
Fig. 4.
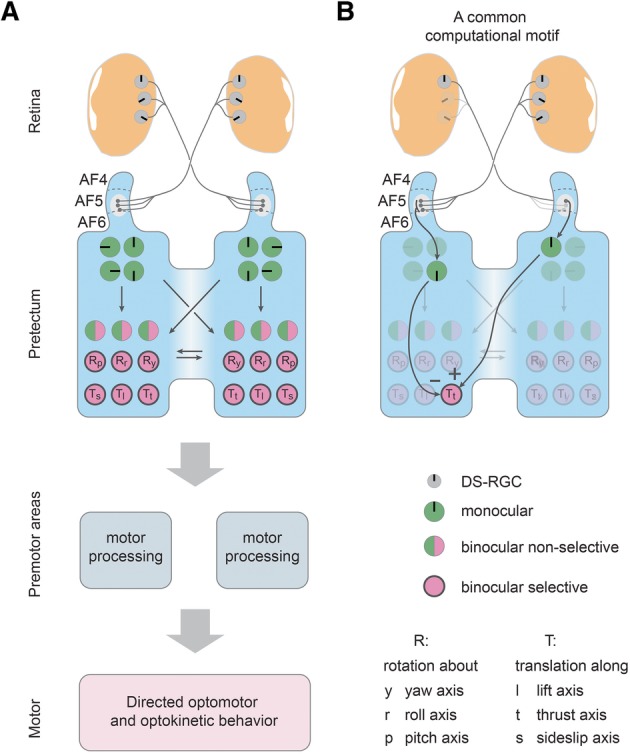
Proposed circuit model for binocular processing of optic flow in the pretectum. a The pretectum receives its DS-RGC inputs mainly via the pretectal arborization field AF5 (F. Kubo, personal communication, October 2018) and a transformation of represented motion axes occurs between retina (three PDs [14, 15]) and pretectum (four PDs). The monocularly responsive pretectal DS cells (green) send commissural projections (via the posterior commissure) to the contralateral pretectum, resulting in binocular non-selective response types (half green/half magenta) and binocular selective (magenta) response types. The binocular-selective neurons code for a single rotational or translational optic flow direction and could directly drive appropriate optomotor and optokinetic behavior via premotor structures such as the nucleus of the medial longitudinal fasciculus and oculomotor nuclei. Rp, Rr, Ry: neurons selective for rotations about the pitch, roll and yaw axes, respectively. Ts, Tt, Tl: neurons selective for translations along the sideslip (sidewards left/right), thrust (forward/backward), and lift (up/down) axes, respectively. Please note that only these six axes were tested in this study and it is possible that other (oblique) global optic flow axes result in even stronger responses, as suggested by previous reports on other species (see “Discussion”). b A proposed common computational motif shared across different planes of motion is antagonistic inputs from monocular pretectal neurons to binocular-selective neurons coding for motion in the same plane (here in the horizontal plane): A left eye monocular neuron in the right pretectal half excites the Forward-selective neuron (Tt) during forward translation, while a right eye monocular neuron in the left pretectal half inhibits the selective neuron during nasal-to-temporal motion in the right eye, thus establishing forward selectivity