

Abstract
Diatoms contribute nearly half of the marine primary production. These microalgae differ from other phytoplankton groups in having a silicified cell wall, which is the strongest known biological material relative to its density. While it has been suggested that a siliceous wall may have evolved as a mechanical protection against grazing, empirical evidence of its defensive role is limited. Here, we experimentally demonstrate that grazing by adult copepods and nauplii on diatoms is approximately inversely proportional to their silica content, both within and among diatom species. While a sixfold increase in silica content leads to a fourfold decrease in copepod grazing, silicification provides no protection against protozoan grazers that directly engulf their prey. We also found that the wall provides limited protection to cells ingested by copepods, since less than 1% of consumed cells were alive in the faecal pellets. Moreover, silica deposition in diatoms decreases with increasing growth rates, suggesting a possible cost of defence. Overall, our results demonstrate that thickening of silica walls is an effective defence strategy against copepods. This suggests that the plasticity of silicification in diatoms may have evolved as a response to copepod grazing pressure, whose specialized tools to break silicified walls have coevolved with diatoms.
Keywords: silica wall, diatoms, traits and trade-offs, mechanical defence, zooplankton grazing
1. Introduction
Higher plants have a wide range of morphological and chemical defensive traits to avoid or reduce consumption by herbivores [1]. Similarly, unicellular organisms of the pelagic realm have evolved a stunning array of defences, including escape behaviours, toxin production and morphological traits, such as colony formation and strengthened cell walls [2]. In many cases, these traits are inducible, meaning that they are activated or intensified in response to the presence of grazers or grazer cues, which strongly supports their defensive role [3–6]. However, often the benefits to the defended organism remain somewhat speculative, and quantitative estimates of the trade-offs are rare [2]. This also applies to the role of silicified cell walls in diatoms, whose protective function together with a number of other adaptive roles have been suggested. For example, the siliceous wall may provide necessary support for the large vacuole [7–9], enhance nutrient uptake [10], facilitate light harvesting [11] and/or protect the cell against UV radiation [12]. Hence, the silica wall may not necessarily only, or mainly, represent an adaptation to reduce herbivory, as is the case in higher plants [13], and the scarcity of experimental evidence for its defensive role in diatoms makes ample room for speculations.
The grazers of diatoms are diverse and include both metazoans (e.g. copepods and euphausiids) and protozoans (e.g. ciliates and heterotrophic dinoflagellates) [7,14,15]. Copepods are key grazers in the ocean since they account for approximately 80% of the mesozooplankton biomass [16]. Calanoid copepods bear powerful mandibles lined with silica-reinforced teeth, well suited to crack diatom shells [17]. Although diatoms that are smaller than the copepod mouth may be directly ingested and survive the passage through the copepod guts [18–20], it has been suggested that the copepods taste and break diatoms before ingestion [21]. However, in some cases, the siliceous wall is able to withstand the crushing forces exerted by the copepod mandibles [21–23]. Planktonic protozoans, such as heterotrophic dinoflagellates and large ciliates, are another major group of grazers in the ocean [14]. Although the silica content in diatoms can affect the palatability and digestibility to protozoan grazers [24], little is known about the potential defensive role of silicified walls against these grazers.
While all diatoms have silicified cell walls, there is a large variation in silica content among and even within species [25–30]. This variation in the silica content may imply different levels of mechanical strength and consequently susceptibility to copepod grazing. Although some studies have suggested that diatoms with a weakly silicified frustule experience higher grazing mortality [23,30], the quantitative relationship between silica content and predation mortality remains unknown. On the other hand, the silica shell is probably of little consequence to heterotrophic dinoflagellates and large ciliates [14]—thus far, very little is known about the potential defensive role of silicified walls against protozoan grazers.
According to optimal defence theory, the advantage of a defence mechanism must come at some cost. Since diatoms have evolved an obligate requirement for dissolved silica, low concentrations of silicon in the environment will limit their growth, which can be regarded as a potential cost of silicification [9,31]. Furthermore, silicon deposition depends on the growth rate of diatoms. Many diatom species do not store sufficient amounts of silicon for the formation of a new valve and must, therefore, harvest most of the required quantities immediately before the cell division [25]. Consequently, fast growth will lead to a reduced period available for silicon uptake, which will result in lighter silicification of the walls, even under non-limiting silicon conditions [32]. Therefore, if increased silicification in response to grazer cues requires reduced growth, this may represent a direct cost of defence. Other costs appear to be less important, and include biochemical costs related to the uptake and deposition of dissolved silica [33], and the costs associated with the potential losses due to elevated sinking rates of heavy silicified cells [2,34].
Here, we experimentally investigated how different degrees of silicification within and among diatom species affect predation by different types of planktonic grazer: adult copepods, copepod nauplii and heterotrophic dinoflagellates that engulf their prey. Moreover, we examined whether the survival of diatoms during the gut passage of copepods is related to the silica content of the cells. Finally, we assessed the potential cost of diatom silicification in terms of growth reduction.
2. Material and methods
(a). Organisms and culture conditions
The diatom species used as prey were Amphiprora paludosa (CCMP125), Cyclotella cryptica (CCMP331), Cyclotella meneghiniana (CCMP336), Navicula incerta (CCMP542), Nitzschia laevis (CCMP559), Thalassiosira guillardii (CCMP988) and Thalassiosira weissflogii (unknown strain, UNK). The first six species were obtained from the NCMA at Bigelow Laboratory, Maine, while T. weissflogii was obtained from DTU Aqua, Denmark. The species are all non-chain forming, have no spines or other cell structures, are non-toxic and of similar size to minimize other effects than silica content on grazing (table 1). We used different levels of light to acquire different degrees of silicification within a species, as silica content in cells increases under conditions of light limitation [35]. Specifically, each of the seven monocultures was split into two subcultures that were placed under low (15 µmol photons m−2 s−1) or high (120 µmol photons m−2 s−1) light conditions (14 : 10 L : D cycle). All subcultures were maintained at a temperature of 15°C in B1 medium with silica [36], and grown until the stationary phase, where the cells contained maximum concentrations of biogenic silica.
Table 1.
Functional group of phytoplankton, prey species, abbreviated species name, cell size expressed as equivalent spherical diameter (ESD) and longest dimension, and type of grazer (adult copepods, nauplii NIII-IV and dinoflagellates). ESD was measured with a Multisizer 3 Coulter Counter and the longest dimension of pennate diatoms with an optical microscope (Leica, magnification 200×; measured number of cells > 130).
| prey group | prey species | abbreviated name | ESD (μm) | longest dimension (μm) | grazer |
|---|---|---|---|---|---|
| pennate diatom | Amphiprora paludosa | AP | 8 | 15 | copepod, dinoflagellate |
| centric diatom | Cyclotella cryptica | CC | 9 | 9 | copepod, nauplii, dinoflagellate |
| centric diatom | Cyclotella meneghiniana | CM | 8 | 8 | copepod, dinoflagellate |
| pennate diatom | Navicula incerta | NI | 7 | 13 | copepod, dinoflagellate |
| pennate diatom | Nitzschia laevis | NL | 6 | 14 | copepod |
| centric diatom | Thalassiosira guillardii | TG | 6 | 6 | copepod |
| centric diatom | Thalassiosira weissflogii | TW | 13 | 13 | copepod, nauplii |
| nanoflagellate | Heterosigma akashiwo | NFL | 11 | 11 | dinoflagellate |
| nanoflagellate | Isochrysis galbana | NFL | 4 | 4 | dinoflagellate |
| nanoflagellate | Rhodomonas salina | NFL | 6 | 6 | nauplii, dinoflagellate |
| nanoflagellate | Tetraselmis sp. | NFL | 8 | 8 | dinoflagellate |
We also measured grazing on various species of nanoflagellates (non-silicified prey) in the experiments with copepod nauplii and heterotrophic dinoflagellates to separate size- and silica effects on grazing rates. The nanoflagellate species used as prey were Heterosigma akashiwo (UNK), Isochrysis galbana (UNK), Rhodomonas salina (UNK) and Tetraselmis sp. (table 1). All species were obtained from DTU Aqua (Denmark) and maintained in cultures in B1 medium.
Grazers were adults and nauplii of the copepod Temora longicornis (approx. 1 mm cephalothorax length and 0.3 mm body length, respectively) isolated from the Danish coastal waters and maintained in continuous laboratory cultures at 15–18°C and a salinity of 32 psu in the dark, on a mixed diet of phytoflagellates and diatoms for several generations. To obtain nauplii cohorts of T. longicornis, adults were separated from the stock culture with a 200-µm-mesh sieve and placed in a new tank. After 48 h, the adults were removed and newly produced eggs were transferred to a new tank with food ad libitum for 4 days until nauplii stages NIII-IV were obtained. The dinoflagellate Oxyrrhis marina (ESD 17 µm) was obtained from DTU Aqua (Denmark) and maintained in culture on a diet of the cryptophyte Rhodomonas salina for many generations.
New diatom batch cultures were established from inoculums of the above subcultures at initial densities of 16 (±6) × 106 µm3 ml−1 and maintained as described above. The growth rates, cellular silica contents, and copepod or dinoflagellate grazing on the diatoms were monitored every second day for several days. Because silica contents in diatoms scale approximately with their volume (figure 1a), we normalized cellular silica content with cell volume and computed the daily relative change in silica content as
| 2.1 |
where Si0 and Si1 are biogenic silica concentrations (fmol Si µm−3) at times t0 and t1.
Figure 1.

(a) Log–log relationship between biogenic silica content (pmol cell−1) and cell volume (µm3). The regression line is log(Si) = 0.87 log(V) – 2.89 (r2 = 0.84; p < 0.05). Data are from this study (average silica content and cell volume of diatoms grown under high light conditions) and [25,37–40]. (b) Relationship between relative change in biogenic silica content (SiR; d−1) and growth rate (GR; d−1). The regression line (with 95% confidence limits) is SiR = –0.29GR + 0.02 (r2 = 0.6; p < 0.05). Details about experimental organisms are provided in table 1. HL, high light conditions; LL, low light conditions. (Online version in colour.)
(b). Copepod and nauplii grazing experiments
Every second day (days 1, 3, 5, 7 and 9), ingestion rates of T. longicornis adult copepods and nauplii on diatoms were measured. To separate the size and silica effects on grazing, the ingestion rates of nauplii were additionally measured on small non-silicified nanoflagellates for 6 days (days 1, 3 and 5) (table 1). Prey suspensions at concentrations of 3.5 × 106 and 5 × 105 µm3 ml−1, respectively, were prepared in B1 + Si medium. For each diatom species and treatment (low and high light), the prey suspension was added to nine 68 ml glass bottles: triplicate initials, triplicate controls (without grazers) and triplicate grazing bottles (with grazers). From each initial bottle, a subsample of 5 ml was removed for cell enumeration and fixed with acidified Lugol's solution; a subsample of 10 ml was withdrawn for the biogenic silica analysis and treated as described below; and a subsample of 15 ml for measurement of the cell volume. On the experimental days, 15 copepods or 40 nauplii that had starved for 24 h were added to each grazing bottle, and control and grazing bottles were then mounted on a slowly rotating (0.5 rpm) plankton wheel (diameter of 37 cm) for 24 h in darkness and at a temperature of 15°C. At the end of the incubation, subsamples of 5 ml for cell enumeration were fixed with acidified Lugol's solution. The number of copepods or nauplii was counted and their ingestion rates calculated according to [41].
The remaining volume from the grazing bottles with adult copepods was dispensed into culture plates, and approximately 30 faecal pellets from each bottle picked under a dissecting microscope. Faecal pellets were immediately examined under a fluorescence microscope (Olympus, blue filter, magnification 200×), where the number of viable cells per faecal pellet was counted. Afterwards, the remaining contents were fixed with acidified Lugol's solution, and the number of faecal pellets determined. Survival of diatoms during gut passage was calculated as a fraction of ingested diatoms that survived gut passage.
(c). Dinoflagellate grazing experiment
Predation by the dinoflagellate Oxyrrhis marina was measured on four diatom species every second day for 10 days (days 1, 3, 5, 7 and 9) and on nanoflagellates for 4 days (days 1 and 3). Diatom and nanoflagellate prey suspensions at concentrations of 1 × 106 µm3 ml−1 were prepared in B1 + Si and B1 medium, respectively. The prey suspensions were then dispensed in twelve 68 ml glass bottles: six controls and six grazing bottles with 100 dinoflagellates ml−1. Three bottles from each treatment were immediately harvested for determination of initial prey and grazer concentrations as well as biogenic silica content in diatoms. All the other bottles were mounted on a plankton wheel and harvested at the end of the 24 h incubation. The ingestion rates of dinoflagellates were calculated according to [41,42].
(d). Biogenic silica analysis
The total cellular silica content (biogenic silica) was determined according to [37]. Briefly, 10 ml subsamples were filtered on polycarbonate filters with a pore size of 3 µm, and washed twice with acidic MilliQ water (pH of MilliQ was adjusted to 5.0 using concentrated hydrochloric acid to minimize dissolution of silica). Filters were then placed in 50 ml sterile polyethylene tubes, dried at 65°C for 90 min, and stored at −20°C until analysis. Dissolution of biogenic silica was later achieved by adding 15 ml of 0.5% (w/v) sodium carbonate solution to the tubes and heating to 85°C for 90 min. When cool, the pH was adjusted to 5.0–6.0 by adding concentrated sulfuric acid. The reactive silicate concentration was determined according to the molybdate method of [43]) using a SmartChem 200 wet chemistry analyser (Unity Scientific, MA).
(e). Statistical analysis
R package lmerTest [44] was used to perform a linear mixed-effects analysis on the relationship between ingestion rates, silica content and size, with species as random effects. The Akaike information criterion (AIC) was used to select the best model. The model was validated by visual inspection of residual plots.
3. Results
(a). Growth and biogenic silica content
Our data together with the data compiled from the literature show that the silica content is nearly proportional to the cell volume (figure 1a). Hence, we normalized cellular silica content (i.e. pmol cell−1) with cell volumes (i.e. pmol µm−3).
Diatoms cultured at low light intensities experienced slower growth compared to those cultured under high light conditions (electronic supplementary material, figure S1). The silica contents of all diatom species decreased during the development of the cultures, and were in most cases higher in light-limited than in light-saturated cells (electronic supplementary material, figure S2). The silica content of cells was not related to their growth rate (electronic supplementary material, figure S3A) because cultures never achieved steady state, but the rate of change in silica content declined with the growth rate of diatoms (figure 1b). Overall, we obtained diatoms with a sixfold variation in silica content among species, and a nearly twofold variation within species.
(b). Grazing on diatoms with different degrees of silicification
Ingestion rates of the adult copepod T. longicornis decreased with increasing silica content of diatoms when compared across species, and varied approximately inversely with the silica content (hyperbolic relation) (figure 2a). A hyperbolic relationship implies that grazing mortality initially decreases rapidly with increasing silica content but as silica content increases further the mortality reduction diminishes. Overall, a sixfold variation in silica content from 1 to 6 fmol Si µm−3 lead to a fourfold decrease in grazing mortality. The same pattern of reduced grazing mortality with increasing silica content was also found within all species and was significant in four out of seven cases.
Figure 2.

(a) Log–log relationship between copepod ingestion rate (IR; cell volume copepod−1 h−1) and biogenic silica content (Si; fmol µm−3), and (b) between copepod ingestion rate and cell size (size; µm). See table 2 for the model selection. (c) Relationship between nauplii ingestion rate (IR; cell volume nauplius−1 h−1) and biogenic silica content (Si; fmol µm−3). A nonlinear regression was fitted to the silica range: IR = 19360 × exp(−0.81Si) [r2 = 0.71; p < 0.05]; long-dashed lines are 95% confidence intervals. (d) Relationship between nauplii ingestion rate and cell size (size; µm) (p > 0.05). (e) Relationship between dinoflagellate ingestion rate (IR; cell volume dinoflagellate−1 h−1) and biogenic silica content (fmol µm−3) (p > 0.05). (f) Relationship between dinoflagellate ingestion rate and prey size (size; µm). A log-normal distribution was fitted to the prey size spectrum: IR = 309.1 × exp[−0.5 × ((log(size) – 7.7)/2.9)2] (r2 = 0.55; p < 0.05); long-dashed lines are 95% confidence intervals. Details about experimental organisms are provided in table 1. HL, high light conditions; LL, low light conditions. (Online version in colour.)
Cell size was a confounding factor since ingestion rates also varied with prey size, although much less than with silica content (figure 2b). Among the tested models for the ingestion rates, the AIC indicated that the best fit was with silica content per volume as a fixed factor, and diatom species as a random effect (table 2).
Table 2.
Model comparisons of ingestion rates log(IR), where cell size log(size), silica content per volume log(Si) or silica content per volume and cell size log(Si)+log(size) are fixed factors, and species is a random effect. Selected model is presented in italics. *p < 0.05; **p < 0.01; and ***p < 0.001.
| model | response variable | fixed structure | random structure | slope | confidence intervals | AIC |
|---|---|---|---|---|---|---|
| 1 | log(IR) | log(size)** | species*** | 1.67 | [0.58, 2.76] | 4.6 |
| 2 | log(IR) | log(Si)*** | species** | −0.79 | [−0.98, −0.60] | −23.7 |
| 3 | log(IR) | log(Si)*** + log(size) | species** | −0.70 0.32 |
[−0.91, −0.48] [−0.32, 0.95] |
−23.1 |
Similarly, ingestion rates of the copepod nauplii decreased with increasing silica content of diatoms both within and among the species (figure 2c). Overall, less than a threefold increase in silica content led to almost negligible ingestion rates. Moreover, prey size variation within the examined range did not affect the ingestion rates of the nauplii (figure 2d).
Grazing of the dinoflagellate Oxyrrhis marina on diatoms and flagellates was unaffected by silica content, and was similar between the two prey groups (non-silicified nanoflagellates versus diatoms) (figure 2e). On the other hand, the grazing rates significantly varied with prey size (figure 2f).
(c). Survival of ingested diatoms
A small fraction of the ingested diatoms survived gut passage intact (up to approx. 0.5%); however, survival of ingested diatoms was independent of biogenic silica content (figure 3a) and/or size of diatoms (figure 3b).
Figure 3.

(a) Relationship between fraction of viable cells in copepod faecal pellets (survivors; %) and biogenic silica content of ingested cells (fmol µm−3) (p > 0.05); and (b) between viable cells and size of diatoms (µm) (p > 0.05). Details about experimental organisms are provided in table 1. HL, high light conditions; LL, low light conditions. (Online version in colour.)
4. Discussion
(a). Protective value of silicified cell walls
Here, we have shown that the siliceous shell provides partial protection against copepod grazing, and that the protective value increases with the silica content in the cells (figure 2a,c). The relations found here are, of course, specific to the grazers used, and smaller or larger copepod species may be more or less constrained by the silica shell than found here—just as the small nauplii are more impacted by the shell thickening than the larger adults are. The fact that very few cells pass the gut intact suggests that even small cells are broken during or after ingestion, and this is consistent with mechanical protection. This may also explain why the less powerful copepod nauplii, in particular, were unable to consume heavy silicified cells. Note that the differences in silicification among species can be higher than those obtained in our experiments [25], and therefore expected differences in predation mortality from copepods can be even more drastic than observed here. Similar protective value of silicification may be expected against euphausiids, which also need to crush diatoms before ingestion.
Other major metazoan grazers of diatoms in the ocean are tunicates. Tunicates are considered unselective feeders that ingest all prey within their food size spectrum. However, unlike copepods, consumed diatoms are not broken during ingestion and digestion, and thus the silica frustule will allow higher survival in tunicate faecal pellets [45].
This dependency of copepod grazing mortality on silica content is consistent with experimental measurements as well as finite-element method (FEM) simulations, demonstrating that the mechanical strength of diatom shells increases with increasing silica content [23,29,46,47]. It is also consistent with scaling arguments (see below) that both the fracture strength and the critical buckling load of a diatom shell increase approximately with the cell wall thickness raised to the power of 3. This scaling additionally explains, in a general sense, why the advantage of further thickening decreases as the silica content increases: the un-breakability of cells increases rapidly with their silica content.
While the diatom wall reduces copepod grazing mortality in our experiments, the wall seems to provide no protection against the protistan grazer Oxyrrhis marina (figure 2e). Phagotrophic dinoflagellates display a variety of feeding mechanisms, such as direct engulfment of prey, pallium feeding and tube-feeding mechanisms [48], but regardless of the strategy, the ability of dinoflagellates to use a specific prey is governed mainly by the size and shape of the prey [49]. For pallium feeders and those that engulf their prey, such as Oxyrrhis marina, crushing resistance of the silicified cell walls is of no importance, as these organisms digest the cell contents without damaging the walls [8]. It has been speculated that the wall may increase the prey handling time and hence decrease feeding rate [7], but we did not find any significant effects. On the other hand, a silicified wall may be an obstacle to tube-feeding dinoflagellates that pierce the prey by means of a peduncle and suck out its contents, and therefore it is not surprising that these dinoflagellates predominantly feed on naked cells [22,50]. However, there are some parasitoid peduncle-feeding dinoflagellates that successfully feed on diatoms, although their host range is generally restricted to closely related species [50], possibly indicating coevolution of silica walls and attack strategies based on the mode of accessing the frustule [22].
Thus, the defensive role of a silicified cell wall in diatoms appears to primarily provide partial protection against grazers that crush the cells. Below we discuss how the mechanical properties of silica shells vary with cell size and silica content in the context of copepod predation. We next discuss the cost of silicification and explore the trade-off function (i.e. the relation between the costs and benefits of the siliceous shell).
(b). Size scaling of the mechanical properties of the diatom shell
The material properties of the diatom shell are unique: it has the highest mechanical strength relative to its density among any known biological material, and for diatom shells this ratio is close to the theoretical maximum [51]. In addition to several other potential functions of the shell—to support the vacuole [52], enhance nutrient uptake rate [10] and facilitate light harvesting [53,54]—its outstanding strength strongly suggests that it also has a protective and defensive function, as demonstrated here. The silica content of diatoms scales approximately with cell volume (figure 1a), demonstrating that the thickness of the shell increases with cell size. As rationalized below, the observed scaling is further evidence that the shell mainly serves a protective function. We can explore the scaling by asking how the silica content must increase with cell size to be able to resist the same force before it cracks or buckles. For this crude analysis, we ignore structural differences in the shell among species.
For a thin-walled structure (shell thickness h, size L and surface area A ≈ L2), the silica wall mass W scales with its volume as
| 4.1 |
Several criteria may be derived for the mechanical strength, such as maximal (fracture) stress, or the critical buckling load. Assume first that failure is due to fracture. Consider a ring of radius R and cross section h × h that is subject to a centrally directed force F, and apply the Bernoulli–Euler beam equation. The ring would experience a maximum moment M = RF/π and maximal stress σ = 6RF/πh3, implying for R ≈ L that h ≈ L1/3. If this relation, on grounds of dimensional analysis, also applies to a sphere, spheroid or cylinder, then equation (4.2) leads to the scaling law
| 4.2 |
With similar arguments, the exact same scaling arises if one considers a plate that is subject to a central force applied by a bar, maybe better resembling the mandible acting on a diatom cell. Next, consider instead that failure is due to buckling. The buckling load P for a column (cross section h × h and length L) is P = π2EI/4L2, where E is Young's modulus, and the moment of inertia I scales as I ≈ h4 (as derived by Euler in 1755 [55]). This implies I/L2 ≈ h4/L2 ≈ constant or h ≈ L1/2. Attempting to approach the morphology of a diatom shell, assume the column has a cross-section h × b with b ≫ h (i.e. a plate of thickness h), then the buckling load would scale as P ≈ h3b/L2 ≈ h3 just as suggested by the FEM simulation of [47]. For this case, using the criterion P ≈ h3b/L2 ≈ constant (i.e. h ≈ L2/3), equation (4.2) now leads to the scaling law
| 4.3 |
where V is the cell volume (assumed proportional to the cube of linear dimension of the cell, L3). Incidentally, this is identical to the observed size scaling of silica content in diatoms (figure 1a). Both arguments therefore suggest that shell thickness should increase with size to retain constant resistance to external forces, thus further supporting the adaptive value of the shell as a defensive structure.
(c). Trade-offs
While our experiments demonstrate the benefits of the silica wall in terms of reduced predation mortality, the actual magnitude of the benefit of course depends linearly on the concentration of grazers. However, there may be costs of the defence. These costs are not immediately obvious from our experiments, but may nevertheless be explored from available information. The biochemical costs of forming a siliceous wall seem to be negligibly small; in fact, building a silicified cell wall appears to be much cheaper than the formation of ordinary cell walls [33]. There is evidence that shell thickening can be induced by grazer cues [5], and that thickening of the silica wall may require reduced cell division rate, suggesting a possible trade-off. This is because the incorporation of silica is linked to the duration of the cell wall synthesis phase in the cell division cycle. Since slow growth prolongs this phase, this will lead to higher incorporation of silica in the frustule [32]. As a consequence, silica content in diatoms will decline with growth rate under non-limiting silica conditions, as demonstrated repeatedly [32,35,37,56]. If the silica deposition rate was constant and independent of cell division rate, the silica content at steady state would vary inversely with growth rate (i.e. growth rate raised to the power of −1). Because the mortality rate due to copepod grazing in our experiments varies approximately inversely with silica content, this would imply a linear relationship between growth rate and grazing mortality (i.e. suggesting a linear trade-off). However, data from the literature suggest that silica incorporation rate does depend on cell division rate and that the steady-state silica content rather varies with the cell division rate raised to a power of b that is smaller than −1 (figure 4). From the five experiments reported in figure 4, an average slope of −0.54 is estimated. This, in turn, implies that the grazing mortality increases with the growth rate raised to the power of 0.43 (when IR ∼ Si−0.79 from table 2). This suggests that as growth decreases, the reduction in predation mortality accelerates.
Figure 4.
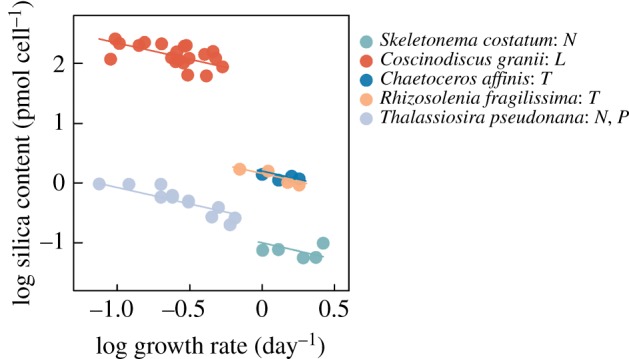
Steady-state silica content in diatoms as a function of cell division rate for diatoms that are limited by factors other than silica (nitrogen N, phosphorus P, light L or temperature T, as indicated in the legend). Data obtained from [32,35,37,56]. A fitted mixed linear model with one slope and random intercepts was fitted to the log–log relationships between silica content and growth rate. The estimated slope is −0.54 [−0.36, −0.72]. (Online version in colour.)
While the above demonstrates how shell thickness is a ‘passive’ function of cell division rate, there is evidence for one species, Thallasiosira weissflogii, that shell thickening may be induced by grazer cues [5]. If this observation can be generalized, it remains to be demonstrated whether and to what extent induced shell thickening requires a reduced growth rate, as suggested by the above, or whether the thickening is the result of an increased rate of silica incorporation in the frustule. However, inducible defences suggest that the defence is costly [57], and those costs must eventually lead to a reduced cell division rate. More experiments are required to clarify this and to quantify the trade-off function between the benefits and costs of induced thickening of cell walls in diatoms.
(d). Survival of diatoms during gut passage
Previous studies reported intact diatoms in copepod faecal pellets, suggesting that the silica wall may enable viable gut passage of ingested diatoms [18,19]. Our experiments show, however, that only a minute fraction of the cells survive and that the survival is independent of the silica content in the cells (figure 3a). However, the number of live cells may have been underestimated in our study, as some viable cells are not auto-fluorescent [58]. The few cells that do survive gut passage may sink to the seafloor together with the faecal pellet and thus be removed from the pelagic population. Sinking and overwintering in the sediment are an integral part of the diatom life-cycle strategy [34], and sedimented cells contribute to reseeding the pelagic the following year when brought back to the surface by upwelling or vertical mixing [9].
5. Concluding remarks
Diatoms are the major contributors to phytoplankton blooms because their mortality rates are lower than those of other groups with similar growth rates, making them central in aquatic ecosystems as well as the global carbon cycle [46]. But what makes diatoms unique are their silicified cell walls, whose mechanical strength relative to density is the highest among any known biological material [51]. Our results demonstrate that the silicified cell wall truly is an effective morphological defence mechanism against copepods, whose opal teeth have probably coevolved with their prey, but ineffective against protozoans with direct engulfment as feeding mechanisms. However, a specific trait does not need to perform perfectly to be maintained in a population, and if the grazing mortality rate is decreased by even a small factor, then as long as this benefit offsets the costs of employing the trait, it can be retained [8].
Supplementary Material
Supplementary Material
Acknowledgements
We thank Hans van Someren Gréve for kindly providing graphics of zooplankton for figure 2. We also thank Poul Scheel Larsen for help with the mechanical modelling of the silica wall, and Aurore Maureaud for providing the R script for linear mixed effects analysis and valuable discussions. The Centre for Ocean Life is a Villum Kahn Rasmussen Centre for Excellence funded by the Villum Foundation.
Data accessibility
Data available as part of the electronic supplementary material.
Competing interests
We declare we have no competing interests.
Funding
R.A. was funded through a DFF-Individual postdoctoral grant no. (17023) from the Danish Council for Independent Research, a Marie Curie Intra-European fellowship from the People Programme of the European Union's Seventh Framework Programme FP7/2007–2013/under REA grant agreement no. 6240979 and an H. C. Ørsted Postdoctoral fellowship from DTU. Additional support was received from the Gordon & Betty Moore Foundation through award no. 5479.
References
- 1.Mortensen B. 2013. Plant resistance against herbivory. Nat. Educ. Knowl. 4, 1–5. [Google Scholar]
- 2.Pančić M, Kiørboe T. 2018. Phytoplankton defence mechanisms: traits and trade-offs. Biol. Rev. 93, 1269–1303. ( 10.1111/brv.12395) [DOI] [PubMed] [Google Scholar]
- 3.Jakobsen HH, Tang KW. 2002. Effects of protozoan grazing on colony formation in Phaeocystis globosa (Prymnesiophyceae) and the potential costs and benefits. Aquat. Microb. Ecol. 27, 261–273. ( 10.3354/ame027261) [DOI] [Google Scholar]
- 4.Selander E, Thor P, Toth G, Pavia H. 2006. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B 273, 1673–1680. ( 10.1098/rspb.2006.3502) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pondaven P, Gallinari M, Chollet S, Bucciarelli E, Sarthou G, Schultes S, Jean F. 2007. Grazing-induced changes in cell wall silicification in a marine diatom. Protist 158, 21–28. ( 10.1016/j.protis.2006.09.002) [DOI] [PubMed] [Google Scholar]
- 6.Harðardóttir S, Pančić M, Tammilehto A, Krock B, Møller EF, Nielsen TG, Lundholm N. 2015. Dangerous relations in the Arctic marine food web: interactions between toxin producing Pseudo-nitzschia diatoms and Calanus copepodites. Mar. Drugs 13, 3809–3835. ( 10.3390/md13063809) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Smetacek V. 1999. Diatoms and the ocean carbon cycle. Protist 150, 25–32. ( 10.1016/S1434-4610(99)70006-4) [DOI] [PubMed] [Google Scholar]
- 8.Finkel ZV, Kotrc B. 2010. Silica use through time: macroevolutionary change in the morphology of the diatom fustule. Geomicrobiol. J. 27, 596–608. ( 10.1080/01490451003702941) [DOI] [Google Scholar]
- 9.Raven JA, Waite AM. 2004. The evolution of silicification in diatoms: inescapable sinking and sinking as escape? New Phytol. 162, 45–61. ( 10.1111/j.1469-8137.2004.01022.x) [DOI] [Google Scholar]
- 10.Mitchell JG, Seuront L, Doubell MJ, Losic D, Voelcker NH, Seymour J, Lal R. 2013. The role of diatom nanostructures in biasing diffusion to improve uptake in a patchy nutrient environment. PLoS ONE 8, 1–9. ( 10.1371/journal.pone.0059548) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Romann J, Valmalette J-C, Chauton MS, Tranell G, Einarsrud M-A, Vadstein O. 2015. Wavelength and orientation dependent capture of light by diatom frustule nanostructures. Sci. Rep. 5, 1–6. ( 10.1038/srep17403) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Aguirre LE, Ouyang L, Elfwing A, Hedblom M, Wulff A, Inganäs O. 2018. Diatom frustules protect DNA from ultraviolet light. Sci. Rep. 8, 1–6. ( 10.1038/s41598-018-21810-2) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hartley SE, DeGabriel JL. 2016. The ecology of herbivore-induced silicon defences in grasses. Funct. Ecol. 30, 1311–1322. ( 10.1111/1365-2435.12706) [DOI] [Google Scholar]
- 14.Sherr EB, Sherr BF. 2007. Heterotrophic dinoflagellates: a significant component of microzooplankton biomass and major grazers of diatoms in the sea. Mar. Ecol. Prog. Ser. 352, 187–197. ( 10.3354/meps07161) [DOI] [Google Scholar]
- 15.Hansen PJ. 1991. Quantitative importance and trophic role of heterotrophic dinoflagellates in a coastal pelagial food web. Mar. Ecol. Prog. Ser. 73, 253–261. [Google Scholar]
- 16.Thompson GA, Dinofrio EO, Alder VA. 2013. Structure, abundance and biomass size spectra of copepods and other zooplankton communities in upper waters of the Southwestern Atlantic Ocean during summer. J. Plankton Res. 35, 610–629. ( 10.1093/plankt/fbt014) [DOI] [Google Scholar]
- 17.Miller CB, Nelson DM, Weiss C, Soeldner AH. 1990. Morphogenesis of opal teeth in calanoid copepods. Mar. Biol. 106, 91–101. ( 10.1007/BF02114678) [DOI] [Google Scholar]
- 18.Fowler SW, Fisher NS. 1983. Viability of marine phytoplankton in zooplankton fecal pellets. Deep Sea Res. Part A, Oceanogr. Res. Pap. 30, 963–969. ( 10.1016/0198-0149(83)90051-1) [DOI] [Google Scholar]
- 19.Jansen S, Bathmann U. 2007. Algae viability within copepod faecal pellets: evidence from microscopic examinations. Mar. Ecol. Prog. Ser. 337, 145–153. ( 10.3354/meps337145) [DOI] [Google Scholar]
- 20.Kruse S, Jansen S, Kraegefsky S, Bathmann U. 2009. Gut content analyses of three dominant Antarctic copepod species during an induced phytoplankton bloom EIFEX (European iron fertilization experiment). Mar. Ecol. Evol. Perspect. 30, 301–312. ( 10.1111/j.1439-0485.2009.00284.x) [DOI] [Google Scholar]
- 21.Michels J, Vogt J, Gorb SN. 2012. Tools for crushing diatoms: opal teeth in copepods feature a rubber-like bearing composed of resilin. Sci. Rep. 2, 1–6. ( 10.1038/srep00465) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hamm C, Smetacek V. 2007. Armor: why, when, and how. In Evolution of primary producers in the sea (eds Falkowski PG, Knoll AH), pp. 311–332. New York, NY: Academic Press. [Google Scholar]
- 23.Friedrichs L, Hoernig M, Schulze L, Bertram A, Jansen S, Hamm C. 2013. Size and biomechanic properties of diatom frustules influence food uptake by copepods. Mar. Ecol. Prog. Ser. 481, 41–51. ( 10.3354/meps10227) [DOI] [Google Scholar]
- 24.Zhang S, Liu H, Ke Y, Li B. 2017. Effect of the silica content of diatoms on protozoan grazing. Front. Mar. Sci. 4, 202 ( 10.3389/fmars.2017.00202) [DOI] [Google Scholar]
- 25.Brzezinski MA. 1985. The Si : C : N ratio of marine diatoms: interspecific variability and the effect of some environmental variables. J. Phycol. 21, 347–357. ( 10.1111/j.0022-3646.1985.00347.x) [DOI] [Google Scholar]
- 26.Claquin P, Martin-Jézéquel V, Kromkamp JC, Veldhuis MJW, Kraay GW. 2002. Uncoupling of silicon compared with carbon and nitrogen metabolisms and the role of the cell cycle in continuous cultures of Thalassiosira pseudonana (Bacillariophyceae) under light, nitrogen, and phosphorus control. J. Phycol. 38, 922–930. ( 10.1046/j.1529-8817.2002.t01-1-01220.x) [DOI] [Google Scholar]
- 27.De La Rocha CL, Hutchins DA, Brzezinski MA, Zhang Y.. 2000. Effects of iron and zinc deficiency on elemental composition and silica production by diatoms. Mar. Ecol. Prog. Ser. 195, 71–79. ( 10.3354/meps195071) [DOI] [Google Scholar]
- 28.Furnas M. 1978. Influence of temperature and cell size on the division rate and chemical content of the diatom Chaetoceros curvisetum Cleve. J. Exp. Mar. Bio. Ecol. 34, 97–109. ( 10.1016/0022-0981(78)90034-5) [DOI] [Google Scholar]
- 29.Wilken S, Hoffmann B, Hersch N, Kirchgessner N, Dieluweit S, Rubner W, Hoffmann LJ, Merkel R, Peeken I. 2011. Diatom frustules show increased mechanical strength and altered valve morphology under iron limitation. Limnol. Oceanogr. 56, 1399–1410. ( 10.4319/lo.2011.56.4.1399) [DOI] [Google Scholar]
- 30.Liu H, Chen M, Zhu F, Harrison PJ. 2016. Effect of diatom silica content on copepod grazing, growth and reproduction. Front. Mar. Sci. 3, 1–7. ( 10.3389/fmars.2016.00089) [DOI] [Google Scholar]
- 31.Brzezinski MA, Olson RJ, Chisholm SW. 1990. Silicon availability and cell-cycle progression in marine diatoms. Mar. Ecol. Prog. Ser. 67, 83–96. ( 10.3354/meps067083) [DOI] [Google Scholar]
- 32.Martin-Jézéquel V, Hildebrand M, Brzezinski MA. 2000. Silicon metabolism in diatoms: implications for growth. J. Phycol. 36, 821–840. ( 10.1046/j.1529-8817.2000.00019.x) [DOI] [Google Scholar]
- 33.Raven JA. 1983. The transport and function of silicon in plants. Biol. Rev. 58, 179–207. ( 10.1111/j.1469-185X.1983.tb00385.x) [DOI] [Google Scholar]
- 34.Smetacek VS. 1985. Role of sinking in diatom life-history cycles: ecological, evolutionary and geological significance. Mar. Biol. 84, 239–251. ( 10.1007/BF00392493) [DOI] [Google Scholar]
- 35.Taylor NJ. 1985. Silica incorporation in the diatom Cosinodiscus granii as affected by light intensity. Br. Phycol. J. 20, 365–374. ( 10.1080/00071618500650371) [DOI] [Google Scholar]
- 36.Hansen PJ. 1989. The red tide dinoflagellate Alexandrium tamarense: effects on behaviour and growth of a tintinnid ciliate. Mar. Ecol. Prog. Ser. 53, 105–116. ( 10.3354/meps053105) [DOI] [Google Scholar]
- 37.Paasche E. 1980. Silicon content of five marine plankton diatom species measured with a rapid filter method. Limnol. Oceanogr. 25, 474–480. ( 10.4319/lo.1980.25.3.0474) [DOI] [Google Scholar]
- 38.Eppley RW, Holmes RW, Paasche E. 1967. Periodicity in cell division and physiological behavior of Ditylum brightwellii, a marine planktonic diatom, during growth in light-dark cycles. Arch. Mikrobiol. 56, 305–323. ( 10.1007/BF00425206) [DOI] [Google Scholar]
- 39.Harrison PJ, Conway HL, Holmes RW, Dugdale RC. 1977. Marine diatoms grown in chemostats under silicate or ammonium limitation. III. Cellular chemical composition and morphology of Chaetoceros debilis, Skeletonema costatum, and Thalassiosira gravida. Mar. Biol. 43, 19–31. ( 10.1007/BF00390940) [DOI] [Google Scholar]
- 40.Parsons TR, Stephens K, Strickland JDH. 1961. On the chemical composition of eleven species of marine phytoplankters. J. Fish. Res. Board Canada 18, 1001–1016. ( 10.1139/f61-063) [DOI] [Google Scholar]
- 41.Frost BW. 1972. Effects of size and concentration of food particles on the feeding behavior of the marine planktonic copepod Calanus pacificus. Limnol. Oceanogr. 17, 805–815. ( 10.4319/lo.1972.17.6.0805) [DOI] [Google Scholar]
- 42.Heinbokel JF. 1978. Studies on the functional role of tintinnids in the Southern California Bight. I. Grazing and growth rates in laboratory cultures. Mar. Biol. 47, 177–189. ( 10.1007/BF00395638) [DOI] [Google Scholar]
- 43.Strickland JDH, Parsons TR.. 1972. A practical handbook of seawater analysis. London, ON: Fisheries Research Board of Canada. [Google Scholar]
- 44.Kuznetsova A, Brockhoff PB, Christensen RHB. 2017. lmerTest package: tests in linear mixed effects models. J. Stat. Softw. 82, 1–26. ( 10.18637/jss.v082.i13) [DOI] [Google Scholar]
- 45.Paffenhöfer GA, Köster M. 2005. Digestion of diatoms by planktonic copepods and doliolids. Mar. Ecol. Prog. Ser. 297, 303–310. ( 10.3354/meps297303) [DOI] [Google Scholar]
- 46.Hamm CE, Merkel R, Springer O, Jurkojc P, Maier C, Prechtel K, Smetacek V. 2003. Architecture and material properties of diatom shells provide effective mechanical protection. Nature 421, 841–843. ( 10.1038/nature01416) [DOI] [PubMed] [Google Scholar]
- 47.Gutiérrez A, Gordon R, Dávila LP. 2017. Deformation modes and structural response of diatom frustules. J. Mater. Sci. Eng. Adv. Technol. 15, 105–134. ( 10.18642/jmseat_7100121810) [DOI] [Google Scholar]
- 48.Hansen PJ. 1992. Prey size selection, feeding rates and growth dynamics of heterotrophic dinoflagellates with special emphasis on Gyrodinium spirale. Mar. Biol. 114, 327–334. ( 10.1007/BF00349535) [DOI] [Google Scholar]
- 49.Tillmann U. 2004. Interactions between planktonic microalgae and protozoan grazers. J. Eukaryot. Microbiol. 51, 156–168. ( 10.1111/j.1550-7408.2004.tb00540.x) [DOI] [PubMed] [Google Scholar]
- 50.Kühn SF, Medlin LK. 2005. The systematic position of the parasitoid marine dinoflagellate Paulsenella vonstoschii (Dinophyceae) inferred from nuclear-encoded small subunit ribosomal DNA. Protist 156, 393–398. ( 10.1016/J.PROTIS.2005.09.002) [DOI] [PubMed] [Google Scholar]
- 51.Aitken ZH, Luo S, Reynolds SN, Thaulow C, Greer JR. 2016. Microstructure provides insights into evolutionary design and resilience of Coscinodiscus sp. frustule. Proc. Natl Acad. Sci. USA 113, 2017–2022. ( 10.1073/PNAS.1519790113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Thingstad TF, Øvreås L, Egge JK, Løvdal T, Heldal M. 2005. Use of non-limiting substrates to increase size; a generic strategy to simultaneously optimize uptake and minimize predation in pelagic osmotrophs? Ecol. Lett. 8, 675–682. ( 10.1111/j.1461-0248.2005.00768.x) [DOI] [Google Scholar]
- 53.Fuhrmann T, Landwehr S, El Rharbi-Kucki M, Sumper M. 2004. Diatoms as living photonic crystals. Appl. Phys. B 78, 257–260. ( 10.1007/s00340-004-1419-4) [DOI] [Google Scholar]
- 54.De Tommasi E, Gielis J, Rogato A.. 2017. Diatom frustule morphogenesis and function: a multidisciplinary survey. Mar. Genomics 35, 1–18. ( 10.1016/j.margen.2017.07.001) [DOI] [PubMed] [Google Scholar]
- 55.Euler L. 1755. Principes generaux du mouvement des fluides. Hist. l'Académie des Sci. des B.-lett. Berlin11, E226.
- 56.Harrison PJ, Conway HL, Dugdale RC. 1976. Marine diatoms grown in chemostats under silicate or ammonium limitation. I. Cellular chemical composition and steady state growth kinetics of Skeletonema costatum. Mar. Biol. 35, 177–186. ( 10.1007/BF00390939) [DOI] [Google Scholar]
- 57.Karban R. 2011. The ecology and evolution of induced resistance against herbivores. Funct. Ecol. 25, 339–347. ( 10.1111/j.1365-2435.2010.01789.x) [DOI] [Google Scholar]
- 58.Zetsche E-M, El Mallahi A, Meysman FJR. 2016. Digital holographic microscopy: a novel tool to study the morphology, physiology and ecology of diatoms. Diatom Res. 31, 1–16. ( 10.1080/0269249X.2016.1140679) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data available as part of the electronic supplementary material.