
Figure 3.
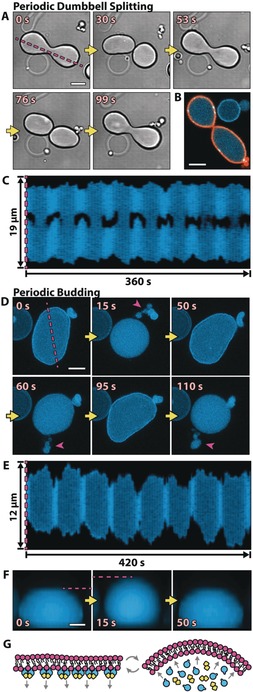
The vesicle shapes change in concert with Min oscillations. A) Time series of DIC images of a dumbbell‐shaped GUV that repetitively splits into two compartments connected by a narrow membrane neck. B) The confocal image of a shape‐oscillating GUV with fluorescently labeled lipids (orange) shows that the GUV does not undergo a full fission into two separate GUVs, but rather that the two compartments are still connected by a membrane neck. C) Kymograph along the line parallel to the axis of rotational symmetry of a dynamic GUV (see the magenta line in (A)). Whenever MinD is in the lumen, the GUV is split into two, almost separate compartments. Movie S6 features all three vesicle examples in (A) to (C). D) A GUV that undergoes periodic budding and subsequent fusion of the buds with the mother vesicle. Buds are highlighted with magenta arrows. Shown are z projections of five confocal planes. E) Kymograph (see the magenta line in (D)) of a repetitively budding vesicle, showing the reduction in size in the x and y dimensions of a GUV whenever budding occurs. F) Sequential images of a side view of a periodically budding vesicle, showing that the reduction in size in the x–y plane is accompanied by an increase in size in the z direction (see also Figure S6 C). Movie S7 shows the vesicles from (D), (E), and (F) sequentially. Scale bars: 5 μm. G) Suspected mechanism: Membrane‐bound MinD increases the surface area of the inner membrane leaflet. Upon protein detachment, the intrinsic curvature of the bilayer increases, and membrane deformations occur.