

Summary
Interfamily transfer of plant pattern recognition receptors (PRRs) represents a promising biotechnological approach to engineer broad‐spectrum, and potentially durable, disease resistance in crops. It is however unclear whether new recognition specificities to given pathogen‐associated molecular patterns (PAMPs) affect the interaction of the recipient plant with beneficial microbes. To test this in a direct reductionist approach, we transferred the Brassicaceae‐specific PRR ELONGATION FACTOR‐THERMO UNSTABLE RECEPTOR (EFR), conferring recognition of the bacterial EF‐Tu protein, from Arabidopsis thaliana to the legume Medicago truncatula. Constitutive EFR expression led to EFR accumulation and activation of immune responses upon treatment with the EF‐Tu‐derived elf18 peptide in leaves and roots. The interaction of M. truncatula with the bacterial symbiont Sinorhizobium meliloti is characterized by the formation of root nodules that fix atmospheric nitrogen. Although nodule numbers were slightly reduced at an early stage of the infection in EFR‐Medicago when compared to control lines, nodulation was similar in all lines at later stages. Furthermore, nodule colonization by rhizobia, and nitrogen fixation were not compromised by EFR expression. Importantly, the M. truncatula lines expressing EFR were substantially more resistant to the root bacterial pathogen Ralstonia solanacearum. Our data suggest that the transfer of EFR to M. truncatula does not impede root nodule symbiosis, but has a positive impact on disease resistance against a bacterial pathogen. In addition, our results indicate that Rhizobium can either avoid PAMP recognition during the infection process, or is able to actively suppress immune signaling.
Keywords: immunity, PAMPs, PRRs, symbiosis, disease resistance, biotechnology
Introduction
Plant pattern recognition receptors (PRRs) perceive conserved characteristic microbial features, termed pathogen‐ or microbe‐associated molecular patterns (PAMPs or MAMPs), and trigger an immune response commonly referred to as PAMP‐ or pattern‐triggered immunity (PTI). This confers basal disease resistance against adapted pathogens and plays a major role in non‐host resistance against non‐adapted pathogens (Boutrot and Zipfel, 2017; Lee et al., 2017). Typically, plant PRRs are receptor kinases (RKs) or receptor‐like proteins (RLPs), which consist of an extracellular ligand binding domain, a transmembrane domain, and an intracellular kinase (in the case of RKs) or a cytoplasmic C‐terminal extension (in the case of RLPs; Boutrot and Zipfel, 2017). While some PRRs, such as FLAGELLIN SENSING 2 (FLS2, which detects the PAMP epitope flg22 from bacterial flagellin) are present in all higher plant species, others have only evolved in certain plant families (Boller and Felix, 2009; Boutrot and Zipfel, 2017). For example, the ELONGATION FACTOR‐TU RECEPTOR (EFR), which recognizes the highly abundant and conserved bacterial protein EF‐Tu (or the PAMP epitope elf18) and has been identified in Arabidopsis thaliana, seems to be present only in Brassicaceae (Boller and Felix, 2009). Plants recognise a wide variety of PAMPs, and it is becoming increasingly clear with the identification of new plant PRRs that many PRRs have evolved in a family‐ or even species‐specific manner (Boutrot and Zipfel, 2017). Based on these observations, the ability to transfer novel PAMP recognition capabilities across plant species, families or even classes, represents a promising biotechnological strategy to engineer broad‐spectrum (and potentially durable) disease resistance in crops (Boutrot and Zipfel, 2017; Dangl et al., 2013; Michelmore et al., 2017; Rodriguez‐Moreno et al., 2017). For example, the transgenic expression of EFR in other plant species, such as tomato (Solanum lycopersicum), Nicotiana benthamiana, wheat (Triticum aestivum), or rice (Oryza sativa) confers elf18 recognition and quantitative resistance to a range of bacterial pathogens including Ralstonia solanacearum, Pseudomonas syringae, Xanthomonas perforans, X. oryzae and Acidovorax avenae (Lacombe et al., 2010; Lu et al., 2015; Schoonbeek et al., 2015; Schwessinger et al., 2015; Zipfel et al., 2006). In addition, the PRR XA21 (which recognises the tyrosine‐sulfated peptide RaxX; Pruitt et al., 2015) from the wild rice O. longistaminata confers increased resistance against Xanthomonas spp. when expressed in banana (Musa sp.), sweet orange (Citrus sinensis) or N. benthamiana (Holton et al., 2015; Mendes et al., 2010; Tripathi et al., 2014). Similarly, the PRR ELICITIN RECEPTOR (ELR) from the wild potato S. microdontum or the A. thaliana PRR RECEPTOR‐LIKE PROTEIN 23 (RLP23, which recognises the taxonomically conserved peptide nlp20) confer increased resistance to the oomycete Phytophthora infestans when expressed in cultivated potato (S. tuberosum) (Albert et al., 2015; Du et al., 2015). These recent selected examples illustrate that PRRs normally restricted to specific plant taxonomic lineages can remain functional when expressed in other plant species. Beyond the biotechnological usefulness of this property, this also illustrates that immune signaling components acting downstream of PRRs must be (at least partially) functionally conserved.
While plants must constantly defend themselves against potential invaders, they also form close interactions with beneficial microbes, in what is commonly referred to as the plant microbiome (Hacquard et al., 2017; Müller et al., 2016). While all plants express PRRs as part of their innate immune system, it is however still unclear whether the engineering of novel PAMP recognition specificities through heterologous PRR expression affects the beneficial interaction of plants with commensal microbes.
The symbiosis between rhizobia and legumes is a defined and well understood interaction involving mutual communication. The symbiotic interaction starts with plant roots secreting chemical signals, including flavonoids, to attract host‐compatible rhizobia. In turn, rhizobia produce symbiosis‐inducing Nod factor, which is perceived by the plant and triggers two independent, yet coordinated, developmental processes: nodule organogenesis and bacterial infection (Madsen et al., 2010; Murray et al., 2007; Oldroyd et al., 2011; Radutoiu et al., 2003; Tirichine et al., 2006). Bacteria attach to the root hair tip and form micro‐colonies, from which they invade the plant tissue by growing inside a tubular structure called an infection thread. In parallel to the root hair infection, plant cortical cells divide and develop a new organ, known as a nodule, which ultimately accommodates the rhizobia. In mature nodules, bacteria live as membrane‐encased bacteroids and fix atmospheric nitrogen making it available to the plant (Oldroyd et al., 2011; Udvardi and Poole, 2013). The harmonious interplay between both organisms requires continuous signal exchange and can be terminated at various stages (Cao et al., 2017; Ferguson et al., 2018; Gibson et al., 2008).
Although the role of PAMP perception and immune signaling during symbiosis has not been extensively studied, there is accumulating evidence to suggest that rhizobia are initially perceived as potential invaders (Cao et al., 2017; Gourion et al., 2015). The apparent overlap of components and concepts between immunity and symbiosis signaling pathways in legumes is both intriguing, and relevant to the question about the importance of PAMP recognition during these contrasting processes (Zipfel and Oldroyd, 2017). Rhizobia are clearly capable of eliciting PTI, because suspension cultures of Mesorhizobium loti can trigger defense‐associated responses in the legume Lotus japonicus, such as ethylene production, MAP kinase (MAPK) activation and immune gene transcription in a similar way to the flg22 peptide (Lopez‐Gomez et al., 2012). In addition, PTI signaling triggered by exogenous application of the Pseudomonas aeruginosa‐derived flg22 peptide delays nodulation and reduces nodule numbers during the symbiosis between L. japonicus and M. loti (Lopez‐Gomez et al., 2012). Transcriptional studies in different legume species also reported the transient upregulation of immune‐related genes upon first encounter with its rhizobial symbiont, followed by a downregulation during the onset of symbiosis. For example, immune and stress‐related genes in M. truncatula roots were upregulated 1 h post‐inoculation (hpi) and subsequently downregulated to a minimal level at 12 hpi in response to S. meliloti inoculation (Lohar et al., 2006). Similarly, transcriptome analysis of root hair cells from soybean (Libault et al., 2010) and M. truncatula (Breakspear et al., 2014) showed induction of defense genes 24 hpi, then a marked reduction at later time‐points after infection with Bradyrhizobium japonicum and S. meliloti, respectively. Interestingly, the early activation of plant defense may play a role in the selection of symbionts and endophytes versus pathogens during the early stages of the rhizobia‐legume interaction (Zgadzaj et al., 2015).
To test directly in a reductionist approach whether a novel PAMP recognition ability affects the symbiotic interaction between legumes and rhizobia, we expressed the A. thaliana EFR (EFR) gene in M. truncatula to engineer the perception of EF‐Tu (or elf18 peptide) from its symbiont S. meliloti. After confirming EFR functionality, we then tested if EFR expression had an impact on rhizobial infection and the symbiotic interaction. While infection was unaffected, nodulation was slightly reduced at an early time‐point, but recovered fully by the later stages of symbiosis. Importantly, rhizobia in nodulated EFR‐Medicago plants fixed atmospheric nitrogen as efficiently as in control plants. Despite the lack of effect on rhizobial infection and nodulation, EFR expression conferred quantitative resistance to the bacterial root pathogen R. solanacearum, suggesting that the transfer of PRRs is an efficient biotechnological tool to confer increased legume resistance to pathogens, with minimal impact on symbiotic interactions.
Results
Transgenic expression of EFR in Medicago truncatula confers elf18 recognition in leaves and roots
To study the effect of EFR expression on symbiotic and pathogenic interactions with M. truncatula, we stably transformed M. truncatula ecotype R108 with pCaMV35S::EFR‐HA by Agrobacterium tumefaciens‐mediated transformation (Cosson et al., 2006). Two independently transformed lines with a single insertion event were isolated (lines 26‐8 and 18‐1) and characterized alongside their respective null segregants as controls (lines 26‐2 and 18‐3). Transgenic EFR‐Medicago plants showed similar growth and development as their control lines (Figure S1), and EFR accumulation could be detected in leaf and root tissue of both EFR‐Medicago lines by western blot analysis (Figure 1a,b). EFR specifically perceives the PAMP elf18 from various bacterial species (Figure S2), including the M. truncatula symbiont S. meliloti, and initiates immune signaling (Kunze et al., 2004; Lacombe et al., 2010). Transgenic EFR‐Medicago plants responded to local treatment with the elf18 peptide by production of a transient burst of reactive oxygen species (ROS) in leaves (Figure 1c) and roots (Figure 1d). We also tested responsiveness to the PAMP flg22, and confirmed that all lines responded to the peptide (Figure S3), showing that the presence of EFR does not interfere with the function of an endogenous PRR (i.e. FLS2). In addition, phosphorylation of MAPKs and induction of immune‐related genes were observed in the EFR‐expressing lines 18‐1 and 26‐8 only after elf18 treatment (Figure S4), showing that these lines do not exhibit constitutive activation of immune signaling. These results show that EFR is functional in M. truncatula as it provides responsiveness to the PAMP elf18.
Figure 1.
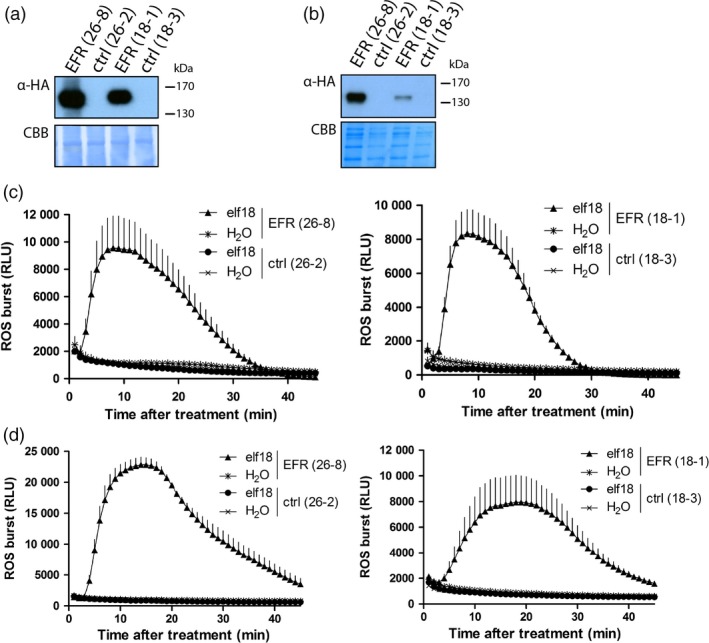
Transgenic EFR‐Medicago responds to elf18 peptide. Western blot of leaf (a) and root (b) material from indicated lines using α‐HA antibody to detect AtEFR‐HA. Blot was stained with Coomassie Brilliant Blue (CBB) as loading control. This experiment was repeated four times with similar results. ROS burst was monitored in (c) leaf discs and (d) root segments from lines 26‐8 (left panels) and 18‐1 (right panels) after 100 nM elf18 treatment and displayed as relative light units (RLU). Values are means ± standard error (n = 8). The experiment was done three times.
EFR expression does not affect the long‐term symbiosis between S. meliloti and M. truncatula
We next tested whether heterologous expression of EFR affects the symbiosis between S. meliloti and M. truncatula. Expression of EFR in M. truncatula did not have a negative effect on plant growth after inoculation with S. meliloti, as the plant phenotype and fresh weight were similar in EFR‐expressing and control plants when symbiosis was established at 4 weeks after inoculation (Figure 2a,b). EFR expression is driven by the ubiquitous CaMV35S promoter, and we were able to detect EFR accumulation in different root tissues, such as the main root, lateral roots and nodules (Figure S5).
Figure 2.
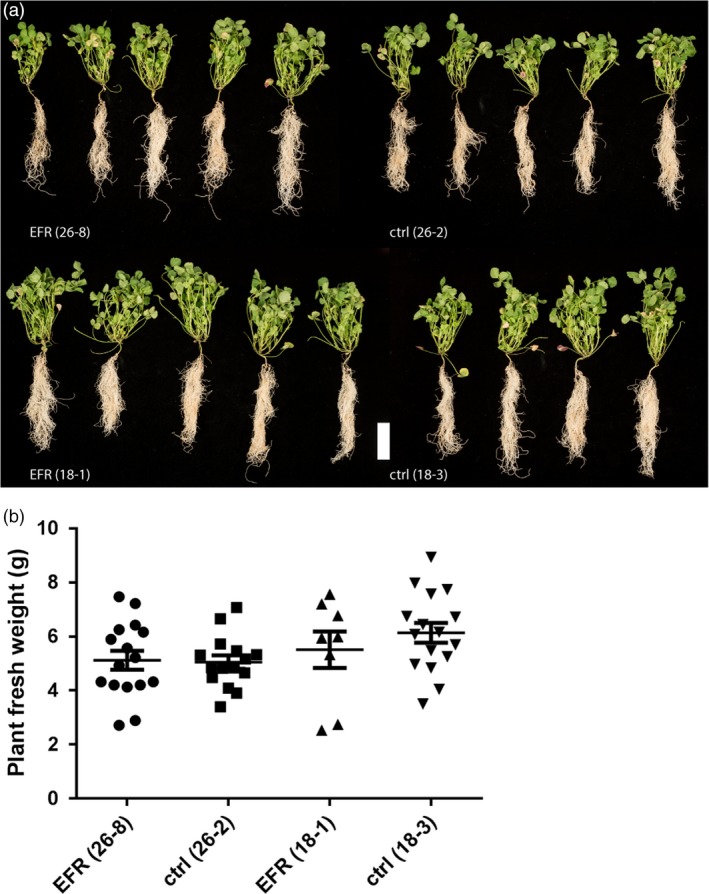
EFR expression does not affect development and fresh weight of M. truncatula infected with S. meliloti. (a) Plant pictures and (b) fresh weight was assessed of 5‐week‐old M. truncatula plants expressing EFR (26‐8 and 18‐1) and respective control lines (26‐2 and 18‐3) inoculated with Sm1021‐lacZ and harvested at 28 dpi. White scale bar, 5 cm. The experiment was done three times.
Next, we looked at different stages of the rhizobial infection and the nodulation process. Perception of PAMPs and PTI signaling presumably happens at the beginning of an infection, when the plant first encounters the microbe. We therefore tested whether EF‐Tu recognition affects symbiotic interaction at this early stage. The formation of micro‐colonies at the root hair tip, the number of infection threads and nodule primordia were similar between EFR‐Medicago and the control lines (Figure 3a). Scoring total nodule numbers of the root at an early time‐point (10 dpi) we observed a small, but significant, reduction in EFR‐Medicago lines compared to control lines (Figure 3b). Total nodule numbers were reduced by 35% in line 26‐8 and by 25% in line 18‐1. Importantly, all nodules were colonized by rhizobia, as detected by staining for β‐galactosidase activity in nodules colonized by the S. meliloti strain 1021‐lacZ, and the spectrum of nodule morphology was similar for all lines. Notably, we observed a significant difference between the transformed null segregant control line 18‐3 and the untransformed wild‐type (Figure 3b), which encouraged us to use the null segregants as an appropriate control to avoid artefacts that could be linked to the genetic transformation and/or the associated in vitro culture process.
Figure 3.
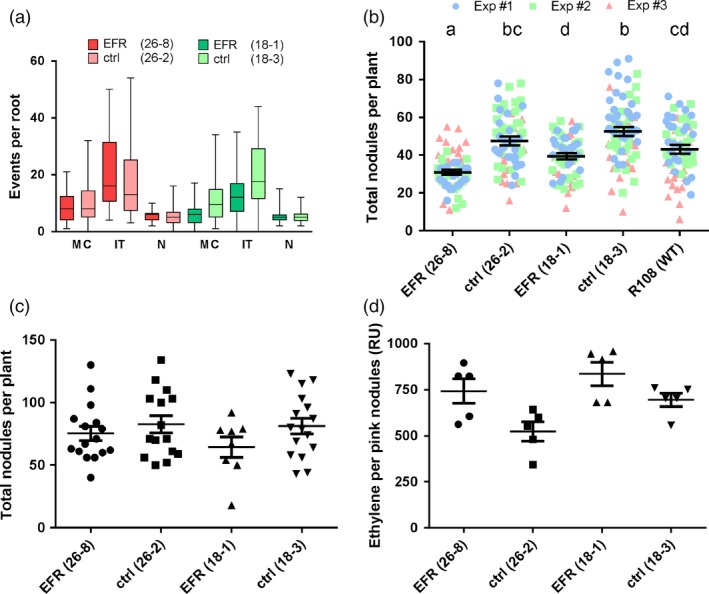
Symbiosis between M. truncatula and S. meliloti is not affected by EFR expression. (a) Infection events were scored at 7 dpi on roots of M. truncatula lines expressing EFR (26‐8 and 18‐1) and control lines (26‐2 and 18‐3) infected with Sm1021‐lacZ. MC: micro‐colonies. IT: infection threads. N: nodule primordia. Data from three independent experiments (each n = 10) were combined. (b) Total nodules were scored at 10 dpi on roots of M. truncatula lines expressing EFR (26‐8 and 18‐1), control lines (26‐2 and 18‐3) and untransformed wild‐type R108 infected with Sm1021‐lacZ. Data from three independent experiments (each n = 25) were combined. Letters indicate statistical significance groups with P < 0.05 after One‐way ANOVA (Kruskal–Wallis's test and Dunn's multiple comparison). (c) Total nodules were scored at 28 dpi on roots of M. truncatula lines expressing EFR (26‐8 and 18‐1) and control lines (26‐2 and 18‐3) infected with Sm1021‐lacZ. One‐way ANOVA with P < 0.05 did not indicate statistical significant differences. (d) Acetylene reduction to ethylene was measured on whole plants of M. truncatula lines expressing EFR (26‐8 and 18‐1) and control lines (26‐2 and 18‐3) infected with Sm1021 —lacZ at 28 dpi. Production of ethylene is displayed as relative units (RU) per pink nodules of each root system. One‐way ANOVA with P < 0.05 did not indicate statistical significant differences. The experiments were performed three times.
We next assessed nodulation during the later stages of symbiosis. Four weeks after inoculation, symbiosis is well established and nodules are actively fixing atmospheric nitrogen (Oldroyd et al., 2011). By this stage, there was no difference in total nodule numbers between either of the EFR‐Medicago lines and their respective controls (Figure 3c). In addition, we measured the enzymatic activity of rhizobial nitrogenase inside nodules, which can be used as an indicator of the nitrogen fixation rate (Price et al., 2015). Notably, root systems from EFR‐Medicago and from the control lines fixed nitrogen at similar rates (Figure 3d). In addition, the nodule morphology was similar in all lines, and no macroscopic signs of defense phenotypes or early senescence could be observed.
Together, our results thus indicate that EFR expression in M. truncatula may cause a slight initial delay in nodule formation, but overall does not negatively affect either rhizobial infection or long term‐nitrogen‐fixing symbiosis.
EFR expression increases the resistance of M. truncatula to the bacterial root pathogen R. solanacearum
The proteobacterium R. solanacearum is a root pathogen that causes bacterial wilt disease in different plant species including M. truncatula (Mansfield et al., 2012; Vailleau et al., 2007). M. truncatula plants infected with R. solanacearum develop disease symptoms such as chlorosis and wilting, ultimately leading to plant death (Vailleau et al., 2007). To test whether EFR can protect M. truncatula against bacterial pathogens, we infected EFR‐Medicago and control lines with R. solanacearum GMI1000, and monitored disease progression and survival of the plants over several days. EFR‐expressing plants displayed a consistently higher survival rate than plants from the control lines (Figure 4a,b). Although these results were only statistically significant for line 26‐8 (based on Mantel‐Cox test), we observed a tendency for slower disease progression in line 18‐1 across five independent experiments (Figure 4b and Figure S6). A possible explanation for the enhanced disease resistance of line 26‐8 compared to the transgenic line 18‐3 may be the different EFR accumulation levels in these plants (Figure 1a,b and Figure S3), which also translate into stronger elf18‐induced ROS production in the root of line 26‐8 compared to the line 18‐1 (Figure S7).
Figure 4.
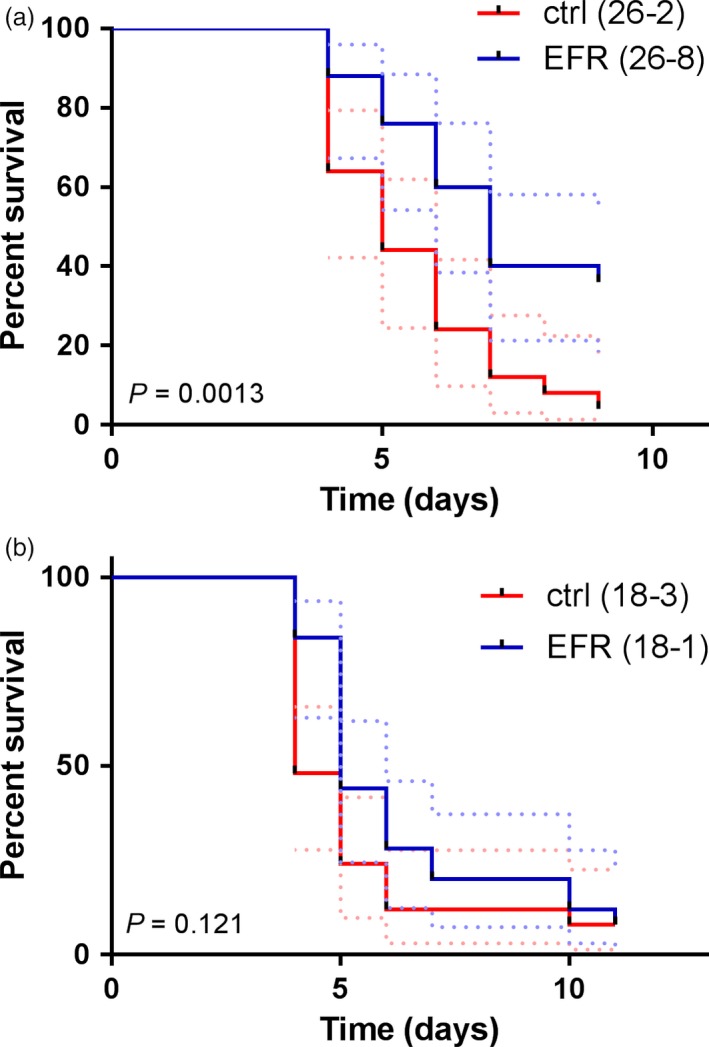
EFR expression in M. truncatula provides quantitative resistance against the pathogen R. solanacearum. (a) M. truncatula lines expressing EFR 26‐8 and control line 26‐2 were infected with R. solanacearum GMI1000 and d ase symptoms assessed daily. Survival rate is displayed over 9 days and statistical analysis performed with Mantel‐Cox test, P = 0.0013 (n = 25). Experiment was repeated four times with similar results. (b) M. truncatula lines expressing EFR 18‐1 and control line 18‐3 were infected with R. solanacearum GMI1000 and disease symptoms assessed daily. Survival rate is displayed over time and statistical analysis performed with Mantel‐Cox test, P = 0.121 (n = 25). Dashed lines represent 95% confidence interval. Experiment was performed four times for 26‐2 and 26‐8, and five times for 18‐3 and 18‐1, with similar tendency (Figure S6).
Overall, as previously observed with other EFR‐expressing plant species infected with bacterial pathogens (Lacombe et al., 2010; Lu et al., 2015; Schoonbeek et al., 2015; Schwessinger et al., 2015; Zipfel et al., 2006), EFR conferred increased quantitative resistance, delayed disease progression and increased survival under our experimental conditions.
Discussion
Many PRRs have been successfully expressed in heterologous hosts across taxonomically diverse plant species to improve disease resistance, and PRRs have thus become attractive tools as part of the biotechnological arsenal to genetically engineer disease resistance in crops (Boutrot and Zipfel, 2017; Dangl et al., 2013; Michelmore et al., 2017; Rodriguez‐Moreno et al., 2017). It was however still unclear whether such PRR transfer may negatively impact the association of the recipient plants with symbiotic microbes.
In this study, we generated transgenic M. truncatula lines that express the heterologous PRR EFR to confer recognition of the endogenous PAMP EF‐Tu from the symbiotic bacterium S. meliloti. These EFR‐Medicago plants recognized elf18 peptide and initiated a PAMP‐induced ROS burst in both leaves and roots (Figure 1c,d). As previously reported (Holton et al., 2015; Schwessinger et al., 2015), these results indicate components involved in the PTI signaling pathway (and in PRR biogenesis) are also present and conserved in the root and leaves of M. truncatula, as they enable the functionality of EFR.
Legumes benefit from the symbiosis with rhizobia during growth in nitrogen‐limited soil due to the additional bioavailable nitrogen supplied by the bacteria. The growth and development of plants with an established symbiosis was similar between transgenic EFR‐Medicago and control lines. Therefore, we concluded that EFR expression did not exert a negative effect on growth of infected plants, or on long‐term symbiosis (Figure 2a,b). Likewise, early interaction events were unaltered by EFR expression. There was no major difference in the occurrence of micro‐colonies, infection threads and nodule primordia formation between EFR‐Medicago and control lines (Figure 3a). Although infection and nodulation are triggered simultaneously, both processes belong to different developmental programs (Murray et al., 2007; Tirichine et al., 2006). Interestingly, at an early time‐point after rhizobial inoculation nodule numbers were slightly reduced in EFR expressing lines (Figure 3b). However, at later stages of symbiosis nodule numbers were similar in all lines (Figure 3c). These data indicate that while nodulation might be delayed at early stages, the expression of EFR did not impede long‐term nodulation. Importantly, the nitrogen‐fixation capacity of nodules was unaffected by EFR expression (Figure 3d).
The absence of detrimental effects of EFR expression on rhizobial nitrogen‐fixing symbiosis may at first appear counterintuitive. Indeed, the EF‐Tu‐derived EFR ligand elf18 from S. meliloti is able to induce immune responses (Kunze et al., 2004; Lacombe et al., 2010), and it has previously been shown that elicitation of PTI using exogenous PAMP treatment can affect the interaction between rhizobia and legumes. For example, application of flg22 peptide or M. loti cells to L. japonicus triggered similar PTI responses leading to a delay in nodule formation and reduced nodule numbers (Lopez‐Gomez et al., 2012). Furthermore, co‐inoculation experiments in M. truncatula recently showed that the pathogenic bacterium P. syringae pv. tomato (Pto) DC3000 induces immune responses and suppresses the establishment of the symbiosis with S. meliloti (Chen et al., 2017). In addition, constitutive activation of immune responses in M. truncatula in specific mutants or over‐expression lines impairs nodule formation and symbiosis (Berrabah et al., 2014, 2015; Bourcy et al., 2013; Domonkos et al., 2013; Ryu et al., 2017; Wang et al., 2016). Interestingly, nodulation in M. truncatula was only impaired when plants were co‐treated with the PAMP peptide flg22 together with M. loti, but not when flg22 was applied after symbiosis was established. PAMP treatment seems to delay nodule development rather than impairing rhizobial fitness, as spontaneously nodulating snf1 mutant plants were similarly affected by flg22 treatment (Lopez‐Gomez et al., 2012). It thus appears that the timing of PTI activation and symbiotic signaling may be important to study the impact of PAMP recognition on symbiosis. In this context, it is important to note that EFR expression in recipient transgenic plants does not seem associated with constitutive activation of immune responses, as no detrimental effects on plant growth or development have been observed in these plants, in either axenic or non‐sterile soil conditions (Lacombe et al., 2010; Lu et al., 2015; Schoonbeek et al., 2015; Schwessinger et al., 2015; Figure 2 and Figure S1). Thus, our findings that EFR expression negatively affects early nodulation but not infection events or nodule numbers, as well as the observation that nitrogen fixation was unchanged at later stages of symbiosis, support the notion that the perception of rhizobial PAMPs might have an early transient effect on plant nodulation but does not compromise rhizobial fitness, infection, or the ultimate establishment of a functional nitrogen‐fixing symbiosis during the natural infection process.
Our data therefore suggest that during a natural infection process, S. meliloti either evades EF‐Tu recognition or actively suppresses PTI in the host. While rhizobia are known to carry a flg22 allele of the flagellin gene that is not recognized by the plant FLS2 receptor (Felix et al., 1999), the EF‐Tu‐derived elf18 peptide from S. meliloti is recognised by EFR (Kunze et al., 2004; Lacombe et al., 2010), suggesting that an immune evasion strategy is not conceivable here. Despite the manifold evidence that EFR expression provides efficient disease resistance, it is still unclear how the intracellular EF‐Tu protein (and by extension the elf18 epitope) gets exposed to the EFR receptor during infection (Zipfel et al., 2006). EF‐Tu has been found in the cell‐free supernatant from cultures of different bacterial species (Kazemi‐Pour et al., 2004; Watt et al., 2005), and an active role for EF‐Tu has been suggested during effector translocation via the type‐6 secretion system (T6SS) in P. aeruginosa (Whitney et al., 2015). Interestingly, EF‐Tu was recently identified in bacterial outer membrane vesicles, which was linked to the ability of these vesicles to induce EFR‐dependent immune responses in A. thaliana (Bahar et al., 2016), illustrating a possible mechanism by which this potent PAMP can be released. While we cannot therefore completely exclude the possibility that rhizobia can control the release of EF‐Tu proteins, this seems unlikely as legumes do not normally express a functional EFR homolog, and thus no selective advantage would be conferred through this strategy.
Previous transcriptomic studies indicate that rhizobia initially elicit an immune response, which is then suppressed as symbiosis proceeds (Breakspear et al., 2014; Libault et al., 2010; Lohar et al., 2006). In addition, co‐inoculation with S. meliloti suppresses immune responses normally triggered by Pto DC3000 in M. truncatula (Chen et al., 2017). These results suggest that rhizobia have active mechanisms to suppress PTI. Consistent with this, it has recently been shown that the plant‐growth promoting rhizobacterium Pseudomonas simiae WCS417 actively suppresses many transcriptional changes induced by the PAMP flg22 (Stringlis et al., 2018). Plant pathogenic bacteria can suppress host immunity by secretion of effectors, many of which interfere with the canonical PTI pathway at different stages (Pfeilmeier et al., 2016). Many of these PTI‐suppressing effectors are translocated within plant cells via the type‐3 secretion system (T3SS) (Macho and Zipfel, 2015). While there is evidence suggesting that rhizobia also use effectors to suppress plant immunity, only a few rhizobial effectors (Nop proteins) have been characterized (Staehelin and Krishnan, 2015). Rhizobial genomes encode several different secretion pathways, and the importance of T3SS, type‐4 secretion systems (T4SS) and T6SS for symbiosis has been demonstrated genetically in certain rhizobial species (Bladergroen et al., 2003; Nelson and Sadowsky, 2015; Sugawara et al., 2013). Sinorhizobium sp. NGR234 translocates multiple type‐3 secreted effectors including NopM, NopL, NopP and NopT to interfere with immune signaling (Dai et al., 2008; Ge et al., 2016; Skorpil et al., 2005; Xin et al., 2012). However, the S. meliloti strain 1021 used in our study, Sm1021, only carries a T4SS gene cluster for translocation of effectors into host cytoplasm, and deletion mutant studies showed no impact of mutating the T4SS on nodulation (Jones et al., 2007; Nelson et al., 2016).
Other bacterial mechanisms have been suggested to suppress PTI (Cao et al., 2017). For example, Nod factors not only play a role in initiating and maintaining symbiosis signaling, but also in suppression of plant immunity (Liang et al., 2013). Application of B. japonicum Nod factor to the non‐host plant A. thaliana resulted in reduced accumulation of the immune receptors FLS2 and EFR at the plasma membrane (Liang et al., 2013). Although this partial suppression of PTI seems to be conserved in legume and non‐legume plants, the impact on rhizobial‐legume root infection has not been directly tested. Contrary to these findings, we could detect EFR accumulation in the nodules of our transgenic M. truncatula plants (Figure S5). Exopolysaccharide (EPS) production is a common factor among plant‐associated bacteria and has been previously associated with evasion of PTI (Aslam et al., 2008; D'Haeze and Holsters, 2004). Cell surface polysaccharides such as EPS, lipopolysaccharides (LPS) and cyclic β‐glucans have been implicated in facilitating symbiotic interaction (Mithofer et al., 1996; Niehaus et al., 1993; Tellstroem et al., 2007). Notably, the EPS receptor EPR3 from L. japonicus specifically detects EPS from its symbiont and acts as a positive regulator of infection (Kawaharada et al., 2015; Muszynski et al., 2016). Mutant strains defective in cell surface polysaccharides result in impaired infections or ineffective nitrogen‐fixing nodules (D'Haeze and Holsters, 2004). For example, the succinoglycan‐deficient Sm1021 exoY mutant induces immune‐related genes more strongly than wild‐type, indicating a possible involvement of succinoglycan in the suppression of immunity (Jones et al., 2008). Additionally, purified LPS from S. meliloti can suppress ROS burst in M. truncatula suspension cells treated with invertase (Tellstroem et al., 2007). However, the phenotypes of cell surface polysaccharide mutants are often difficult to interpret, because they seem to be specific for the type of polysaccharides, the rhizobial species and the host plant. It is therefore likely that cell surface polysaccharides contribute to the symbiotic interaction in multiple ways in addition to facilitating immune evasion (Gourion et al., 2015). It will be thus interesting to investigate in future studies the exact mechanisms employed by rhizobia to suppress PTI. It is also conceivable that legumes themselves specifically suppress PTI in a local and timely manner in response to rhizobial signals to facilitate infection by their symbionts. For example, limiting trafficking of PRRs into the infection thread and/or peribacteroid membranes could restrict recognition of those PAMPs. In addition, it has been recently shown that the Medicago genes DNF2 and SymCRK contribute to the repression of plant defences via the inhibition of the ethylene pathway in nodules (Berrabah et al., 2018).
The transfer of EFR has already been shown to confer increased quantitative resistance against different bacterial pathogens in a wide range of plant species, including tomato, N. benthamiana, wheat and rice (Lacombe et al., 2010; Lu et al., 2015; Schoonbeek et al., 2015; Schwessinger et al., 2015; Zipfel et al., 2006). The present study expands this list and reveals that EFR is also functional when expressed in legumes (at least as demonstrated here for M. truncatula) and increases the survival of M. truncatula plants upon inoculation by the root bacterial pathogen R. solanacearum (Figure 4a,b and Figure S6). Thus, together, our data demonstrate that EFR expression can protect M. truncatula from the destructive root pathogen R. solanacearum, without compromising the overall symbiotic interaction with S. meliloti, which allows fixation of atmospheric nitrogen. Our results suggest that legumes can be engineered with novel PRRs without affecting the nitrogen‐fixing symbiosis, and may also be relevant in the future as attempts to transfer this important symbiosis into non‐legume plants are currently ongoing (Zipfel and Oldroyd, 2017). More generally, it also illustrates that the transfer of PRRs across plant species does not necessarily come at a cost for the plant, but actually increases its fitness when faced with aggressive pathogens. It will be interesting in the future to expand the reductionist approach used in this study to test whether heterologous PRR expression affects the composition and function of other commensals in the plant microbiome. A potential effect on the microbiome would then however need to be reconciled with the absence of obvious growth defects of transgenic plants expressing PRRs in non‐sterile soil, and counterbalanced in an agricultural sense against the benefit conferred by PRR transfer in term of disease resistance under strong pathogen pressure.
Material and methods
Bacterial growth
S. meliloti strain 1021 (also known as Rm1021) pXLGD4 phemA::lacZ (Sm1021‐lacZ) (Leong et al., 1985) was grown at 30 °C in TY medium (tryptone 5 g/L, yeast extract 3 g/L) containing appropriate antibiotics, streptomycin 50 μg/mL and tetracycline 12.5 μg/mL. Ralstonia solanacearum strain GMI1000 was grown at 28 °C in complete BG medium (bacto peptone 10 g/L, casamino acid 1 g/L, yeast extract 1 g/L).
Plant growth
M. truncatula seeds were scarified with 98% sulfuric acid treatment for 8 min, extensively washed with water and then surface‐sterilized with 10% NaOCl for 2 min. After washing with sterile water, the seeds were left for 3 h to imbibe water before being placed on inverted agar plates in the dark for 3 days at 4 °C and subsequently germinated over‐night at 20 °C. For sterile growth, seedlings were transferred to squared 1% agar plates and sandwiched between two Whatman filter papers (GE Healthcare, Amersham, UK). Plates were incubated vertically in a growth chamber at 21 °C, with a 16 h light period and 80% rel. humidity.
For growth in soil, germinated seedlings were transferred to sterile 1 : 1 mixture of terragreen (Oil‐dri UK ltd, Wisbech, UK) and sharp sand (BB Minerals, Norwich, UK) for rhizobial inoculations, in loam based compost (John Innes Cereal Mix) for seed bulking, or in Jiffy Peat Pellets for inoculation with Ralstonia solanacearum. Plants were grown in controlled environment chambers with a 16‐h photoperiod at 20 °C with 120–180 μmol/m2/s2 light intensity and 80% rel. humidity.
Stable transformation of Medicago truncatula
M. truncatula ecotype R108 was stably transformed by A. tumefaciens AGL1 carrying the recombinant binary vector pBIN19‐CaMV35S::EFR‐HA (Lacombe et al., 2010). Plant transformation was carried out as previously described (Cosson et al., 2006) with some minor, but important, changes; in vitro grown plants, only, were used for the transformations, excised leaves were sliced through with a scalpel and not vacuum infiltrated, the A. tumefaciens culture was used at OD600 = 0.4 and re‐suspended in SH3a broth with 300 μm acetosyringone, the leaflet explants were submerged in the bacterial suspension for 20 min only, shaking in the dark, leaves were co‐cultivated and callus generated with their adaxial surface in contact with the media, explants were washed in SH3a media broth post co‐cultivation, excess Agrobacteria was eliminated on solid media using 320 mg/L ticarcillin disodium/potassium clavulanate and finally, callus growth was carried out in the dark for 8 weeks rather than 6. Five transgenic plants were recovered by somatic embryogenesis, rescued and selected on kanamycin plates. Homozygous plants were identified by quantitative real‐time PCR of a segregating T1 population and confirmed in the T2 stage by responsiveness to elf18 peptide. Two homozygous lines with only a single insertion locus carrying two EFR copies were identified and used for physiological and symbiotic characterizations. In addition, null segregants were isolated for each primary transformant and used as control lines. All experiments were performed with plants from the T3 population.
Rhizobial inoculations
Plants were grown in pots (50 or 80 mL volume) in terragreen/sharp sand mix for 2 days (infection thread counting) or 7 days (nodule counting and acetylene reduction measurements) before inoculation with Sm1021‐lacZ. Bacteria were grown in TY to OD600 = 1.5, washed in 10 mm MgCl2 and diluted to OD600 = 0.0002. Plants were inoculated with 5 mL of S. meliloti suspension equally spread across the pot. Plants were harvested at indicated time‐points, carefully rinsed with water and stained with X‐Gal for visualization of LacZ activity. Stained nodules were counted under a stereo microscope.
For late‐time point experiments (e.g. 28 dpi), plants were grown in bigger pots (500 mL volume) to allow enough space for root development and were inoculated with 10 mL S. meliloti diluted to OD600 = 0.0002. Plant nodules were scored visually and were not stained.
X‐Gal staining of infection structures and nodules
For staining of infection threads on plants grown in terragreen/sand mixture, whole roots were detached from shoot and placed in fixing solution containing phosphate buffer, pH = 7 (61 mm Na2HPO4, 39 mm NaH2PO4, 10 mm KCl, 1 mm MgCl2) and 2.5% glutaraldehyde. Vacuum was applied for 5 min and roots were incubated for 1 h at room temperature. Roots were washed three times in phosphate buffer before staining solution (5 mm K4[Fe(CN)6], 5 mm K3[Fe(CN)6], 0.08% X‐Gal (Formedium, King's Lynn, UK) in phosphate buffer) was added and roots incubated in the dark at 30 °C over‐night. Roots were washed in phosphate buffer three times and stored at 4 °C until analysis.
Stained infection structures were scored in brightfield mode using a Leica DMR microscope (Leica, Wetzlar, Germany). The infection events were classified into three categories: micro‐colony formation at curled root hair, elongated and ramified infection threads and nodule primordia.
Ralstonia solanacearum infection of Medicago truncatula
After germination, M. truncatula plants were transferred and grown in jiffy peat pellets. For inoculation, the 1/3 bottom half of the Jiffy pots was severed then soaked in a R. solanacearum solution at OD600 = 0.1. Potting soil was used to absorb the remaining inoculum and spread on the bottom of the tray before putting Jiffy pots back on. Disease symptoms were scored daily. Statistical analysis was performed as previously outlined (Remigi et al., 2011).
Acetylene reduction measurements
Nitrogenase activity was determined by gas chromatography measuring the enzymatic conversion of acetylene gas to ethylene as previously described (Trinick et al., 1976). Infected plants at 28 dpi, placed in a 50‐mL plastic vial and sealed with a rubber lid. Acetylene gas (BOC, Manchester, UK) was injected into the vials with 2% (v/v) final concentration, incubated for 1 h at 23 °C and 1 mL sample taken for analysis. Conversion of acetylene to ethylene by rhizobial nitrogenase was recorded on a Clarus 480 (Perkin Elmer) gas chromatograph with N2 as the carrier gas set to a flow rate of 25 mL/min, a HayeSep N 80/100 mesh column, connected to a flame ionisation detector at 100 °C. Acetylene was applied in excess and peak areas of ethylene were quantified using TotalChrom Workstation software (Perkin Elmer) and displayed as relative units.
ROS burst measurements
M. truncatula seedlings were grown sterile for 7 days under 16‐h photoperiod at 21 °C. Roots were cut into 3 mm segments and recovered in water over‐night. Alternatively, leaf discs were sampled from soil‐grown 4–5 weeks‐old plants and recovered in water. ROS burst was measured as described previously. The water was replaced with solution containing 200 μg/mL horseradish peroxidase (HRP) (Sigma‐Aldrich, Gillingham, UK) and 1 μm L‐012 (Sigma‐Aldrich), incubated for 5 min and topped up with solution containing flg22 or elf18 peptide (EZBiolab, Westfield, IN) with 100 nm final concentration. Flg22 and elf18 peptide sequences are derived from P. aeruginosa and E. coli, respectively (Figure S2). Luminescence was recorded over 45 min using a charge‐coupled device camera (Photek Ltd., St Leonards on Sea, East Sussex, UK).
MAPK activation and gene induction
For MAPK activation and gene induction, tissues from 13‐day‐old seedlings, grown in vitro on modified Fahraeus medium, were transferred to 24‐well plates containing 0.5 mL water. Three leaf discs (4 mm) or 10 root segments (1 cm) were used for each condition. The next day, tissues were treated with 0.5 mL of either 1 μm elf18 solution or water. After incubation, tissues were dried on paper and then frozen in liquid nitrogen immediately for protein or RNA extraction.
RNA extraction and qPCR analysis
Total RNA was extracted using Trizol reagent (Invitrogen, Loughborough, UK) according to the manufacturer's instructions. RNA samples were treated with Turbo DNA‐free DNase (Ambion/Thermo fisher Scientific, Waltham, MA) according to the manufacturer's instructions. RNA was quantified with a Nanodrop spectrophotometer (Thermo fisher Scientific). cDNA was synthesized from RNA using RevertAid (Thermo fisher Scientific) according to the manufacturer's instructions. cDNA was amplified by quantitative PCR using SYBR Green JumpStart Taq ReadyMix (Sigma‐Aldrich) and the PTC‐200 Peltier Thermal Cycler (Bio‐Rad Laboratories, Hercules, CA). The relative expression values were determined using Actin as reference and the comparative Ct method (). The following primers were used for quantitative RT‐PCR: MtACTIN2 (Medtr2g008050) F: TGGCATCACTCAGTACCTTTCAACAG; R: ACCCAAAGCATCAAATAATAAGTCAACC; MtWRKY33 (Medtr3g031220.1) F: ACCAGGAATGGCAATGGAAGGTC; R: AATAGCCTCTTGGATGCATGGC; MtMAPK3 (Medtr4g061130)F: TGGTTGCTGTGAAGAAGATA; R: TGAGTTCGGTGGTTATGTAA (Chen et al., 2017).
Immunoblot analysis
Plant tissue was ground in liquid nitrogen and protein was extracted using a buffer containing 100 mm Tris‐HCl, pH 7.2, 150 mm NaCl, 5 mm EDTA, 5% SDS, 2 m urea, 10 mm DTT and 1% (v/v) Protease Inhibitor Cocktail (P9599, Sigma‐Aldrich), boiled for 10 min and debris removed by centrifugation for 2 min at 12,000g. Protein samples were separated on an 8% or 12% (pMAPK) sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS‐PAGE) and blotted on polyvinylidene difluoride (PVDF) membrane (Thermo Fisher Scientific). Immunoblotting were performed with antibodies diluted in blocking solution [5% nonfat milk in TBS with 0.1% (v/v) Tween‐20] at the following dilutions: α‐HA‐horseradish peroxidase (HRP) antibody (3F10, Roche), 1 : 2000; α‐p44/42‐ERK (Cell Signaling Technology), 1 : 3000. Blots were developed with Pierce ECL pico Western Blotting substrate (Thermo Fisher Scientific). Equal loading of protein was determined by Coomassie Brilliant Blue staining of the blotted membrane.
Author contributions
SP, JGM and CZ designed the study. MS transformed the plants. AM and NP did the pathogen infection experiments. SP and JG performed all other experiments with technical assistance and advice from SR, LS and JAD. SP, JGM and CZ wrote the manuscript with input from all authors.
Supporting information
Figure S1 EFR expression does not affect development of M. truncatula.
Figure S2 Alignment of elf18 peptide sequences.
Figure S3 M. truncatula responds to flg22 peptide.
Figure S4 MAPK activation and marker gene induction is induced upon elf18 treatment in EFR‐expressing lines.
Figure S5 Transgenic EFR‐Medicago roots and nodules accumulate EFR.
Figure S6 EFR expression in M. truncatula provides quantitative resistance against the pathogen R. solanacearum.
Figure S7 Dose‐dependent ROS response of M. truncatula roots from EFR‐expressing lines 26‐8 and 18‐1 to elf18 peptide.
Acknowledgements
The authors thank technical assistance from the John Innes Centre Horticultural Services, and helpful discussions with members of the Zipfel and Malone laboratories. SP is funded by a studentship from the Norwich Research Park. Research in the Malone and Zipfel laboratories is supported by BBSRC Institute Strategic Program Grant BB/J004553/1. The Zipfel laboratory is further supported by the Gatsby Charitable Foundation. The work done at LIPM, France was supported by the Laboratoire d'Excellence (LABEX) TULIP (ANR‐10‐LABX‐41). JAD thanks the John Innes Foundation for an Emeritus fellowship. Dr Zipfel collaborates with the 2Blades Foundation to develop commercial and charitable applications of plant disease resistance mediated by pattern recognition receptors. Dr Zipfel is an inventor on a patent filing on the EFR gene.
References
- Albert, I. , Bohm, H. , Albert, M. , Feiler, C.E. , Imkampe, J. , Wallmeroth, N. , Brancato, C. et al (2015) An RLP23‐SOBIR1‐BAK1 complex mediates NLP‐triggered immunity. Nat. Plants, 1, 15140. [DOI] [PubMed] [Google Scholar]
- Aslam, S.N. , Newman, M.A. , Erbs, G. , Morrissey, K.L. , Chinchilla, D. , Boller, T. , Jensen, T.T. et al (2008) Bacterial polysaccharides suppress induced innate immunity by calcium chelation. Curr. Biol. 18, 1078–1083. [DOI] [PubMed] [Google Scholar]
- Bahar, O. , Mordukhovich, G. , Luu, D.D. , Schwessinger, B. , Daudi, A. , Jehle, A.K. , Felix, G. et al (2016) Bacterial outer membrane vesicles induce plant immune responses. Mol. Plant Microbe Interact. 29, 374–384. [DOI] [PubMed] [Google Scholar]
- Berrabah, F. , Bourcy, M. , Eschstruth, A. , Cayrel, A. , Guefrachi, I. , Mergaert, P. , Wen, J. et al (2014) A nonRD receptor‐like kinase prevents nodule early senescence and defense‐like reactions during symbiosis. New Phytol. 203, 1305–1314. [DOI] [PubMed] [Google Scholar]
- Berrabah, F. , Ratet, P. and Gourion, B. (2015) Multiple steps control immunity during the intracellular accommodation of rhizobia. J. Exp. Bot. 66, 1977–1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berrabah, F. , Balliau, T. , Aït‐Salem, E.H. , George, J. , Zivy, M. , Ratet, P. and Gourion, B. (2018) Control of the ethylene signaling pathway prevents plant defenses during intracellular accommodation of the rhizobia. New Phytol. 219, 310–323. [DOI] [PubMed] [Google Scholar]
- Bladergroen, M.R. , Badelt, K. and Spaink, H.P. (2003) Infection‐blocking genes of a symbiotic Rhizobium leguminosarum strain that are involved in temperature‐dependent protein secretion. Mol. Plant Microbe Interact. 16, 53–64. [DOI] [PubMed] [Google Scholar]
- Boller, T. and Felix, G. (2009) A renaissance of elicitors: perception of microbe‐associated molecular patterns and danger signals by pattern‐recognition receptors. Annu. Rev. Plant Biol. 60, 379–406. [DOI] [PubMed] [Google Scholar]
- Bourcy, M. , Brocard, L. , Pislariu, C.I. , Cosson, V. , Mergaert, P. , Tadege, M. , Mysore, K.S. et al (2013) Medicago truncatula DNF2 is a PI‐PLC‐XD‐containing protein required for bacteroid persistence and prevention of nodule early senescence and defense‐like reactions. New Phytol. 197, 1250–1261. [DOI] [PubMed] [Google Scholar]
- Boutrot, F. and Zipfel, C. (2017) Function, discovery, and exploitation of plant pattern recognition receptors for broad‐spectrum disease resistance. Annu. Rev. Phytopathol. 55, 257–286. [DOI] [PubMed] [Google Scholar]
- Breakspear, A. , Liu, C. , Roy, S. , Stacey, N. , Rogers, C. , Trick, M. , Morieri, G. et al (2014) The root hair “infectome” of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for auxin signaling in rhizobial infection. Plant Cell, 26, 4680–4701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao, Y. , Halane, M.K. , Gassmann, W. and Stacey, G. (2017) The role of plant innate immunity in the legume‐rhizobium symbiosis. Annu. Rev. Plant Biol. 68, 535–561. [DOI] [PubMed] [Google Scholar]
- Chen, T. , Duan, L. , Zhou, B. , Yu, H. , Zhu, H. , Cao, Y. and Zhang, Z. (2017) Interplay of pathogen‐induced defense responses and symbiotic establishment in Medicago truncatula . Front. Microbiol. 8, Article 973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosson, V. , Durand, P. , d'Erfurth, I. , Kondorosi, A. and Ratet, P. (2006) Medicago truncatula transformation using leaf explants. Methods Mol. Biol. 343, 115–127. [DOI] [PubMed] [Google Scholar]
- Dai, W.J. , Zeng, Y. , Xie, Z.P. and Staehelin, C. (2008) Symbiosis‐promoting and deleterious effects of NopT, a novel type 3 effector of Rhizobium sp. strain NGR234. J. Bacteriol. 190, 5101–5110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dangl, J.L. , Horvath, D.M. and Staskawicz, B.J. (2013) Pivoting the plant immune system from dissection to deployment. Science, 341, 746–751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Haeze, W. and Holsters, M. (2004) Surface polysaccharides enable bacteria to evade plant immunity. Trends Microbiol. 12, 555–561. [DOI] [PubMed] [Google Scholar]
- Domonkos, A. , Horvath, B. , Marsh, J.F. , Halasz, G. , Ayaydin, F. , Oldroyd, G.E. and Kalo, P. (2013) The identification of novel loci required for appropriate nodule development in Medicago truncatula . BMC Plant Biol. 13, 157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du, J. , Verzaux, E. , Chaparro‐Garcia, A. , Bijsterbosch, G. , Keizer, L.C. , Zhou, J. , Liebrand, T.W. et al (2015) Elicitin recognition confers enhanced resistance to Phytophthora infestans in potato. Nat. Plants, 1, 15034. [DOI] [PubMed] [Google Scholar]
- Felix, G. , Duran, J.D. , Volko, S. and Boller, T. (1999) Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 18, 265–276. [DOI] [PubMed] [Google Scholar]
- Ferguson, B.J. , Mens, C. , Hastwell, A.H. , Zhang, M. , Su, H. , Jones, C.H. , Chu, X. et al (2018) Legume nodulation: the host controls the party. Plant, Cell Environ. 1–11. 10.1111/pce.13348 [DOI] [PubMed] [Google Scholar]
- Ge, Y.‐Y. , Xiang, Q.‐W. , Wagner, C. , Zhang, D. , Xie, Z.‐P. and Staehelin, C. (2016) The type 3 effector NopL of Sinorhizobium sp. strain NGR234 is a mitogen‐activated protein kinase substrate. J. Exp. Bot. 67, 2483–2494. [DOI] [PubMed] [Google Scholar]
- Gibson, K.E. , Kobayashi, H. and Walker, G.C. (2008) Molecular determinants of a symbiotic chronic infection. Annu. Rev. Genet. 42, 413–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gourion, B. , Berrabah, F. , Ratet, P. and Stacey, G. (2015) Rhizobium‐legume symbioses: the crucial role of plant immunity. Trends Plant Sci. 20, 186–194. [DOI] [PubMed] [Google Scholar]
- Hacquard, S. , Spaepen, S. , Garrido‐Oter, R. and Schulze‐Lefert, P. (2017) Interplay between innate immunity and the plant microbiota. Annu. Rev. Phytopathol. 55, 565–589. [DOI] [PubMed] [Google Scholar]
- Holton, N. , Nekrasov, V. , Ronald, P.C. and Zipfel, C. (2015) The phylogenetically‐related pattern recognition receptors EFR and XA21 recruit similar immune signaling components in monocots and dicots. PLoS Pathog. 11, e1004602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones, K.M. , Lloret, J. , Daniele, J.R. and Walker, G.C. (2007) The type IV secretion system of Sinorhizobium meliloti strain 1021 is required for conjugation but not for intracellular symbiosis. J. Bacteriol. 189, 2133–2138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones, K.M. , Sharopova, N. , Lohar, D.P. , Zhang, J.Q. , VandenBosch, K.A. and Walker, G.C. (2008) Differential response of the plant Medicago truncatula to its symbiont Sinorhizobium meliloti or an exopolysaccharide‐deficient mutant. Proc. Natl Acad. Sci. USA, 105, 704–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawaharada, Y. , Kelly, S. , Nielsen, M.W. , Hjuler, C.T. , Gysel, K. , Muszynski, A. , Carlson, R.W. et al (2015) Receptor‐mediated exopolysaccharide perception controls bacterial infection. Nature, 523, 308–312. [DOI] [PubMed] [Google Scholar]
- Kazemi‐Pour, N. , Condemine, G. and Hugouvieux‐Cotte‐Pattat, N. (2004) The secretome of the plant pathogenic bacterium Erwinia chrysanthemi . Proteomics, 4, 3177–3186. [DOI] [PubMed] [Google Scholar]
- Kunze, G. , Zipfel, C. , Robatzek, S. , Niehaus, K. , Boller, T. and Felix, G. (2004) The N terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. Plant Cell, 16, 3496–3507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacombe, S. , Rougon‐Cardoso, A. , Sherwood, E. , Peeters, N. , Dahlbeck, D. , van Esse, H.P. , Smoker, M. et al (2010) Interfamily transfer of a plant pattern‐recognition receptor confers broad‐spectrum bacterial resistance. Nat. Biotechnol. 28, 365–369. [DOI] [PubMed] [Google Scholar]
- Lee, H.‐A. , Lee, H.‐Y. , Seo, E. , Lee, J. , Kim, S.‐B. , Oh, S. , Choi, E. et al (2017) Current understandings of plant nonhost resistance. Mol. Plant Microbe Interact. 30, 5–15. [DOI] [PubMed] [Google Scholar]
- Leong, S.A. , Williams, P.H. and Ditta, G.S. (1985) Analysis of the 5’ regulatory region of the gene for delta‐aminolevulinic acid synthetase of Rhizobium meliloti . Nucleic Acids Res. 13, 5965–5976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang, Y. , Cao, Y.R. , Tanaka, K. , Thibivilliers, S. , Wan, J.R. , Choi, J. , Kang, C.H. et al (2013) Nonlegumes respond to rhizobial Nod factors by suppressing the innate immune response. Science, 341, 1384–1387. [DOI] [PubMed] [Google Scholar]
- Libault, M. , Farmer, A. , Brechenmacher, L. , Drnevich, J. , Langley, R.J. , Bilgin, D.D. , Radwan, O. et al (2010) Complete transcriptome of the soybean root hair cell, a single‐cell model, and its alteration in response to Bradyrhizobium japonicum infection. Plant Physiol. 152, 541–552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lohar, D.P. , Sharopova, N. , Endre, G. , Penuela, S. , Samac, D. , Town, C. , Silverstein, K.A. et al (2006) Transcript analysis of early nodulation events in Medicago truncatula . Plant Physiol. 140, 221–234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez‐Gomez, M. , Sandal, N. , Stougaard, J. and Boller, T. (2012) Interplay of flg22‐induced defence responses and nodulation in lotus japonicus. J. Exp. Bot. 63, 393–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu, F. , Wang, H. , Wang, S. , Jiang, W. , Shan, C. , Li, B. , Yang, J. et al (2015) Enhancement of innate immune system in monocot rice by transferring the dicotyledonous elongation factor Tu receptor EFR. J. Integr. Plant Biol. 57, 641–652. [DOI] [PubMed] [Google Scholar]
- Macho, A.P. and Zipfel, C. (2015) Targeting of plant pattern recognition receptor‐triggered immunity by bacterial type‐III secretion system effectors. Curr. Opin. Microbiol. 23, 14–22. [DOI] [PubMed] [Google Scholar]
- Madsen, L.H. , Tirichine, L. , Jurkiewicz, A. , Sullivan, J.T. , Heckmann, A.B. , Bek, A.S. , Ronson, C.W. et al (2010) The molecular network governing nodule organogenesis and infection in the model legume Lotus japonicus . Nat. Commun. 1, 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansfield, J. , Genin, S. , Magori, S. , Citovsky, V. , Sriariyanum, M. , Ronald, P. , Dow, M. et al (2012) Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 13, 614–629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendes, B.M.J. , Cardoso, S.C. , Boscariol‐Camargo, R.L. , Cruz, R.B. , Mourão Filho, F.A.A. and Bergamin Filho, A. (2010) Reduction in susceptibility to Xanthomonas axonopodis pv. citri in transgenic Citrus sinensis expressing the rice Xa21 gene. Plant. Pathol. 59, 68–75. [Google Scholar]
- Michelmore, R. , Coaker, G. , Bart, R. , Beattie, G. , Bent, A. , Bruce, T. , Cameron, D. et al (2017) Foundational and translational research opportunities to improve plant health. Mol. Plant Microbe Interact. 30, 515–516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mithofer, A. , Bhagwat, A.A. , Feger, M. and Ebel, J. (1996) Suppression of fungal beta‐glucan‐induced plant defence in soybean (Glycine max l) by cyclic 1,3‐1,6‐beta‐glucans from the symbiont Bradyrhizobium japonicum . Planta, 199, 270–275. [Google Scholar]
- Müller, D.B. , Vogel, C. , Bai, Y. and Vorholt, J.A. (2016) The plant microbiota: systems‐level insights and perspectives. Annu. Rev. Genet. 50, 211–234. [DOI] [PubMed] [Google Scholar]
- Murray, J.D. , Karas, B.J. , Sato, S. , Tabata, S. , Amyot, L. and Szczyglowski, K. (2007) A cytokinin perception mutant colonized by Rhizobium in the absence of nodule organogenesis. Science, 315, 101–104. [DOI] [PubMed] [Google Scholar]
- Muszynski, A. , Heiss, C. , Hjuler, C.T. , Sullivan, J.T. , Kelly, S.J. , Thygesen, M.B. , Stougaard, J. et al (2016) Structures of exopolysaccharides involved in receptor‐mediated perception of Mesorhizobium loti by Lotus japonicus . J. Biol. Chem. 291, 20946–20961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson, M.S. and Sadowsky, M.J. (2015) Secretion systems and signal exchange between nitrogen‐fixing rhizobia and legumes. Front. Plant Sci. 6, Article 491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson, M.S. , Chun, C.L. and Sadowsky, M.J. (2016) Type IV effector proteins involved in the medicago‐sinorhizobium symbiosis. Mol. Plant Microbe Interact. 30, 28–34. [DOI] [PubMed] [Google Scholar]
- Niehaus, K. , Kapp, D. and Pühler, A. (1993) Plant defence and delayed infection of pseudonodules induced by an exopolysaccharide (EPS I)‐deficient Rhizobium meliloti mutant. Planta, 190, 415–425. [Google Scholar]
- Oldroyd, G.E.D. , Murray, J.D. , Poole, P.S. and Downie, J.A. (2011) The rules of engagement in the legume‐rhizobial symbiosis. Annu. Rev. Genet. 45, 119–144. [DOI] [PubMed] [Google Scholar]
- Pfeilmeier, S. , Caly, D.L. and Malone, J.G. (2016) Bacterial pathogenesis of plants: future challenges from a microbial perspective: challenges in bacterial molecular plant pathology. Mol. Plant Pathol. 17, 1298–1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price, P.A. , Tanner, H.R. , Dillon, B.A. , Shabab, M. , Walker, G.C. and Griffitts, J.S. (2015) Rhizobial peptidase HrrP cleaves host‐encoded signaling peptides and mediates symbiotic compatibility. Proc. Natl Acad. Sci. USA, 112, 15244–15249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pruitt, R.N. , Schwessinger, B. , Joe, A. , Thomas, N. , Liu, F. , Albert, M. , Robinson, M.R. et al (2015) The rice immune receptor XA21 recognizes a tyrosine‐sulfated protein from a Gram‐negative bacterium. Sci. Adv. 1, e1500245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radutoiu, S. , Madsen, L.H. , Madsen, E.B. , Felle, H.H. , Umehara, Y. , Grønlund, M. , Sato, S. et al (2003) Plant recognition of symbiotic bacteria requires two LysM receptor‐like kinases. Nature, 425, 585. [DOI] [PubMed] [Google Scholar]
- Remigi, P. , Anisimova, M. , Guidot, A. , Genin, S. and Peeters, N. (2011) Functional diversification of the Gala type III effector family contributes to Ralstonia solanacearum adaptation on different plant hosts. New Phytol. 192, 976–987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez‐Moreno, L. , Song, Y. and Thomma, B.P.H.J. (2017) Transfer and engineering of immune receptors to improve recognition capacities in crops. Curr. Opin. Plant Biol. 38, 42–49. [DOI] [PubMed] [Google Scholar]
- Ryu, H. , Laffont, C. , Frugier, F. and Hwang, I. (2017) MAP kinase‐mediated negative regulation of symbiotic nodule formation in Medicago truncatula . Mol. Cells, 40, 17–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoonbeek, H.J. , Wang, H.H. , Stefanato, F.L. , Craze, M. , Bowden, S. , Wallington, E. , Zipfel, C. et al (2015) Arabidopsis EF‐Tu receptor enhances bacterial disease resistance in transgenic wheat. New Phytol. 206, 606–613. [DOI] [PubMed] [Google Scholar]
- Schwessinger, B. , Bahar, O. , Thomas, N. , Holton, N. , Nekrasov, V. , Ruan, D. , Canlas, P.E. et al (2015) Transgenic expression of the dicotyledonous pattern recognition receptor EFR in rice leads to ligand‐dependent activation of defense responses. PLoS Pathog. 11, e1004809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skorpil, P. , Saad, M.M. , Boukli, N.M. , Kobayashi, H. , Ares‐Orpel, F. , Broughton, W.J. and Deakin, W.J. (2005) NopP, a phosphorylated effector of Rhizobium sp. strain NGR234, is a major determinant of nodulation of the tropical legumes Flemingia congesta and Tephrosia vogelii . Mol. Microbiol. 57, 1304–1317. [DOI] [PubMed] [Google Scholar]
- Staehelin, C. and Krishnan, H.B. (2015) Nodulation outer proteins: double‐edged swords of symbiotic rhizobia. Biochem. J. 470, 263–274. [DOI] [PubMed] [Google Scholar]
- Stringlis, I.A. , Proietti, S. , Hickman, R. , Van Verk, M.C. , Zamioudis, C. and Pieterse, C.M.J. (2018) Root transcriptional dynamics induced by beneficial rhizobacteria and microbial immune elicitors reveal signatures of adaptation to mutualists. Plant J. 93, 166–180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugawara, M. , Epstein, B. , Badgley, B.D. , Unno, T. , Xu, L. , Reese, J. , Gyaneshwar, P. et al (2013) Comparative genomics of the core and accessory genomes of 48 Sinorhizobium strains comprising five genospecies. Genome Biol. 14, R17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tellstroem, V. , Usadel, B. , Thimm, O. , Stitt, M. , Kuester, H. and Niehaus, K. (2007) The lipopolysaccharide of Sinorhizobium meliloti suppresses defense‐associated gene expression in cell cultures of the host plant Medicago truncatula . Plant Physiol. 143, 825–837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tirichine, L. , Imaizumi‐Anraku, H. , Yoshida, S. , Murakami, Y. , Madsen, L.H. , Miwa, H. , Nakagawa, T. et al (2006) Deregulation of a Ca2+/calmodulin‐dependent kinase leads to spontaneous nodule development. Nature, 441, 1153–1156. [DOI] [PubMed] [Google Scholar]
- Trinick, M.J. , Dilworth, M.J. and Grounds, M. (1976) Factors affecting reduction of acetylene by root‐nodules of Lupinus species. New Phytol. 77, 359–370. [Google Scholar]
- Tripathi, J.N. , Lorenzen, J. , Bahar, O. , Ronald, P. and Tripathi, L. (2014) Transgenic expression of the rice Xa21 pattern‐recognition receptor in banana (Musa sp.) confers resistance to Xanthomonas campestris pv. musacearum . Plant Biotechnol. J. 12, 663–673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Udvardi, M. and Poole, P.S. (2013) Transport and metabolism in legume‐rhizobia symbioses. Annu. Rev. Plant Biol. 64, 781–805. [DOI] [PubMed] [Google Scholar]
- Vailleau, F. , Sartorel, E. , Jardinaud, M.F. , Chardon, F. , Genin, S. , Huguet, T. , Gentzbittel, L. et al (2007) Characterization of the interaction between the bacterial wilt pathogen Ralstonia solanacearum and the model legume plant Medicago truncatula . Mol. Plant Microbe Interact. 20, 159–167. [DOI] [PubMed] [Google Scholar]
- Wang, C. , Yu, H. , Luo, L. , Duan, L. , Cai, L. , He, X. , Wen, J. et al (2016) NODULES WITH ACTIVATED DEFENSE 1 is required for maintenance of rhizobial endosymbiosis in Medicago truncatula . New Phytol. 212, 176–191. [DOI] [PubMed] [Google Scholar]
- Watt, S.A. , Wilke, A. , Patschkowski, T. and Niehaus, K. (2005) Comprehensive analysis of the extracellular proteins from Xanthomonas campestris pv. campestris B100. Proteomics, 5, 153–167. [DOI] [PubMed] [Google Scholar]
- Whitney, J.C. , Quentin, D. , Sawai, S. , LeRoux, M. , Harding, B.N. , Ledvina, H.E. , Tran, B.Q. et al (2015) An interbacterial NAD(P)(+) glycohydrolase toxin requires elongation factor Tu for delivery to target cells. Cell, 163, 607–619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xin, D.‐W. , Liao, S. , Xie, Z.‐P. , Hann, D.R. , Steinle, L. , Boller, T. and Staehelin, C. (2012) Functional analysis of NopM, a novel E3 ubiquitin ligase (NEL) domain effector of Rhizobium sp. strain NGR234. PLoS Pathog. 8, e1002707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zgadzaj, R. , James, E.K. , Kelly, S. , Kawaharada, Y. , de Jonge, N. , Jensen, D.B. , Madsen, L.H. et al (2015) A legume genetic framework controls infection of nodules by symbiotic and endophytic bacteria. PLoS Genet. 11, e1005280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zipfel, C. and Oldroyd, G.E. (2017) Plant signalling in symbiosis and immunity. Nature, 543, 328–336. [DOI] [PubMed] [Google Scholar]
- Zipfel, C. , Kunze, G. , Chinchilla, D. , Caniard, A. , Jones, J.D. , Boller, T. and Felix, G. (2006) Perception of the bacterial PAMP EF‐Tu by the receptor EFR restricts Agrobacterium‐mediated transformation. Cell, 125, 749–760. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1 EFR expression does not affect development of M. truncatula.
Figure S2 Alignment of elf18 peptide sequences.
Figure S3 M. truncatula responds to flg22 peptide.
Figure S4 MAPK activation and marker gene induction is induced upon elf18 treatment in EFR‐expressing lines.
Figure S5 Transgenic EFR‐Medicago roots and nodules accumulate EFR.
Figure S6 EFR expression in M. truncatula provides quantitative resistance against the pathogen R. solanacearum.
Figure S7 Dose‐dependent ROS response of M. truncatula roots from EFR‐expressing lines 26‐8 and 18‐1 to elf18 peptide.