
Figure 4. Model of P2RY8-mediated B cell confinement and proposed distribution of B cell guidance cues.
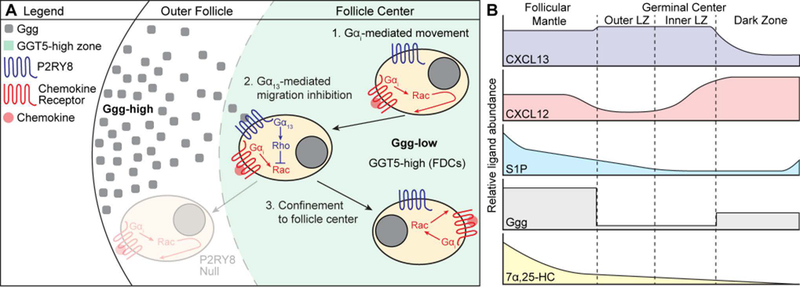
(A) A gradient of Ggg is formed through GGT5-mediated degradation of Ggg at the center of the follicle, resulting in higher levels of Ggg in the outer follicle and lower levels at the center. When P2RY8-expressing B cells experience movement (either random or chemokine-directed) towards outer areas of the follicle, Ggg engages P2RY8 to induce Gα13-mediated activation of Rho, which inhibits Rac-based movement. This confines a P2RY8-expressing cell to the center of the follicle. Movement towards the center of the follicle is not inhibited by Ggg, promoting central clustering. The distribution of chemokine (CXCL13, CXCL12 – red dot) is not shown. (B) Proposed model of B cell guidance cue distribution within GC-containing follicles. We speculate that the high density of GGT5 expression within GCs (being higher on LZ than DZ FDCs) results in a sharp, stepwise drop in Ggg levels. In contrast, degradation of S1P does not depend on FDCs, but may be mediated by many cell types (including B cells), suggesting that the S1P gradient may decay in a more linear fashion. The interplay between these two migration-inhibitory ligands may help support the formation of inner and outer zones in the GC LZ that have been reported in tonsil GCs (117, 145). CXCL13 is present throughout the follicular mantle (region around the GC) and is abundant within the LZ, while CXCL12 is enriched in the DZ. As suggested by Figure 2, the 7α,25-HC gradient is high in the follicular mantle and low at the follicle center. These cues together support GC confinement and zonal organization.