-1716.2007.01714.x. [DOI] [PubMed] [Google Scholar]
- 60.Lemoine JK, Lee JD, Trappe TA. Impact of sex and chronic resistance training on human patellar tendon dry mass, collagen content, and collagen cross-linking. Am J Physiol Regul Integr Comp Physiol [Internet]. 2009;296(1):R119–24. [DOI] [PMC free article] [PubMed]
- 61.Herchenhan A, Uhlenbrock F, Eliasson P, Weis M, Eyre D, Kadler KE, et al. Lysyl oxidase activity is required for ordered collagen fibrillogenesis by tendon cells. J Biol Chem. 2015;290(26):16440–16450. doi: 10.1074/jbc.M115.641670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hansen P, Haraldsson BT, Aagaard P, Kovanen V, Avery NC, Qvortrup K, et al. Lower strength of the human posterior patellar tendon seems unrelated to mature collagen cross-linking and fibril morphology. J Appl Physiol. 2010;108(1):47–52. doi: 10.1152/japplphysiol.00944.2009. [DOI] [PubMed] [Google Scholar]
- 63.Thorpe CT, Stark RJF, Goodship AE, Birch HL. Mechanical properties of the equine superficial digital flexor tendon relate to specific collagen cross-link levels. Equine Vet J. 2010;42(SUPPL. 38):538–543. doi: 10.1111/j.2042-3306.2010.00175.x. [DOI] [PubMed] [Google Scholar]
- 64.Dubowitz N, Xue W, Long Q, Ownby JG, Olson DE, Barb D, et al. Aging is associated with increased HbA1c levels, independently of glucose levels and insulin resistance, and also with decreased HbA1c diagnostic specificity. Diabet Med [Internet]. 2014;31(8):927–935. [DOI] [PubMed]
- 65.Li Y, Fessel G, Georgiadis M, Snedeker JG. Advanced glycation end-products diminish tendon collagen fiber sliding. Matrix Biol. 2013;32(3–4):169–177. doi: 10.1016/j.matbio.2013.01.003. [DOI] [PubMed] [Google Scholar]
- 66.Eriksen C, Svensson RB, Scheijen J, Hag A. MF, Schalkwijk C, Praet SFE, et al. Systemic stiffening of mouse tail tendon is related to dietary advanced glycation end products but not high-fat diet or cholesterol. J Appl Physiol [Internet]. 2014;117(8):840–7. [DOI] [PubMed]
- 67.Svensson RB, Mulder H, Kovanen V, Magnusson SP. Fracture mechanics of collagen fibrils: influence of natural cross-links. Biophys J [Internet] 2013;104(11):2476–2484. doi: 10.1016/j.bpj.2013.04.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Uzel SGM, Buehler MJ. Molecular structure, mechanical behavior and failure mechanism of the C-terminal cross-link domain in type I collagen. J Mech Behav Biomed Mater [Internet] 2011;4(2):153–161. Available from: 10.1016/j.jmbbm.2010.07.003 [DOI] [PubMed]
- 69.Gautieri A, Passini FS, Silván U, Guizar-Sicairos M, Carimati G, Volpi P, et al. Advanced glycation end-products: Mechanics of aged collagen from molecule to tissue. Matrix Biol [Internet]. 2017;59:95–108. Available from: 10.1016/j.matbio.2016.09.001 [DOI] [PubMed]
- 70.Magnusson SP, Hansen M, Langberg H, Miller B, Haraldsson B, Westh EK, Koskinen S, Aagaard P, Kjaer M. The adaptability of tendon to loading differs in men and women. Int J Exp Pathol. 2007;88:237–240. doi: 10.1111/j.1365-2613.2007.00551.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Westh E, Kongsgaard M, Bojsen-Moller J, Aagaard P, Hansen M, Kjaer M, et al. Effect of habitual exercise on the structural and mechanical properties of human tendon, in vivo, in men and women. Scand J Med Sci Sport. 2008;18(1):23–30. doi: 10.1111/j.1600-0838.2007.00638.x. [DOI] [PubMed] [Google Scholar]
- 72.Burgess KE, Pearson SJ, Breen L, Onambélé GNL. Tendon structural and mechanical properties do not differ between genders in a healthy community-dwelling elderly population. J Orthop Res. 2009;27(6):820–825. doi: 10.1002/jor.20811. [DOI] [PubMed] [Google Scholar]
- 73.Kösters A, Wiesinger HP, Bojsen-Møller J, Müller E, Seynnes O. Influence of loading rate on patellar tendon mechanical properties in vivo. Clin Biomech. 2014;29:323–329. doi: 10.1016/j.clinbiomech.2013.12.010. [DOI] [PubMed] [Google Scholar]
- 74.Aagaard P, Simonsen EB, Andersen JL, Magnusson SP, Bojsen- Møller F, Dyhre-Poulsen P. Antagonist muscle coactivation during isokinetic knee extension. Scand J Med Sci Sport. 2000;10:58–67. doi: 10.1034/j.1600-0838.2000.010002058.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.