

Abstract
The auditory system's ability to interpret sounds over a wide range of amplitudes rests on the nonlinear responsiveness of the ear. Whether measured by basilar-membrane vibration, nerve-fiber activity, or perceived loudness, the ear is most sensitive to small signals and grows progressively less responsive as stimulation becomes stronger. Seeking a correlate of this behavior at the level of mechanoelectrical transduction, we examined the responses of hair bundles to direct mechanical stimulation. As reported by the motion of an attached glass fiber, an active hair bundle from the bullfrog's sacculus oscillates spontaneously. Sinusoidal movement of the fiber's base by as little as ±1 nm, corresponding to the application at the bundle's top of a force of ±0.3 pN, causes detectable phase-locking of the bundle's oscillations to the stimulus. Although entrainment increases as the stimulus grows, the amplitude of the hair-bundle movement does not rise until phase-locking is nearly complete. A bundle is most sensitive to stimulation at its frequency of spontaneous oscillation. Far from that frequency, the sensitivity of an active hair bundle resembles that of a passive bundle. Over most of its range, an active hair bundle's response grows as the one-third power of the stimulus amplitude; the bundle's sensitivity declines accordingly in proportion to the negative two-thirds power of the excitation. This scaling behavior, also found in the response of the mammalian basilar membrane to sound, signals the operation of an amplificatory process at the brink of an oscillatory instability, a Hopf bifurcation.
Hearing operates over a remarkably broad dynamic range. The faintest sounds we can sense impart to the ear an amount of energy per cycle of oscillation comparable to thermal energy (1). At the other extreme, the ear endures urban, industrial, and military noise that represents sound pressures 6 orders of magnitude greater than threshold!
To accommodate such diverse stimuli, the ear's responsiveness must be nonlinear. The common use of decibels, a logarithmic metric, for the intensity of sound reflects this fact. Nonlinear compression of responsiveness is evident throughout the ear's transduction process. The perceived loudness, the firing rate of auditory-nerve axons, and the basilar-membrane vibration in the cochlea all grow more gradually than the input signal. In the chinchilla's cochlea, for example, sound-pressure levels ranging over 120 dB are represented as basilar-membrane vibrations spanning but 2 orders of magnitude (ref. 2; reviewed in ref. 3). The basilar membrane's responsiveness, defined as the increase in amplitude of vibration evoked by an increment in sound-pressure level, is greatest for threshold stimuli and then declines progressively at moderate to intense levels.
The nonlinearity of aural responsiveness is associated closely with the ear's great sensitivity, sharp frequency selectivity, and ability to generate otoacoustic emissions. These four characteristics of hearing result from an active process that mechanically amplifies sound inputs (reviewed in refs. 4 and 5). Although the active process of the mammalian cochlea is widely believed to involve voltage-dependent cell-body movements by outer hair cells (reviewed in refs. 4 and 6), the ears of nonmammalian tetrapods lack those cells and presumably the phenomenon of membrane-based electromotility. Because these animals nevertheless display highly sensitive and sharply tuned hearing as well as aural nonlinearities and otoacoustic emissions, their ears must employ some other active process (ref. 7; reviewed in refs. 5 and 8).
Studies on the ears of frogs and turtles have demonstrated that active hair-bundle movements can account for many of the properties of the active process including amplification, distortion-tone production, and spontaneous oscillation (reviewed in refs. 8–10). If hair-bundle motility underlies the ear's active process, bundle movements also should exhibit the nonlinear responsiveness characteristic of hearing. To seek this nonlinearity, we therefore have studied the mechanical responses of individual hair bundles from the bullfrog to stimuli ranging in amplitude from near threshold to an almost saturating level.
Materials and Methods
Experimental Preparation and Recording Procedures.
Experiments were performed on hair bundles from the bullfrog's sacculus with published techniques (11, 12). The accompanying paper (13) describes the dissection, mounting, and maintenance of the experimental preparation as well as the conditions for stimulating and recording from hair bundles.
Hair-Bundle Motion.
We denote by X(t) the displacement of a hair bundle's top relative to its mean position after subtraction of drift. The power spectrum of the bundle's motion is
 |
1 |
in which
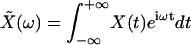 |
is the Fourier transform of X(t) and tEXP is the width of the experimental time window (13). The root-mean-square (RMS) magnitude of the bundle movement over this time window can be computed as
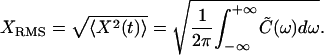 |
2 |
Phase-Locking to a Sinusoidal Stimulus.
Each hair bundle was stimulated with a flexible glass fiber attached by its tip to the bundle's top. Sinusoidal displacement of the fiber's base, Δ(t), evoked a peak, S̃(ω), in the power spectrum of bundle motion, which reflected partial entrainment of a hair bundle's oscillations at the angular frequency of stimulation, ωS. This peak could be extracted from the bundle's power spectrum through the relation
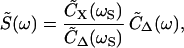 |
3 |
in which C̃Δ(ω) represents the power spectrum of the stimulus fiber's base displacement.
Because small stimuli produced incomplete entrainment, we defined the degree of phase locking as
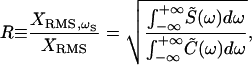 |
4 |
in which XRMS,ωs is the RMS magnitude of the bundle's response at the stimulus frequency and 0 ≤ R ≤ 1.
Hair-Bundle Sensitivity.
The sinusoidal movement, Δ(t), of the fiber's base generated a Fourier component of bundle displacement, <X̃(ωs)>, at the frequency of stimulation. The same phase-locked response would have resulted from a hypothetical periodic force, f(t), applied directly to the bundle's top in the presence of a fiber, the base of which is held at a fixed position. The Fourier amplitude of this force at the stimulus frequency is given by ref. 13 as
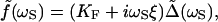 |
5 |
in which KF ≅ 200–300 μN⋅m−1 is the fiber's stiffness and ξ ≅ 80–120 nN⋅s⋅m−1 is a drag coefficient describing the effect of viscous forces. The force f can be used to define a response function at the frequency of stimulation for the combined system of a hair bundle and attached fiber:
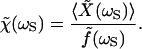 |
6 |
This definition remains valid even when the response is nonlinear. The modulus of the response function, which has the units of a compliance, represents the hair bundle's sensitivity at the frequency of stimulation. The response function's phase reflects the phase of the hair bundle's movement with respect to that of the stimulus.
Results
Spontaneous Hair-Bundle Oscillation.
To examine the mechanical responses of individual hair bundles, we mounted an epithelial preparation of the bullfrog's sacculus in a two-compartment experimental chamber. We mimicked the ionic environment in vivo by exposing the basolateral surfaces of hair cells to standard saline solution while their mechanosensory hair bundles projected into a Ca2+-poor, endolymph-like fluid. Under these conditions, a majority of hair cells displayed spontaneous oscillations of their hair bundles (11–13).
The tip of a flexible glass fiber was attached atop a hair bundle, and its image was projected onto a pair of photodiodes, the output of which indicated the bundle's position. We observed spontaneous hair-bundle oscillations at 5–40 Hz with peak-to-peak magnitudes of 25–80 nm (Fig. 1A). These movements were noisy, fluctuating both in amplitude and phase. Because they violate the fluctuation-dissipation theorem, the spontaneous oscillations require the performance of work by an active hair bundle that draws on an energy source within the hair cell (13). This active process also endows the hair bundle with the ability to amplify its response to sinusoidal stimuli (11). To characterize the amplificatory process further, we measured the dependence of hair-bundle responses on the amplitude and frequency of stimulation.
Figure 1.

Entrainment of a hair bundle's spontaneous oscillation. (A) Although erratic in the absence of stimulation (top trace), a bundle's motion first became more regular and then increased in magnitude with growing stimulus amplitude. The power spectrum of bundle motion, shown to the right of each movement record, was dominated by the Fourier component at the frequency of stimulation. Note that a significant frequency-locked response was apparent already at a stimulus amplitude of 1 nm. Because the response at ±3 nm was not centered exactly at the frequency of stimulation, the bundle probably was driven near but not precisely at its natural frequency. A 9-Hz sinusoidal displacement applied at the stimulus fiber's base provided stimuli to the bundle's top with amplitudes of 0–60 nm. In each record, the stimulus trace (red, smooth curve) is superimposed on the response (black, irregular curve) at the identical gain. (B) A semilogarithmic plot relates the RMS magnitude of bundle motion to the stimulus amplitude. During stimulation near the bundle's natural frequency (black ●, 9 Hz; data of A), the magnitude of movement remained roughly constant for stimuli up to ±10 nm. For stimulation well above the natural frequency (purple ▴, 153 Hz), a relatively intense stimulus of ±50 nm was required to increase the bundle's magnitude of movement significantly. (C) This semilogarithmic plot portrays the degree of entrainment of a hair bundle's movement as a function of stimulus amplitude. When the bundle was stimulated near its natural frequency (black ○, 7 Hz; ●, 9 Hz), significant entrainment occurred for nanometer displacements of the stimulus fiber's base. For a stimulus of ±15 nm, the Fourier component at the frequency of stimulation encompassed more than 90% of the spectral power. In contrast, high-frequency stimuli (purple ▿, 70 Hz; ▴, 153 Hz) were comparatively inefficient: nanometer displacements failed to elicit significant entrainment. Each stimulus corresponded to 30 cycles of a sinusoidal oscillation of the fiber's base save at 9 Hz, for which the stimulus lasted 50 cycles. The spectral density of the unstimulated bundle's movements was centered at a frequency of 8 Hz. The stimulus fiber had a stiffness of 250 μN⋅m−1 and a drag coefficient of 120 nN⋅s⋅m−1.
Entrainment of Spontaneous Oscillations.
Sinusoidal displacement of the stimulus fiber's base at a frequency near that of a hair bundle's spontaneous oscillation rendered bundle movements more regular. A striking feature of our recordings was that oscillatory hair bundles responded to nanometer displacements of the fiber's base, which corresponded to forces of a fraction of a piconewton applied at the bundle's top (Eq. 6). In response to minute stimuli, a narrow peak at or near the frequency of stimulation emerged in the power spectrum of bundle motion (Fig. 1A). As the stimulus increased, the bundle's response at the stimulus frequency grew at the expense of other spectral components. The overall magnitude of bundle oscillation thus remained relatively constant until the stimulus had almost completely entrained the hair bundle's movement (Fig. 1 B and C).
By computing the fraction of a hair bundle's motion that was in phase with the stimulus as a function of the stimulus amplitude, we characterized the bundle's propensity to adjust the phase of its oscillation to that of a sinusoidal input. Phase-locking displayed a steep dependence on amplitude for small stimuli, increasing by 9% per nanometer of displacement of the stimulus fiber's base. A ±10-nm stimulus, corresponding to an applied force near ±3 pN, was intense enough to elicit almost complete entrainment (Fig. 1C).
Frequency Tuning of Responses.
A hair bundle was most sensitive to small stimuli when stimulated near its frequency of spontaneous oscillation, which therefore may be defined as the bundle's natural frequency. Stimulation at a higher frequency, for which the spectrum of spontaneous bundle motion displayed negligible power, evoked no significant entrainment for stimuli smaller than ±10 nm (Fig. 1C). The degree of entrainment increased for higher amplitudes but by only 1% per nanometer of displacement. Moreover, an intense stimulus of ±50 nm was required to entrain the movements wholly and to increase the magnitude of bundle motion significantly.
By measuring the dependence of the hair bundle's response function on stimulus frequency for a given stimulus amplitude, here 15 nm, we evaluated the tuning of the bundle's mechanical responsiveness (Fig. 2A). Stimulation near the natural frequency yielded a sensitivity ≈3-fold that for stimulation at higher frequencies, for which the sensitivity declined to a plateau. The bundle's frequency selectivity was modest, for the peak's sharpness exhibited a quality factor, or Q10 dB, of only 1.2. As a control, we also measured the responsiveness to sinusoidal stimuli of a passive hair cell that did not exhibit pronounced spontaneous oscillation (13). This bundle displayed no frequency selectivity; the sensitivity adopted a value of 1.2 km⋅N−1 for low-frequency stimulation and then gradually decreased with increasing stimulus frequency. This sensitivity corresponded to a hair-bundle stiffness of ≈1 mN⋅m−1, a value typical of the static bundle stiffness in the bullfrog's sacculus (14).
Figure 2.

Frequency tuning of the hair-bundle response. (A) The sensitivity of a spontaneously oscillatory hair bundle (●) peaked near the bundle's natural frequency and then declined slowly with increasing stimulus frequency. By contrast, a passive hair bundle (○), which did not display prominent spontaneous oscillations, maintained a low sensitivity throughout the frequency range. (B) The phase of the active hair bundle's movement (●) with respect to that of the stimulus varied from a phase lead at low frequencies to a phase lag at high frequencies, changing sign near the bundle's natural frequency. The passive bundle (○) instead displayed a slowly increasing phase lag throughout. The stimuli were of fixed amplitudes of 15 nm for the active hair bundle and 300 nm for the control cell. The stimulus fiber was identical to that used to obtain the data for Fig. 1.
The phase of the active hair bundle's response with respect to that of the sinusoidal stimulus crossed zero and changed its sign near the frequency of peak sensitivity (Fig. 2B). This behavior mirrored that of the sign of the external work required to entrain a hair bundle's oscillation (11). Note that the complex shapes of the measured sensitivity and phase curves accorded with a simple model for an active hair-bundle oscillator (13). The control cell displayed the expected passive behavior, for the hair bundle's movement lagged the stimulus at all frequencies.
Nonlinearity of Response.
When an active hair bundle was subjected to stimuli near its natural frequency and of progressively increasing amplitude, the Fourier component of the bundle motion at the stimulus frequency exhibited a compressive nonlinearity (Fig. 3A). The bundle's sensitivity was correspondingly greatest for small stimuli and declined with increasing stimulus amplitude (Fig. 3B). In response to intense stimuli, the response became linear, and the sensitivity reached a plateau. Stimulation at frequencies much higher than the bundle's natural frequency, in contrast, evoked linear responses of constant sensitivity throughout the range of amplitudes (Fig. 3 A and B). Moreover, this sensitivity did not vary markedly with stimulus frequency and equaled that of the bundle when stimulated at intense levels near its natural frequency.
Figure 3.

Nonlinear response compression for an active hair bundle. (A) Stimulation at high frequencies relative to the bundle's natural frequency (purple ▿, 70 Hz; ▴, 153 Hz) elicited essentially linear responses, corresponding to passive behavior. By contrast, the RMS magnitude of the bundle motion's Fourier component at the frequency of stimulation demonstrated strong amplification of small inputs at the natural frequency (●, 9 Hz). The abscissa represents the amplitude of the sinusoidal deflection applied at the base of the stimulus fiber. This hair bundle, the responses of which are depicted also in Figs. 1 and 2, displayed amplification even for stimuli as great as ±100 nm to ±150 nm. (B) When the hair bundle was driven near its natural frequency, its sensitivity was greatest for small inputs and declined to a plateau for more intense stimuli. At higher frequencies, the bundle maintained a constant, low sensitivity identical to that of the active bundle at the highest frequencies tested. The symbols are as described for A; only responses to stimuli greater than ±5 nm are shown. (C) This doubly logarithmic plot relates the magnitude of the phase-locked bundle response to the amplitude of motion at the stimulus fiber's base for five spontaneously oscillatory hair bundles. Three response regimes are evident. For stimuli only a few nanometers in amplitude, the responses were erratic but roughly linear (green line). Stimulation at slightly greater amplitudes and up to ≈ ±100 nm elicited nonlinear responses, the amplitudes of which grew as the one-third power of the stimulus amplitude (red line). The responses then approached a linear regime for still larger stimuli (green line). The data marked with open symbols represent results from four distinct hair bundles. The remaining data resulted from analyses of four data sets from the additional bundle, the responses of which are shown in Fig. 1; data from that illustration are marked with the symbol ●. (D) The hair-bundle sensitivity, plotted in doubly logarithmic form from the data in C, displayed saturation for displacements of the fiber's base by a few nanometers (green line). For moderate to intense levels of stimulation, the sensitivity entered a nonlinear regime, declining as the negative two-thirds power of the stimulus amplitude (red line). Finally, the sensitivity approached a constant, low value in response to intense stimuli (green line).
We collected responses from five spontaneously oscillating hair bundles, each stimulated at a frequency within 1 Hz of its natural frequency. Doubly logarithmic plots of each bundle's response magnitude (Fig. 3C) and sensitivity (Fig. 3D) at the stimulus frequency revealed three regimes. Noise dominated the response to stimuli of amplitudes smaller than 1–10 nm; the response appeared approximately linear, and the bundle's sensitivity generally remained in the range of 5–20 km⋅N−1. For moderate to intense stimuli, the bundle's response displayed a compressive nonlinearity. In this amplitude range of 15–150 nm, the dependence of the response magnitude, XRMS,ωs, on the stimulus amplitude, Δ, was well approximated by a power law, XRMS,ωs ∝ Δ1/3. The hair bundle's sensitivity correspondingly varied as Δ−2/3. Finally, for stimuli exceeding ±150 nm, the response approached linearity, and the sensitivity declined toward 1.2 km⋅N−1, the value characteristic of a passive hair bundle (Fig. 2A).
Discussion
The Hair-Bundle Amplifier.
What is the effect of the active process that powers spontaneous hair-bundle oscillation on a hair cell's mechanical responsiveness? A bundle's sensitivity is greatest for stimulation near the natural frequency and at a small amplitude. For stimulation at a substantially higher frequency, the sensitivity declines to a value similar to that of a passive cell that displays no spontaneous oscillation of its hair bundle (Fig. 2A). Even at the natural frequency, stimuli of large amplitude reduce an active hair bundle's sensitivity nearly to the value characteristic of a passive bundle (Fig. 3B).
Comparison of a hair bundle's active and passive behaviors provides a means of assessing the benefit of hair-bundle activity. The active process amplifies the phase-locked response to minute stimuli ≈10-fold (Fig. 3A). By so doing, it endows the hair bundle with sensitivity to forces of less than 1 pN, here corresponding to nanometer displacements of the fiber's base. The amplifier's gain may be defined as the ratio of a bundle's active to its passive sensitivity. The gain can be as great as 25, corresponding to 28 dB, for stimuli of ±1 nm, but averages ≈5 for small stimuli (Fig. 3D). The gain declines to unity for intense stimuli, which turn off or perhaps overwhelm the hair bundle's amplifier.
It is probable that the present results do not reflect the full extent of amplification of which hair bundles in the bullfrog's sacculus are capable. In addition to whatever damage the hair cells sustained during experimental preparation, two experimental conditions may not have been ideal. First, the mechanical loading of each hair bundle during recording, when a relatively fine stimulus fiber was used, probably was less than that in vivo, where an otolithic membrane and myriad otoconia are present (15). The natural frequency of spontaneous hair-bundle oscillation increases with elastic loading (P.M. and A.J.H., unpublished observations), which would better accord with the range of saccular responsiveness, 5–130 Hz (16). Second, because of limitations on the duration of recordings from individual hair cells, mechanical stimulation was not always applied precisely at the natural frequency. As expected for an active oscillator operating near an oscillatory instability (17, 18), this deficiency probably limited the range of strikingly nonlinear responsiveness to only one decade of stimulus amplitude, thus reducing the amplification of weak signals.
Responses to Low-Level Stimulation.
Spontaneously oscillatory hair bundles of the bullfrog's sacculus can respond by partial entrainment to sinusoidal displacements of the fiber's base as small as ±1 nm (Fig. 1A). Although this result resembles previous behavioral evidence in various vertebrate species that the auditory threshold corresponds to basilar-membrane vibrations of similar magnitude (19–21), two factors complicate the interpretation of this agreement. First, it remains unclear that an animal's behavioral threshold is in fact associated with a change in the regularity of auditory-nerve activity. Second, because the impedance of the elements in the organ of Corti that mechanically couple the basilar membrane to hair bundles is uncertain, the magnitude of the force experienced by a hair bundle near threshold is unknown.
In all classes of tetrapods, the threshold for phase-locking of spontaneous neural activity may be 10–20 dB lower than that for increased firing of auditory-nerve fibers (22–24). In our experiments, the principal effect of stimuli 1–10 nm in amplitude was an increase in the phase-locking of the response (Fig. 1C); only for still stronger stimulation did the magnitude of hair-bundle movement grow appreciably (Fig. 1B). The correspondence between these observations suggests that hair bundles in general undergo noisy spontaneous oscillations that are entrained increasingly by stimuli of growing size. Acting through a more regular release of synaptic transmitter, this entrainment in turn progressively extends the phase-locking of afferent-nerve activity. When stimuli become strong enough to increase the magnitude of hair-bundle motion, an augmented pace of neurotransmitter release raises the nerve fibers' firing rate as well.
Scaling of Responses with Stimulation.
As the amplitude of stimulation increases from the threshold required to elicit significant phase-locking, a hair bundle's responsiveness declines progressively. Over an order of magnitude, the sensitivity scales as the negative two-thirds power of the stimulus amplitude (Fig. 3D). A power-law relation of this slope is one signature of an active system operating at a Hopf bifurcation, on the verge of an oscillatory instability (17, 18). Although the present results do not further constrain the mechanism of amplification, two active processes have been identified in hair bundles of the bullfrog's sacculus. Myosin-based adaptation of mechanoelectrical transduction can both account for active hair-bundle oscillations at low frequencies and explain the behavior of the bundle's linear response function to small stimuli as a function of frequency (12, 13). Similar systems that involve the concerted movements of clustered myosin molecules are known to be subject to a Hopf bifurcation (25). Ca2+-driven reclosure of transduction channels (14, 26), a mechanism that can operate at relatively high frequencies, has also been shown by modeling to experience a Hopf bifurcation (27).
The scaling of the response to the amplitude of stimulation described in this study applies as well to data collected from the mammalian inner ear. The sensitivity of basilar-membrane movements in the chinchilla's cochlea displays a power-law relation to the acoustical input over 4 orders of stimulus amplitude (figure 4 of ref. 2). Although the point was not emphasized in the original report, a slope of negative two-thirds fits the data well. This result suggests that the active process of the mammalian cochlea also operates near a Hopf bifurcation. Despite the importance of cochlear mechanics in distributing sound energy to appropriately tuned parts of the basilar membrane, the responsiveness of each such segment appears to be dominated by a local mechanical amplifier poised at a Hopf bifurcation (18).
Relation of Hair-Bundle Motility to the Active Process.
Active hair-bundle motility now has been found to display correlates of six features of the inner ear's active process. First, as shown here and elsewhere (11), hair bundles can amplify their mechanical inputs. More precisely, they provide a power gain, an increase in the energy content of a processed signal with respect to the input, that enhances the magnitude of the bundle's response at the frequency of stimulation. Second, also as documented here, amplification by hair bundles is nonlinear. Over much of its range of responsiveness, a bundle's mechanical sensitivity scales as the negative two-thirds power of the input amplitude. The basilar membrane in the mammalian cochlea exhibits the same behavior (ref. 2; reviewed in ref. 3). Third, a hair bundle adopts a passive, linear behavior as the stimulus reaches a high level. A comparable approach to linearity is apparent in basilar-membrane movement (2). Fourth, because of the mechanical nonlinearity of the transduction process (12, 14), hair bundles produce distortion products when exposed to sinusoidal inputs of more than one frequency (28). Similar distortion tones have been documented by human psychophysical experimentation (29, 30), recording of auditory-nerve activity (31), measurement of sound pressure in the ear canal (reviewed in ref. 32), and analysis of basilar-membrane motion (33). Fifth, as documented here and previously (11), mechanical amplification by hair bundles is tuned. Frequency selectivity is one of the principal benefits of the cochlear active process (reviewed in ref. 3). Finally, hair bundles can display spontaneous oscillations (12, 13, 26, 34–36). Unprovoked movements of some constituent of the ear's amplifier are expected to underlie the production of spontaneous otoacoustic emissions (reviewed in refs. 32 and 37).
The six features that characterize the ear's active process are signatures of any dynamical system operating near a Hopf bifurcation (17, 18, 38). That active hair-bundle motility displays those properties therefore does not preclude the possibility that the ear contains oscillators in addition to hair bundles. The similarity of the active process throughout the vertebrates, however, suggests that a common amplificatory mechanism is at work in all mechanically active acousticolateralis organs, the mammalian cochlea among them. It is probable that active hair-bundle motility is essential for amplification and in fact constitutes the active process.
Acknowledgments
We thank Dr. F. Jülicher for assistance with data analysis and Mr. B. Fabella for computer programming. Dr. Jülicher and Drs. J. Prost and M. Magnasco provided stimulating discussions; Drs. G. Manley and D. Kirk and the members of our research group offered valuable comments on the manuscript. This investigation was supported by National Institutes of Health Grant DC00241. During the initial phase of this study, P.M. was an Associate of Howard Hughes Medical Institute, of which A.J.H. remains an Investigator.
Abbreviation
- RMS
root-mean-square
References
- 1.Sivian L J, White S D. J Acoust Soc Am. 1933;4:288–321. [Google Scholar]
- 2.Ruggero M A, Rich N C, Recio A, Narayan S S, Robles L. J Acoust Soc Am. 1997;101:2151–2163. doi: 10.1121/1.418265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ruggero M A. Curr Opin Neurobiol. 1992;2:449–456. doi: 10.1016/0959-4388(92)90179-o. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dallos P. J Neurosci. 1992;12:4575–4585. doi: 10.1523/JNEUROSCI.12-12-04575.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Manley G. Proc Natl Acad Sci USA. 2000;97:11736–11743. doi: 10.1073/pnas.97.22.11736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nobili R, Mammano F, Ashmore J. Trends Neurosci. 1998;21:159–167. doi: 10.1016/s0166-2236(97)01192-2. [DOI] [PubMed] [Google Scholar]
- 7.Manley G A, Kirk D L, Köppl C, Yates G K. Proc Natl Acad Sci USA. 2001;98:2826–2831. doi: 10.1073/pnas.041604998. . (First Published February 13, 2001; 10.1073/pnas.041604998) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hudspeth A J, Choe Y, Mehta A D, Martin P. Proc Natl Acad Sci USA. 2000;97:11765–11772. doi: 10.1073/pnas.97.22.11765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hudspeth A J. Curr Opin Neurobiol. 1997;7:480–486. doi: 10.1016/s0959-4388(97)80026-8. [DOI] [PubMed] [Google Scholar]
- 10.Fettiplace R, Ricci A J, Hackney C M. Trends Neurosci. 2001;24:169–175. doi: 10.1016/s0166-2236(00)01740-9. [DOI] [PubMed] [Google Scholar]
- 11.Martin P, Hudspeth A J. Proc Natl Acad Sci USA. 1999;96:14306–14311. doi: 10.1073/pnas.96.25.14306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Martin P, Mehta A D, Hudspeth A J. Proc Natl Acad Sci USA. 2000;97:12026–12031. doi: 10.1073/pnas.210389497. . (First Published October 10, 2000; 10.1073/pnas.210389497) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Martin P, Hudspeth A J, Jülicher F. Proc Natl Acad Sci USA. 2001;98:14380–14385. doi: 10.1073/pnas.251530598. . (First Published November 27, 2001; 10.1073/pnas.251530598) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Howard J, Hudspeth A J. Neuron. 1988;1:189–199. doi: 10.1016/0896-6273(88)90139-0. [DOI] [PubMed] [Google Scholar]
- 15.Benser M E, Issa N P, Hudspeth A J. Hear Res. 1993;68:243–252. doi: 10.1016/0378-5955(93)90128-n. [DOI] [PubMed] [Google Scholar]
- 16.Yu X, Lewis E R, Feld D. J Comp Physiol A. 1991;169:241–248. doi: 10.1007/BF00215871. [DOI] [PubMed] [Google Scholar]
- 17.Camalet S, Duke T, Jülicher F, Prost J. Proc Natl Acad Sci USA. 2000;97:3183–3188. doi: 10.1073/pnas.97.7.3183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Eguíluz V M, Ospeck M, Choe Y, Hudspeth A J, Magnasco M O. Phys Rev Lett. 2000;84:5232–5235. doi: 10.1103/PhysRevLett.84.5232. [DOI] [PubMed] [Google Scholar]
- 19.Sellick P M, Patuzzi R, Johnstone B M. Hear Res. 1983;10:93–100. doi: 10.1016/0378-5955(83)90019-9. [DOI] [PubMed] [Google Scholar]
- 20.Crawford A C, Fettiplace R. Hear Res. 1983;12:199–208. doi: 10.1016/0378-5955(83)90106-5. [DOI] [PubMed] [Google Scholar]
- 21.Narayan S S, Temchin A N, Recio A, Ruggero M A. Science. 1998;282:1882–1884. doi: 10.1126/science.282.5395.1882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hillery C M, Narins P M. Science. 1984;225:1037–1039. doi: 10.1126/science.6474164. [DOI] [PubMed] [Google Scholar]
- 23.Gleich O, Narins P M. Hear Res. 1988;32:81–91. doi: 10.1016/0378-5955(88)90148-7. [DOI] [PubMed] [Google Scholar]
- 24.Köppl C J. Neuroscience. 1997;17:3312–3321. doi: 10.1523/JNEUROSCI.17-09-03312.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jülicher F, Prost J. Phys Rev Lett. 1997;78:4510–4513. [Google Scholar]
- 26.Benser M E, Marquis R E, Hudspeth A J. J Neurosci. 1996;16:5629–5643. doi: 10.1523/JNEUROSCI.16-18-05629.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Choe Y, Magnasco M O, Hudspeth A J. Proc Natl Acad Sci USA. 1998;95:15321–15326. doi: 10.1073/pnas.95.26.15321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jaramillo F, Markin V S, Hudspeth A J. Nature (London) 1993;364:527–529. doi: 10.1038/364527a0. [DOI] [PubMed] [Google Scholar]
- 29.Goldstein J L. J Acoust Soc Am. 1967;41:676–689. doi: 10.1121/1.1910396. [DOI] [PubMed] [Google Scholar]
- 30.Smoorenburg G F. J Acoust Soc Am. 1972;52:615–632. [Google Scholar]
- 31.Goldstein J L, Kiang N Y S. Proc IEEE. 1968;56:981–992. [Google Scholar]
- 32.Manley G A, Taschenberger G, Kössl M, Fastl H, Janssen T. In: Auditory Worlds: Sensory Analysis and Perception in Animals and Man. Manley G A, Fastl H, Kössl M, Oeckinghaus H, Klump G, editors. Weinheim, Germany: Wiley–VCH; 2000. pp. 93–141. [Google Scholar]
- 33.Robles L, Ruggero M A, Rich N C. Nature (London) 1991;349:413–414. doi: 10.1038/349413a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Crawford A C, Fettiplace R. J Physiol. 1985;364:359–379. doi: 10.1113/jphysiol.1985.sp015750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Howard J, Hudspeth A J. Proc Natl Acad Sci USA. 1987;84:3064–3068. doi: 10.1073/pnas.84.9.3064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Denk W, Webb W W. Hear Res. 1992;60:89–102. doi: 10.1016/0378-5955(92)90062-r. [DOI] [PubMed] [Google Scholar]
- 37.Probst R. Adv Otorhinolaryngol. 1990;44:1–91. doi: 10.1159/000417719. [DOI] [PubMed] [Google Scholar]
- 38.Jülicher F, Andor D, Duke T. Proc Natl Acad Sci USA. 2001;98:9080–9085. doi: 10.1073/pnas.151257898. [DOI] [PMC free article] [PubMed] [Google Scholar]