

Abstract
Phosphate (Pi) is one of the least available plant nutrients found in the soil. A significant amount of phosphate is bound in organic forms in the rhizosphere. Phosphatases produced by plants and microbes are presumed to convert organic phosphorus into available Pi, which is absorbed by plants. In this study we describe the isolation and characterization of a novel tomato (Lycopersicon esculentum) phosphate starvation-induced gene (LePS2) representing an acid phosphatase. LePS2 is a member of a small gene family in tomato. The cDNA is 942 bp long and contains an open reading frame encoding a 269-amino acid polypeptide. The amino acid sequence of LePS2 has a significant similarity with a phosphatase from chicken. Distinct regions of the peptide also share significant identity with the members of HAD and DDDD super families of phosphohydrolases. Many plant homologs of LePS2 are found in the databases. The LePS2 transcripts are induced rapidly in tomato plant and cell culture in the absence of Pi. However, the induction is repressible in the presence of Pi. Divided root studies indicate that internal Pi levels regulate the expression of LePS2. The enhanced expression of LePS2 is a specific response to Pi starvation, and it is not affected by starvation of other nutrients or abiotic stresses. The bacterially (Escherichia coli) expressed protein exhibits phosphatase activity against the synthetic substrate p-nitrophenyl phosphate. The pH optimum of the enzyme activity suggests that LePS2 is an acid phosphatase.
Phosphate (Pi) availability is one of the major growth-limiting factors for plants in many natural ecosystems (Barber, 1980). Plants absorb Pi from the soil as inorganic orthophosphate ions, but their availability is severely restricted by reactions of inorganic and organic phosphates with soil constituents. A significant amount (20%–80%) of the nutrient may be found in organic forms (Richardson, 1994). In response to persistent Pi deficiency plants have developed many adaptive mechanisms to enhance the availability and increase the uptake of Pi. One such adaptive mechanism is production and secretion of phosphatases to release Pi from organic forms (Goldstein et al., 1988a, 1988b; Duff et al., 1989). An increase in the Pi uptake rate of roots and cell cultures following an episode of Pi starvation has also been well documented in plants (Drew and Saker, 1984; Shimogawara and Usada, 1995). A combination of phosphatase activity and enhanced Pi uptake may help plants to acquire required amounts of Pi from the rhizosphere (Plaxton and Carswell, 1999; Raghothama 1999, 2000).
The induction of acid phosphatases (ACPs) is a distinct and universal response of higher plants to Pi starvation (Duff et al., 1994). Due to this unique feature, phosphatase activity has been used as a potential marker of the Pi status of plants (Ascencio, 1994). Changes in specific isoforms of phosphatases under Pi starvation are commonly observed (Goldstein et al., 1988a, 1988b; Trull and Deikman, 1998). Production of extracellular and intracellular phosphatases increases during Pi starvation (Duff et al., 1994). The extracellular phosphatases are likely to be involved in recapturing Pi from organic phosphorus compounds found in the extracellular matrix. The extracellular phosphatases produced in shoots and leaves may also be involved in scavenging Pi from xylem-derived phosphocholine, which is a major phosphorylated compound in the xylem sap (Plaxton and Carswell, 1999). Due to the low substrate specificity, ACPs are presumed to be involved in the non-specific hydrolysis of organic Pi resulting in the restoration of much-needed Pi pool. The control of extracellular, cell wall-associated phosphatase synthesis is a repression-derepression process mediated by Pi status in the tomato (Lycopersicon esculentum) roots (Boütin et al., 1981). A concurrent increase in phosphatases, phytase, and RNases was observed in the absence of exogenous supply of Pi (Bosse and Kock, 1998). Phosphatases are primarily localized, but not limited, to the vacuole and the apoplastic surfaces (Goldstein et al., 1988a; Hubel and Beck, 1996; Li and Todano, 1996; Ascencio, 1997). It is interesting that during Pi starvation, increased activity of phosphatase and phytase is found throughout plant tissues and in the rhizosphere (Hubel and Beck, 1996). Plants such as lupins, which are more efficient in acquiring Pi from the soil, produce significantly more phosphatase compared with cereals (Todano and Komatsu, 1994). In addition, absorption of Pi was enhanced when an extract of ACPs secreted by lupine was added to the rhizosphere of tomato. These findings suggest that production of ACPs is a part of an adaptive strategy by plants to overcome the deficiency of Pi in soils.
Although the role of phosphatases during Pi deficiency is well recognized and significant information is available on enzyme characterization, very few Pi starvation-induced ACP genes have been cloned and characterized. The purple ACP gene isolated from Arabidopsis (GenBank accession no. U48448) and lupine (Wasaki et al., 1999) are few of the known examples of Pi starvation-induced ACPs in plants. In this study we report the isolation and biochemical characterization of a novel tomato Pi starvation-induced gene (LePS2; accession no. AF305968). The similarity of the deduced peptide to other known phosphatases, and enzymatic activity of the bacterially expressed protein, strongly suggests that LePS2 belongs to a new class of Pi-regulated ACP.
RESULTS
LePS2 Is a Novel Pi Starvation-Induced Gene Coding for a Phosphatase
A fragment of LePS2 encoding a novel Pi starvation-induced gene from tomato was isolated by a differential cDNA subtraction strategy. The full-length cDNA clone was obtained from a root library of Pi-starved plants using the gene fragment as a homologous probe. LePS2 is 942 bp long and contains an 810-bp open reading frame sufficient to encode a 269-amino acid polypeptide (30.6 kD). The open reading frame is flanked by 29 and 103 bp of untranslated sequences at the 5′ and 3′ regions, respectively. Hydropathy plots of the deduced polypeptide suggest that LePS2 is a soluble protein (data not shown). The deduced amino acid sequence of the polypeptide shares significant similarity (40%) with that of a phosphatase (AJ006529) identified in the chicken, Gallus gallus (Fig. 1A; Houston et al., 1999). Sequences with high degree of similarity to LePS2 are also found in other plant species (Fig. 1, B and C). Two expressed sequence tags (ESTs) identical to LePS2 were recently reported from the Medicago truncatula sequencing project. An alignment of the amino acid sequence of LePS2 with phosphatases from prokaryotes and eukaryotes revealed the presence of highly conserved motifs (Fig. 1D). The conserved amino acid residues found in the amino terminus are clustered around two distinct motifs, “DFDXT” (motif 1) and “GDGXXD” (motif 2; Fig. 1D). These motifs are conserved in the members of recently identified haloacid dehalogenase and DDDD superfamilies of enzymes catalyzing a diverse number of hydrolytic and phosphotransferase reactions (Aravind et al., 1998; Collet et al., 1998; Houston et al., 1999; Thaller et al., 1998). The members of these superfamilies includes, but is not limited to, phosphatases that catalyze the release of inorganic Pi from a variety of substrates, phosphomutases that transfer Pi groups between adjacent carbon atoms, and the P-type ATPases. The first Asp in the “DFDXT” motif is believed to be transiently phosphorylated during the Pi transfer reaction (Collet et al., 1998). As a consequence, the active sites of these enzymes share similar structural features. The amino acid sequences in LePS2 matches the consensus minimum motif “DFDXT” that is conserved in the phosphatases and phosphomutases (Fig. 1). This motif is slightly divergent from the conserved “DKTGT” motif of the P-type ATPases. Furthermore, the lack of transmembrane domains suggests that LePS2 is not a P-type ATPase. The second conserved “GDGXXD” motif present in LePS2 is generally found in phosphatases rather than in phosphomutases.
Figure 1.

Comparative sequence analysis of LePS2 peptide. A, Alignment of deduced amino acid sequence of LePS2 with that of 3X11A, a phosphatase from chicken. B, Alignment of the N-terminal regions of deduced amino acid sequence of LePS2 with that of an Arabidopsis and rice homolog. Alignment utility used is CLUSTAL X (1.62b). Asterisk, Fully conserved residue; :, a “strong” group is conserved; ., a “weaker” group is conserved. C, Table showing the peptide similarity of LePS2 homologs from plants. Asterisk, Partial EST; double asterisk, bacteria artificial chromosome sequence. D, Alignment of amino acid sequences corresponding to two peptide motifs that are conserved in the active site of the HAD and DDDD superfamily of phosphotransferases. The Asp residue predicted to be transiently phosphorylated is marked by an arrow. Invariant residues are shown in bold and other conserved residues are highlighted.
LePS2 Is a Member of a Small Gene Family in Tomato
The full-length LePS2 cDNA probes hybridized to six bands of the genomic DNA digested with EcoRI and three with XbaI. The Southern hybridization indicates that LePS2 is represented by a small gene family consisting of at least three members in the tomato genome (Fig. 2). Presence of an internal EcoRI site is responsible for six bands seen in EcoRI digested DNA.
Figure 2.
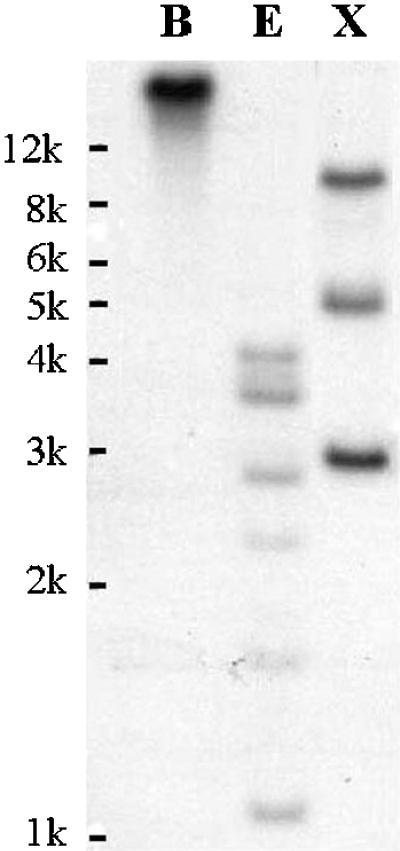
Southern analysis of LePS2. Tomato genomic DNA (10 μg) was digested with BamHI (B), EcoRI (E), and XbaI (X). DNA size markers are indicated.
LePS2 Transcripts Are Induced Specifically by Pi Starvation
The expression of LePS2 was evaluated in tomato plants grown in the presence of 250 μm or no Pi. Comparison was done by northern-blot analysis of total RNA isolated from different tissues. The LePS2 probe hybridized to a 1-kb transcript (Fig. 3, A and B). The expression increased in plants grown under Pi-starved conditions. LePS2 is expressed in all examined parts of the Pi-starved plants (Fig. 3D). An increase in the transcript level was detected within 24 h of Pi starvation in roots and shoots, and it continued to increase with extended duration of Pi starvation reaching a maximum by 5 d. The rapidity of LePS2 induction was quite evident in cell cultures (Fig. 3C). Significant accumulation of the transcripts was observed within 3 h of transferring tomato cell cultures to Pi-deficient media. These results indicate that expression of LePS2 is a rapid response to Pi deficiency in the media.
Figure 3.

Northern-blot analysis of Pi starvation-induced LePS2 gene. Total RNA isolated from roots (A) and leaves (B) of aeroponically grown plants, supplied with one-half strength modified Hoagland solution containing 250 μm Pi (+) or no Pi (−) for the indicated time, were probed with labeled LePS2 cDNA. Expression of LePS2 gene was also analyzed using RNA isolated from cell suspension culture grown in the presence of 250 μm Pi (+) or absence of Pi (−) for indicated time (C). Ethidium bromide-stained gel in the bottom panel indicates RNA integrity and uniform loading. D, Expression of LePS2 in different plant parts under Pi starvation. Tomato plants were grown in the presence of 250 μm Pi (+) or absence of Pi (−) for 7 d. Plants were harvested and dissected into the following parts: S, stem; P, petioles; L, leaf; LL, leaf lamina; and LM, leaf midrib. Total RNA (10 μg/lane) extracted from different parts was used for northern-blot analysis.
The specificity of expression of LePS2 to Pi starvation was analyzed by subjecting the plants to starvation of nitrogen, potassium, and iron as described earlier (Liu et al., 1998). After 5 d a visible retardation in growth and development of deficiency symptoms were noticed in Pi- and nitrogen-starved plants, whereas plants starved of potassium and iron appeared normal. The LePS2 transcript levels increased greatly in Pi-starved plants, but remained at undetectable levels in roots of plants subjected to other nutrient starvation (Fig. 4A). Although we have not tested the effect of starvation of all other essential nutrients, the data suggest a strong correlation between LePS2 expression and Pi deprivation.
Figure 4.

A. LePS2 is induced specifically in response to Pi stress. Plants were provided with one-half strength Hoagland solution deficient in Pi (P−), nitrogen (N–), potassium (K–), or iron (Fe–), and a control solution with all nutrients (P+). Total RNA was isolated and analyzed by northern blots using 32P-labeled LePS2 cDNA. RNA gel stained with ethidium bromide shows uniform loading. B, Abiotic stresses does not affect the expression of LePS2. Plants were grown in hydroponics for 4 d under 4°C or 37°C (root zone temperature), salt (S, 150 mm NaCl), and absence of Pi (P−). A set of plants was allowed to wilt at room temperature (D). Total RNA (10 μg/lane) isolated from these plants was subjected to northern-blot analysis.
Environmental factors such as salt (Pan, 1987) and water deficit (Barret-Lennard et al., 1982) are known to increase the activity of ACPs. The changes in expression of LePS2 in response to abiotic stresses were also examined by northern analysis. Sets of hydroponically grown tomato plants were exposed for 4 d of Pi deficiency, 4°C, 37°C, 150 mm NaCl, and were allowed to wilt at room temperature. By 4 d a visible retardation in growth was observed in 4°C and 37°C stressed plants. The LePS2 transcript levels increased in Pi-starved plants, but remained undetectable in the roots of plants subjected to other stresses (Fig. 4B). These data further support the observation that LePS2 expression is specifically regulated by Pi starvation.
LePS2 Expression Is Dependent on the Availability of Pi in the Medium
To understand the response of LePS2 gene to changing concentration of the nutrient in the media, tomato plants were grown in the presence of different concentrations of Pi. After 5 d of treatment the roots were harvested for isolation of RNA. A noticeable accumulation of LePS2 transcripts was observed in plants provided with 100 μm of Pi and it increased with decreasing concentration of Pi in the media (Fig. 5A). These results suggest a correlation between the amount of phosphorus present in the medium and the level of LePS2 expression. It also points to the existence of a tightly regulated nutrient monitoring system in plants.
Figure 5.

A, Expression of LePS2 in plants supplied with different concentrations of Pi. Plants were supplied with indicated micromolar concentrations of Pi for 5 d. Total RNA (10 μg/lane) isolated from these plants was used for analyzing the expression of LePS2 by northern blots. B, Resupply of Pi suppresses the expression of LePS2. Plants were sprayed with 250 μm Pi (C+) or no Pi (C−) for 5 d. After 5 d a set of plants were continued to grow in the absence of Pi. Another set of Pi-deficient (C−) plants was provided with 250 μm Pi for the indicated time. Bottom, An ethidium bromide-stained gel showing uniform loading of RNA.
The regulation of LePS2 expression by phosphorus availability was further examined by resupplying Pi to the plants that were Pi starved for 5 d and strongly expressing the LePS2. Upon resupply of Pi to these plants, the transcript levels decreased to the levels observed in Pi-sufficient plants within 12 h (Fig. 5B). These observations further support the hypothesis that there is a fine coordination between expression of Pi starvation-induced genes and availability of phosphorus.
LePS2 Expression Is Regulated by Internal Signals
Although it is obvious that LePS2 is responding to changing Pi levels in the media, it is not clear whether changes in the internal or external Pi concentrations act as cues to activate the gene expression. Divided root studies have shown that Pi transporters are induced in response to changes in internal concentration of Pi (Liu et al., 1998). In this study root system of tomato plant was divided into two parts: One-half was exposed to a Pi-deficient solution, and the other half was exposed to a solution with 250 μm Pi. The expression of LePS2 was suppressed in roots and leaves of Pi-sufficient control and divided-root plants (Fig. 6). It is interesting that exposure of one portion of root system to Pi was sufficient to suppress the gene expression in portions of the root that are not provided with Pi. This data suggest that differential uptake of Pi by portions of the root system to meet the Pi demands of plants is sufficient to repress Pi starvation-induced genes such as LePS2. The LePS2 transcript as well as the previously examined Pi starvation-induced genes appear to remain repressed as long as the Pi requirement of plants is adequately met (Liu et al., 1998).
Figure 6.

Internal signals may regulate the expression of LePS2 gene. In this divided-root study, roots of plants were exposed to 250 μm Pi (C+) or no Pi (C−). Roots of another set of plants were separated into two portions and each portion was placed in an aerated hydroponic solution containing 250 μm Pi (D+) or no Pi (D−) for 5 d. Total RNA was isolated and analyzed by northern blot. The L+ and L− represents the RNA isolated from leaves of Pi-sufficient (250 μm) or -deficient plants. Ethidium bromide-stained gel indicates RNA integrity and uniform loading.
Bacterially Expressed LePS2 Protein Has Phosphatase Activity
The LePS2 coding sequence was expressed in Escherichia coli. This system allowed the purification of the LePS2 protein via His tag and metal affinity column (Fig. 7A). Bacteria expressed LePS2 to a high level after induction with isopropylthio-β-galactoside (IPTG), and the protein was found in the soluble fraction and the inclusion bodies. The bacterially expressed protein was resolved as a 30-kD peptide on SDS-PAGE. The presence of His tags facilitated purification of the peptide by His binding-Ni affinity column. Different fractions collected from the column were analyzed for phosphatase activity against the synthetic substrate p-nitrophenyl phosphate (Fig. 7B). The eluted protein showed low, but significant phosphatase activity as compared with similar eluates from bacteria expressing vector sequences alone. The phosphatase activity has a pH optimum of 4 to 5, suggesting that LePS2 is an ACP (Fig. 7C).
Figure 7.

A, Expression of LePS2 in E. coli. The coding sequence of LePS2 gene was cloned into a pET28a vector and transferred to BL21, pLysis S strain of bacteria. The cells were induced to produce recombinant protein by IPTG. Bacterial proteins were separated on a 10% (w/v) SDS-PAGE. M, Marker; BL21, control; 0i, induced BL21 carrying pET28a vector; Lei, induced BL21 carrying LePS2 coding sequence in a pET28a vector; LeP, Ni affinity column purified recombinant LePS2 protein. Arrow indicates 30-kD LePS2 protein. B, Bacterially expressed LePS2 has ACP activity. The synthetic substrate p-nitrophenol Pi was used to quantify the activity of the column-purified LePS2 protein. The assay was done with extracts from bacteria containing only vector (pET-0) as a control or recombinant protein (LePS2). The fractions 1 to 5 were eluted from the Ni affinity column and tested for phosphatase activity. Data suggest that LePS2 is an ACP with a pH optimum of 4 to 5 C, Error bars represent the sd of all samples tested.
DISCUSSION
The two major factors affecting Pi acquisition are availability of Pi in the rhizosphere and ability of plants to absorb available Pi. These factors need to be coordinated for successful completion of the plant life cycle. It is well documented that plants produce and secrete ACPs into the rhizosphere to release Pi from organic Pi complexes. Production of phosphatases is linked to Pi deficiency and a positive correlation between ACP production and Pi nutrition has been proposed (Jungk et al., 1993; Ascenio, 1994). Many ACPs are known to release Pi from internal and extracellular pools of organic Pi, and as a consequence they are of considerable importance to plants growing under the nutrient-limiting conditions. These enzymes can significantly impact the availability and recycling of Pi in and around the plant. LePS2 is likely to represent one such Pi-regulated ACP that could play a role in enhancing the availability and utilization of organic Pi in plants.
The amino acid sequence of LePS2 has significant similarity with that of a recently identified phosphatase from chicken (Houston et al., 1999). The hydrophilic nature of the deduced protein indicates that LePS2 is likely a soluble enzyme. The amino terminus of the protein contains stretches of conserved amino acids found in the active site of members of the HAD and DDDD super families of phosphotransferases. There is no discernable signal peptide at the carboxy or the amino terminus in the deduced protein suggestive of its targeting to chloroplast, vacuole, or other cellular organs. A lack of clear targeting sequences may point to the extracellular localization of the LePS2 by the default pathway (Denecke et al., 1990). Homologs of this gene are also found in a variety of organisms, including vertebrates and plants. It is interesting to note that ESTs similar to LePS2 are found in Pi-starved root library of M. truncatula. The presence of LePS2 homologs in diverse plant species demonstrates that the structure, and possibly the function, of this gene product are conserved across the plant kingdom.
LePS2 is rapidly induced in response to Pi stress in cell cultures and plants. The rapidity of induction, within 3 and 12 h of transfer of cell cultures and plants, respectively, to Pi-deficient media indicate that LePS2 is among the genes induced early in the Pi starvation rescue process. This observation concurs with the data showing that ACP activity was induced within 24 h of transferring tomato cells to Pi-deficient medium (Goldstein et al., 1988b). A 6-fold difference in the amount of excreted phosphatase between Pi-starved and Pi-sufficient cells was observed in these studies. Furthermore, increases in phosphatase activity within hours of Pi starvation in Brassica nigra cells support the rapid induction of phosphatases during Pi deficiency (Plaxton and Carswell, 1999). The pattern of LePS2 expression is similar to that of Pi transporters (Liu et al., 1998) and TPSI1, a novel Pi starvation-induced gene in tomato (Liu et al., 1996). Even though the spatial distribution of LePS2 phosphatase is not thoroughly analyzed, it is becoming clear that expression of Pi starvation-induced genes such as LePS2 and Pi transporters are spatially and temporally regulated. The coordinated expression of the Pi starvation rescue genes is crucial for the survival of a plant in the nature, where it has to compete with other microorganisms for the same pool of Pi in the rhizosphere. Existence of similar mechanisms in microorganisms indicate that Pi starvation rescue mechanisms are conserved throughout evolution (Raghothama, 1999).
The expression of LePS2 is strongly influenced by altered Pi levels in the media. A strong induction of the LePS2 gene was observed when the Pi concentration was reduced to 10 μm. The gene induction in response to decreasing concentration of Pi is similar to the rapid Pi influx response that followed a decline in endogenous Pi in cell cultures (Lefebvre et al., 1990). Furthermore, a correlation between the amount of ACP excreted and the degree of Pi starvation has been established in tomato suspension cultures (Goldstein et al., 1988a). The concentration of Pi at which LePS2 is induced is much higher than the naturally occurring Pi concentrations in the soil (Barber, 1980). This data suggests that Pi starvation-induced genes are activated well in advance of impending biochemical and physiological changes in plants. The induction and repression of LePS2 is a rapid and specific response to altered Pi levels in the media. Transfer of Pi-deficient plants actively expressing LePS2 to Pi-sufficient media repressed the expression of the gene. This response mimics the induction and repression of secreted ACPs by Pi in tomato and tobacco (Ueki and Sato, 1971; Goldstein et al., 1988a).
It is becoming clear that genes involved in Pi starvation-induced rescue mechanisms are regulated by altered Pi concentration in plants. These include phosphatases, Pi transporters, and a novel Pi starvation-induced gene TPSI1, which responded to changes in internal Pi status of tomato (Liu et al., 1998; Baldwin et al., 1999). A detailed analysis of ACP activity and Pi uptake in B. nigra suspension cultures showed that both processes are affected by the Pi status of the cells (Lefebvre et al., 1990). The LePS2 expression concur with earlier observations that once Pi needs of plants are met the ability of plant to absorb the nutrient decreases (Lefebvre and Glass, 1982; Drew and Saker, 1984). The data presented here, taken together with earlier published reports, show that many Pi starvation-induced responses are repressed and derepressed by changing levels of Pi in plant tissues. This pattern of regulation should allow plants to maintain cellular Pi homeostasis without causing a toxic accumulation of Pi or wasteful synthesis of enzymes and proteins involved in the nutrient utilization processes. ACPs induced by Pi deficiency are often under tight local expression control by plants. The control is so precise that in some instances only a small portion of the root may be expressing ACP responding to the distribution of organic P (Boütin et al., 1981). Thus far, the northern analysis showed that LePS2 is expressed in all the plant parts tested. The expression of LePS2 in all parts of plants is suggestive of its global role in enhancing Pi availability and possible recycling of organic Pi compounds.
The biochemical evidence supports the sequence homology prediction that LePS2 is a phosphatase. The LePS2 expressed in E. coli showed small, but significant ACP activity. The low activity of the bacterially expressed protein could be due to several reasons including improper folding of the peptide and lack of post-translational modifications such as glycosylation and phosphorylation. Many excreted proteins such as ACP often contain a disulfide bond as part of its native structure and they generally function as dimers. The observation that majority of the ACP exists as glycoproteins (Duff et al., 1994) suggests that bacterially expressed protein may lack kinetic properties of the native protein. Furthermore, the phosphatase activity of LePS2 was measured with the synthetic substrate p-nitrophenyl phosphate. Although the ACPs secreted by plants under Pi starvation are known to lack substrate specificity in general (Ozawa et al., 1995), there are precedents for the existence of ACP with known substrate specificities (Duff et al., 1994; Bose and Taneja, 1998). In such a situation p-nitrophenyl phosphate may not be the ideal substrate for hydrolysis by LePS2 peptide. Identifying the native substrate(s) will be a challenging part of the future research. The bacterially expressed protein resolves as a 30-kD peptide on the SDS gel. The phosphatases induced under Pi starvation in tomato were resolved into high and low Mr components (Goldstein et al., 1988b). The low Mr component contained a 57-kD protein exhibiting strong phosphatase activity. It is tempting to speculate that a dimer or a glycosylated form of LePS2 may be representing a part of this activity. This speculation is based on the observation that ACPs secreted by tomato and lupin are homodimers (Ozawa et al., 1995; Li and Tadano, 1996). However, LePS2 may well represent a novel phosphatase in plants. These observations need further confirmation by immunolocalization of the peptide and in vivo expression studies in plants.
The data presented here, in combination with earlier reports on the regulation of Pi transporters (Muchhal et al., 1996; Muchhal and Raghothama, 1999) and Pi generating enzymes (Plaxton and Carswell, 1999) by phosphorus, point to the existence of a highly coordinated Pi starvation response mechanism in plants. A synchronized activation of genes such as phosphatases, Pi transporters, Rnases, etc. in combination with rapid root and root hair proliferation, and internal adjustments in Pi homeostasis will help plants to survive under Pi starvation.
MATERIALS AND METHODS
Plant Materials
Tomato (Lycopersicon esculentum) plants were grown in an aeroponic growth facility similar to the one described by Liu et al. (1998). Tomato seeds of the variety OS4 were germinated in seedling trays filled with Ready Earth plug mixture (Scotts, Marysville, OH). When plants reached the four-leaf stage (20 d after sowing) they were removed from the growing medium and the roots were washed free of medium and transferred to aeroponics. In aeroponic culture the roots were sprayed with a fine mist of one-half-strength Hoagland solution (Jones, 1982) for 3 s every 10 min. Phosphorus starvation treatments were initiated 1 week after the transfer to aeroponics. For divided root system studies tomato plants were grown in aeroponics for 1 week as described above. Three aeroponic plants were transferred to an aerated hydroponics solution containing either 250 μm or no Pi. The roots of three other plants were separated into two sections, and each section of the roots was placed in adjacent containers with aerated nutrient solution containing either 250 μm or no Pi. Five days after transfer leaves and roots from P+ (250 μm Pi) and P− (0 μm Pi) and divided root-system plants were harvested, frozen in liquid nitrogen, and stored at −70°C.
For abiotic stress studies, hydroponically grown tomato plants were transferred to 4°C and 37°C (root zone temperature) chambers for 4 d. The salt treatment was initiated by adding 150 mm NaCl to one-half strength Hoagland nutrient solution. Several plants were allowed to wilt at room temperature over a period of at least 3 h as a desiccation treatment. The deficiency of other nutrients was imposed by removing the nutrients (N, K, and Fe) in the solution for 5 d.
Tomato cell cultures (cv VFNT Cherry) were maintained in the laboratory as described earlier (Bressan et al., 1981). Phosphorus deficiency treatments were initiated 7 d after subculturing the cells. A 25-mL portion of cell suspension, grown in Pi-sufficient media, was filtered through Miracloth (Calbiochem, CA) and was washed with Pi-deficient (0 μm) or -sufficient (250 μm) media. The cells were then transferred to the same media used for washing. Samples were collected at indicated times after transferring the cells to Pi-deficient medium, centrifuged at 2,000 rpm for 4 min, and the resulting pellet was used for RNA isolation.
Construction of Subtraction cDNA Library and Isolation of LePS2
The subtraction library was prepared using PCR-Select cDNA subtraction kit according to the manufacturer's (Clonetech, Palo Alto, CA) instructions. The poly(A)+ RNA isolated from roots of tomato plants subjected to phosphorus deficiency for 7 d (P−) or provided with 250 μm Pi (P+) were used for cDNA preparation. The DNA fragments showing significant differences in hybridization with labeled cDNAs prepared with RNA isolated from P− and P+ plant roots were cloned into the pCR script 2.1 vector via TA overhangs. The ligation products were used to transform Escherichia coli (XL1 Blue) cells. The amplified cells represent subtraction libraries enriched with differentially expressed gene fragments.
Differential screening was carried out by slot-blot hybridization (S&S, Keene, NH). Plasmid DNA isolated independently from 600 colonies was denatured and equal quantities of DNA were loaded into replicate slot blots. Sets of replicated blots were probed with 32P cDNA prepared with poly(A)+ RNA representing P+ and P− roots. Hybridizations were carried out in a solution containing 50% (v/v) formamide at 42°C. Final washing of the filters was done with 1× SSC and 0.2% (w/v) SDS at 55°C for 30 min. After autoradiography the blots were striped of the radioactivity and the blots probed with P+ cDNAs were re-probed with P− cDNAs and vice versa. The re-probing of blots was essential to reduce non-specific background signals. The positive clones were then subjected to a second round of screening followed by northern analysis. One of the cDNA clones representing a differentially expressed gene in tomato was named LePS2.
Isolation of Full-Length LePS2 cDNA Clone
A cDNA library prepared with the mRNA isolated from Pi-starved tomato roots (Liu et al., 1996) was screened with 32P-labeled LePS2 according to standard procedures (Sambrook et al., 1989). The hybridization and washing conditions were similar to those described above. Based on the insert size and restriction mapping, one full-length representative was used for further analysis. Both the strands were sequenced at the Iowa State University DNA sequencing facility.
Northern-Blot Analysis of the Transcripts
Total RNA was isolated from roots and leaves of tomato plants by hot-phenol extraction and lithium chloride precipitation (Pawlowski et al., 1994). Ten micrograms of total RNA was electrophoretically separated on 1% (w/v) denaturing formaldehyde agarose gels and blotted onto a nitrocellulose membrane (BA-S, SS, Keene, NH; Sambrook et al., 1989). The nitrocellulose membranes were hybridized overnight with a 32P-labeled probe (106 cpm/mL) in a solution containing 50% (w/v) formamide, 5× Denhart's solution, 0.1% (w/v) SDS, 6× SSPE, and 100 μg/mL denatured salmon-sperm DNA at 42°C. Filters were washed twice in 2× SSC and 0.2% (w/v) SDS at room temperature for 10 min, twice in 1× SSC and 0.2% (w/v) SDS at 55°C for 15 min, and twice in 0.1× SSC and 0.2% (w/v) SDS at 55°C for 20 min before autoradiography.
Southern Blots
High Mr genomic DNA was isolated from young leaves of tomato as described by Dellaporta et al. (1983). Ten micrograms of genomic DNA was digested with restriction enzymes, electrophoresed on a 0.8% (w/v) agarose gels, denatured, and blotted onto a supported nitrocellulose membrane (Sambrook et al., 1989). The hybridization and washing conditions were the same as those used for northern blots.
Expression of LePS2 in E. coli and Purification
The coding sequence of LePS2 was cloned into the pET 28a expression vector and transferred to E. coli (BL21 P-lysis S) cells. The recombinant clone has His tags on the N and C terminus of LePS2 peptide. To express LePS2, cells were grown at 37°C to an optical density of 0.6, and IPTG was added to a final concentration of 1 mm. Cells were grown for 2 h in the presence of IPTG, then washed twice with binding buffer, and resuspended in 15 mL of binding buffer (Novagen, Madison, WI). The cells were sonicated to lysis and processed through Ni affinity column as per the manufacturer's recommendations (Novagen). During elution a series of 1-mL fractions were collected for protein estimation and phosphatase assay. The purity of the affinity column purified LePS2 protein was checked on a SDS-PAGE.
The LePS2 phosphatase activity was measured at 37°C and pH 4.0 to 5.5 using p-nitrophenyl Pi as the substrate as previously described (Sigma Diagnostics, 1985). The reaction was terminated using 0.1 m NaOH and OD was measured at 410 nm in a spectrophotometer.
Computer Analysis
The BLAST programs from NCBI were used to search for homologous genes in the database (Altschul et al., 1990). Multiple sequence alignments were performed with CLUSTAL X (Thompson et al., 1997).
ACKNOWLEDGMENT
We thank Dr. Ronald C. Coolbaugh for critically reviewing the manuscript.
Footnotes
This research is funded in part by the U.S. Department of Agriculture-National Research Initiative Competitive Grants Program (grant no. 97–35100–4211). This is journal paper no. 16,382 of the Purdue University Agricultural Research Programs.
LITERATURE CITED
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- Aravind L, Galperin MY, Koonin EV. The catalytic domain of the P-type ATPase has the haloacid dehalogenase fold. Trends Biochem Sci. 1998;23:127–129. doi: 10.1016/s0968-0004(98)01189-x. [DOI] [PubMed] [Google Scholar]
- Ascencio J. Acid phosphatase as a diagnostic tool. Commun Soil Sci Plant Anal. 1994;25:1553–1564. [Google Scholar]
- Baldwin JC, Karthikeyan AS, Raghothama KG. LePS2, a phosphate starvation induced acid phosphatase gene from tomato. Plant Physiol. 1999;99:S-190. doi: 10.1104/pp.125.2.728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barber SA. Soil-plant interactions in the phosphorus nutrition of plants. In: Khasawneh FE, Sample EC, Kamprath EJ, editors. Role of Phosphorus in Agriculture. Madison, WI: American Society of Agronomy; 1980. pp. 591–615. [Google Scholar]
- Barret-Lennard EG, Robson AD, Greenway H. Effect of phosphorous deficiency and water deficit on phosphatase activities from wheat leaves. J Exp Bot. 1982;33:682–693. [Google Scholar]
- Bose SK, Taneja V. Induction of a germination specific, low molecular weight, acid phosphatase isozyme with specific phosphotyrosine phosphatase activity in lentil (Lens esculenta) seeds. Biochem Biophys Res Commun. 1998;250:629–634. doi: 10.1006/bbrc.1998.9364. [DOI] [PubMed] [Google Scholar]
- Bosse D, Kock M. Influence of phosphate starvation on phosphohydrolases during development of tomato seedlings. Plant Cell Environ. 1998;21:325–32. [Google Scholar]
- Boütin J, Provot M, Roux L. Effect of cycloheximide and renewal of phosphorus supply on surface acid phosphatase activity of phosphorus deficient tomato roots. Physiol Plant. 1981;51:353–360. [Google Scholar]
- Bressan RA, Hasegawa PM, Handa AK. Resistance of cultured higher plant cells to polyethylene glycol-induced water stress. Plant Sci Lett. 1981;21:23–30. [Google Scholar]
- Collet JF, Stroobant V, Pirard M, Delpierre G, Van Schaftingen E. A new class of phosphotransferases phosphorylated on an aspartate residue in an amino-terminal DXDX(T/V) motif. J Biol Chem. 1998;273:14107–14112. doi: 10.1074/jbc.273.23.14107. [DOI] [PubMed] [Google Scholar]
- Dellaporta SL, Wood J, Hicks JB. A plant DNA minipreparation: version II. Plant Mol Biol Rep. 1983;1:19–21. [Google Scholar]
- Denecke J, Botterman J, Deblaere R. Protein secretion in plant cells can occur via a default pathway. Plant Cell. 1990;2:51–59. doi: 10.1105/tpc.2.1.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drew MC, Saker LR. Uptake and long-distance transport of phosphate, potassium and chloride in relation to internal ion concentrations in barley: evidence of non-allosteric regulation. Planta. 1984;160:500–507. doi: 10.1007/BF00411137. [DOI] [PubMed] [Google Scholar]
- Duff SMG, Moorhead GBG, Lefebvre DD, Plaxton WC. Phosphate starvation inducible “Bypasses” of adenylate and phosphate dependent glycolytic enzymes in Brassica nigra suspension cells. Plant Physiol. 1989;90:1275–1278. doi: 10.1104/pp.90.4.1275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duff SMG, Sarath G, Plaxton WC. The role of acid phosphatase in plant phosphorus metabolism. Physiol Plant. 1994;90:791–800. [Google Scholar]
- Goldstein AH, Baertlein DA, Danon A, Tibbot BK. Phosphate inducible metabolism in Lycopersicon esculentum: excretion of acid phosphatase by tomato plants and suspension-cultured cells II. Plant Physiol. 1988a;87:711–715. doi: 10.1104/pp.87.3.711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein AH, Danon A, Baertlein DA, McDaniel RG. Phosphate inducible metabolism in Lycopersicon esculentum: characterization of acid phosphatase by tomato and suspension-cultured cells I. Plant Physiol. 1988b;87:716–720. doi: 10.1104/pp.87.3.716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houston B, Seawright E, Jefferies D, Hoogland E, Lester D, Whitehead C, Farquharson C. Identification and cloning of a novel phosphatase expressed at high levels in differentiating growth plate condrocytes. Biochem Biophys Acta. 1999;1448:500–506. doi: 10.1016/s0167-4889(98)00153-0. [DOI] [PubMed] [Google Scholar]
- Hubel F, Beck E. Maize root phytase: purification, characterization, and localization of enzyme activity and its putative substrate. Plant Physiol. 1996;112:1429–1436. doi: 10.1104/pp.112.4.1429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones JB. Hydroponics: its history and use in plant nutrition studies. J Plant Physiol. 1982;5:1005–1030. [Google Scholar]
- Jungk A, Seeeling B, Gerke J. Mobilization of different phosphate fractions in the rhizosphere. Plant Soil. 1993;155/156:91–94. [Google Scholar]
- Lefebvre DD, Duff SMG, Fife CA, Julien-lnalsingh C, Plaxton WC. Response to phosphate deprivation in Brassica nigra suspension. Physiol Plant. 1990;93:504–511. doi: 10.1104/pp.93.2.504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lefebvre DD, Glass ADM. Regulation of phosphate influx in barley roots: effects of phosphate deprivation and reduction of influx with provision of orthophosphate. Physiol Plant. 1982;54:199–206. [Google Scholar]
- Li M, Todano T. Comparison of characteristics of acid phosphatases secreted from roots of lupin and tomato. Soil Sci Plant Nutr. 1996;42:753–763. [Google Scholar]
- Liu C, Muchhal US, Mukatira U, Kononowicz AK, Raghothama KG. Tomato phosphate transporter genes are differentially regulated in plant tissues by phosphorus. Plant Physiol. 1998;116:91–99. doi: 10.1104/pp.116.1.91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu C, Muchhal US, Raghothama KG. Differential expression of TPSI1, a phosphate starvation-induced gene in tomato. Plant Mol Biol. 1996;33:867–874. doi: 10.1023/a:1005729309569. [DOI] [PubMed] [Google Scholar]
- Muchhal US, Pardo JM, Raghothama KG. Phosphate transporters from the higher plant Arabidopsis thaliana. Proc Natl Acad Sci USA. 1996;93:10519–10523. doi: 10.1073/pnas.93.19.10519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muchhal US, Raghothama KG. Transcriptional regulation of plant phosphate transporters. Proc Natl Acad Sci USA. 1999;96:5868–5872. doi: 10.1073/pnas.96.10.5868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozawa K, Osaki M, Matsui H, Honma M, Tando T. Purification and properties of acid phosphatase secreted from lupin root under phosphorus-deficient conditions. Soil Sci Plant Nutr. 1995;41:461–469. [Google Scholar]
- Pan S. Characteristics of multiple acid phosphatases in salt stressed spinach leaves. Aust J Plant Physiol. 1987;14:117–124. [Google Scholar]
- Pawlowski K, Kunze R, De Vries S, Bisseling T. Isolation of total, poly(A) and polysomal RNA from plant tissues. In: Gelvin SB, Shilperoort RA, editors. Plant Molecular Biology Manual. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1994. pp. 1–13. [Google Scholar]
- Plaxton WC, Carswell MC. Metabolic aspects of the phosphate starvation response in plants. In: Lerner HR, editor. Plant Responses to Environmental Stresses: From Phytohormones to Genome Reorganization. New York: Marcel Dekker; 1999. pp. 349–372. [Google Scholar]
- Raghothama KG. Phosphate acquisition. Annu Rev Plant Physiol Mol Biol. 1999;50:665–693. doi: 10.1146/annurev.arplant.50.1.665. [DOI] [PubMed] [Google Scholar]
- Raghothama KG. Phosphate transport and signaling. Curr Opin Plant Biol. 2000;3:182–187. [PubMed] [Google Scholar]
- Richardson AE. Soil microorganisms and phosphorus availability. In: Pankhurst CE, Doube BM, Gupta VVSR, Grace PR, editors. Soil Biota Management in Sustainable Farming Systems. Australia: CSIRO; 1994. pp. 50–62. [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Shimogawara K, Usuda H. Uptake of inorganic phosphate by suspension-cultured tobacco cells: kinetics and regulation by Pi starvation. Plant Cell Physiol. 1995;36:341–351. [Google Scholar]
- Sigma Diagnostics. Phosphatase, alkaline, acid, prostatic: procedure no. 104. Sigma Chemical Company; 1985. [Google Scholar]
- Thaller MC, Schippa S, Rossolini GM. Conserved sequence motifs among bacterial, eukaryotic and archaeal phosphatases that define a new phosphohydrolase superfamily. Protein Sci. 1998;7:1651–1656. doi: 10.1002/pro.5560070722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997;24:4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todano T, Komatsu K. Utilization of organic phosphorus in the soil by plant roots. Trans 15th World Congress of Soil Sci. 1994;9:521–522. [Google Scholar]
- Trull MC, Deikman J. An Arabidopsis mutant missing one acid phosphatase isoform. Planta. 1998;206:544–550. doi: 10.1007/s004250050431. [DOI] [PubMed] [Google Scholar]
- Ueki K, Sato S. Effect of inorganic phosphate on the extracellular acid phosphatase activity of tobacco cells cultured in vitro. Physiol Plant. 1971;24:506–511. [Google Scholar]
- Wasaki J, Omura M, Osaki M, Ito H, Matsui H, Shinano T, Tadano T. Structure of a cDNA for an acid phosphatase from phosphate-deficient lupin (Lupinus albus L.) roots. Soil Sci Plant Nutr. 1999;45:439–449. [Google Scholar]