

Abstract
Background
Few studies have evaluated the relative cross-protection conferred by infection with different groups of viruses through studies of sequential infections in humans. We investigated the presence of short-lived relative cross-protection conferred by specific prior viral infections against subsequent febrile respiratory illness (FRI).
Methods
Men enlisted in basic military training between December 2009 and December 2014 were recruited, with the first FRI as the study entry point. ResPlex II assays and real-time polymerase chain reaction assays were used to detect viral pathogens in nasal wash samples, and survival analyses were performed to determine whether infection with particular viruses conferred short-lived relative cross-protection against FRI.
Results
Prior infection with adenovirus (hazard ratio [HR], 0.24; 95% confidence interval [CI], .14–.44) or influenza virus (HR, 0.52; 95% CI, .38–.73) conferred relative protection against subsequent FRI episode. Results were statistically significant even after adjustment for the interval between enlistment and FRI (P < .001). Adenovirus-positive participants with FRI episodes tended to be protected against subsequent infection with adenovirus, coronavirus, enterovirus/rhinovirus, and influenza virus (P = .062–.093), while men with influenza virus–positive FRI episodes tended be protected against subsequent infection with adenovirus (P = .044) and influenza virus (P = .081).
Conclusion
Prior adenovirus or influenza virus infection conferred cross-protection against subsequent FRI episodes relative to prior infection due to other circulating viruses.
Keywords: Antibodies, cell-mediated immunity, vaccine, cohort studies, surveillance, cross-protection, serology
Viral interference describes the phenomenon whereby a prior viral infection potentially exerts some effect on subsequent infection with other viruses [1]. Edward Jenner first reported this when herpetic infections were observed to cross-protect against the subsequent development of vaccinia lesions [2]. Then, in the 1950s, Isaacs and Lindenmann discovered the role of interferons in viral interference [3]. Since then, cross-protection has been observed for animal viruses [2], with possible mechanisms elaborated on in multiple animal models [4–6]. In human populations, time-series analyses have demonstrated how epidemics involving a particular virus influence the temporality of epidemics involving other families of respiratory viruses [7–10]. Several case-control and cross-sectional studies also show that co-detection of respiratory viruses is less frequently observed than if the infections caused by different respiratory viruses occur independently of each other [11–13]. However, evidence from studies of sequential infections in humans would provide more-robust evidence that viral infections can reduce the risk of subsequent infection with a different group of viruses and clarify whether such cross-protective effects differ between different types of viruses.
As previously reported, the rollout of routine trivalent seasonal influenza vaccination in the Singapore Armed Forces was accompanied by a dramatic decline in the incidence of laboratory-confirmed influenza virus infections but a much less noticeable decrease in the overall incidence of febrile respiratory illness (FRI) episodes [14]. A subsequent increase in the incidence of adenovirus infections appeared, at least from time-series data, to account for some degree of replacement for the influenza virus infections averted through vaccination [15]. In this article, we set out to clarify the relationship between specific prior viral infections and subsequent FRI episodes in young Singaporean men undergoing basic military training (BMT). We reanalyzed the Singapore Armed Forces FRI surveillance program data by using survival analyses to assess whether there was any evidence, at the individual level, that infection with particular viruses was conferring short-lived relative cross-protection against subsequent FRI episodes, including those caused by other virus groups.
METHODS
Study Setting and Data Collection
Since May 2009, the Singapore Armed Forces has operated a sentinel FRI surveillance program at a major recruit-training center. Recruits who develop FRI would report to the primary healthcare clinic for assessment, where they would receive treatment and would typically be given home leave for 1–2 days. During regular consultation hours, recruits meeting our inclusion criteria (ie, a temperature ≥37.5°C plus either cough or sore throat) were asked to participate in the study. Following receipt of written informed consent from the patient, we administered a questionnaire and collected nasal wash samples from both sides of the nose. We excluded repeat consultations if the patient was determined not to have recovered from the earlier illness episode.
Our study covered recruits undergoing BMT between 15 December 2009 and 31 December 2014, during which inactivated influenza vaccine was routinely administered to recruits who did not have any contraindications. This was initially a monovalent influenza vaccine (MIV) containing only 2009 pandemic influenza A(H1N1) virus (A[H1N1]pdm09), for recruits who enlisted between 11 December 2009 and 4 October 2010. The MIV was superseded by a trivalent influenza vaccine (TIV) that included A(H1N1)pdm09, for recruits who enlisted between 8 November 2010 and 10 December 2014).
The study was reviewed and approved by the Singapore military’s Joint Medical Committee for Research by and the National University of Singapore’s ethics review committee (National University of Singapore Institutional Review Board reference 09-255).
Laboratory Methods
Samples were sent in viral transport medium (Copan Diagnostics, Murrieta, CA) for etiological testing within 24 hours of collection. During the 5-year study period, the ResPlex II multiplex polymerase chain reaction assay was used from December 2009 through 29 June 2012 to detect viral pathogens, after which it was replaced by in-house viral multiplex polymerase chain reaction assays (Supplementary Materials) [16].
Study Design, Outcomes and Exposures of Interest, Potential Confounders, and Multilevel Framework
BMT is tailored to the physical, medical, and vocational needs of a recruit. The majority enlist into the main intake types, with standard durations of around 9, 17, and 19 weeks, with a small proportion having a mix of shorter courses (termed “others”). Dropout rates are low, and our study clinic served most of the healthcare needs for recruits at the training center. We were hence able to reanalyze the individual FRI episodes as a cohort study by linking consultation episodes through coded subject identifiers. The first FRI episode (FRI-1) for a given participant served as the point of entry into the study (t0). Individual participants then accumulated follow-up time (Figure 1A), with subsequent FRI episodes being the event of interest. Those with no further episodes accumulated follow-up time until their exit date from BMT, while participants with ≥2 episodes contributed additional follow-up intervals after each subsequent episode.
Figure 1.

A, Scenarios for febrile respiratory illness (FRI; defined as consultations at the primary healthcare clinic in which a temperature ≥37.5°C was detected, plus either cough or sore throat) episodes and intervals in time-to-event analyses. In scenario 1 (no subsequent FRI episode), a participant accumulates follow-up time from the first FRI episode (FRI-1) to the date of exit from basic military training (BMT). In scenario 2, follow-up time accrues between the first (FRI-1) and second (FRI-2) episodes (interval A, t1–t0), plus a second interval between FRI-2 and the date of exit from BMT (interval B, t2–t1). For ≥3 FRI episodes, additional intervals are used (eg, in scenario 3, interval C is t3–t2). B, FRI incidence rates by time from enlistment, with splitting of intervals into subintervals by BMT phase. Shown is an example based on a participant from the 17-week BMT intake. Interval A has a subinterval in weeks 11–12 of the BMT course and another in weeks 13–14 (with FRI incidences of 0.74 and 0.63 cases/1000 person-days, respectively). Likewise, interval B has 3 subintervals (weeks 13–14, 15–16, and 17–18). C, Levels within multilevel analysis. AdV, adenovirus; CoV, coronavirus; FluV, influenza virus; hMPV, human metapneumovirus; NA, not applicable; RSV, respiratory syncytial virus.
Other than subsequent consultation for any FRI episodes, additional definitions for events of interest were episodes positive for any of the respiratory viruses tested and specific groups of etiological agents. We grouped the agents by using broad categories that accommodated changes in testing protocol during the study period while reflecting virus taxonomy: adenoviruses (AdVs)—species B, E, and others for which the type was undetermined; coronaviruses (CoVs)—CoV-229E, CoV-NL63, CoV-HKU1, CoV-OC43, and other CoVs; enteroviruses and rhinoviruses (ERVs); human metapneumovirus (hMPV); influenza virus (FluV)—influenza A(H3N2) virus, 2009 pandemic influenza A(H1N1) virus (A[H1N1]pdm09), influenza A(H1N1) virus, influenza A virus–positive samples for which the subtype was undetermined, and influenza B virus; human parainfluenza virus (hPIV)—types 1–4; and respiratory syncytial virus (RSV)—RSV-A and RSV-B.
The virus categories described above were also our exposures of interest, to investigate whether an FRI episode caused by one virus group conferred relatively greater protection against a subsequent FRI episodes than FRI episodes testing negative for that virus group. In individuals with ≥1 subsequent FRI episode, exposure to prior infections was regarded as cumulative. For instance, for scenario 3 in Figure 1A, with 2 subsequent FRI episodes, hMPV exposure occurred only during interval C, CoV exposure occurred during intervals B and C, and AdV occurred during all 3 intervals.
Potential confounders included the year of the surveillance program, participant’s age and ethnicity, type of BMT intake, history of smoking and asthma (including childhood asthma), and receipt of influenza vaccine before the FRI episode. Assessment of whether the participant received MIV or TIV was based on self-reported vaccination history up to 1 year prior to the first FRI episode (since antibody titers and vaccine effectiveness could wane thereafter [17–19]), supplemented with Singapore Armed Forces records for vaccines received after enlistment. Since FRI episodes are concentrated differentially during a BMT course, we adjusted for this effect using the FRI incidence rate during successive 2-weekly phases after the time of enlistment for that intake type (Figure 1B and Supplementary Materials). Observation time was split into subintervals if the observation period straddled different BMT phases and was analyzed in a multilevel modeling framework (Figure 1C). The framework included random effects terms for the BMT companies that participants belonged to, within which were nested the intervals (delineated by consecutive FRI episodes), within which, in turn, were nested the subintervals straddling BMT phases (levels 4 to 1 for model 2). Models 1 and 3 only had 3 levels. Model 1 omitted the subintervals used to adjust for FRI incidence. Model 3, which stratified subintervals by whether they ended <4 weeks or ≥4 weeks after the initial FRI episode, used only the first FRI episode for each participant (episode level was therefore redundant). Models 2 and 3 adjusted for FRI incidence rates by time from enlistment and, hence, excluded the BMT intake type termed “other” (for which data used to estimate incidence rates were unavailable).
Statistical Analysis
We present simple tabulations and visualizations of the data at the participant and episode levels, using χ2 and Fisher exact tests where appropriate. We then investigated how FRI caused by specific virus families affected the subsequent risk of FRI episodes relative to those who were unexposed, through Kaplan-Meier plots and log-rank tests, as well as with multilevel survival analyses (with survival time modeled using a Weibull distribution), which adjusted for potential confounders. We also investigated whether exposure to FluV and AdV affected the subsequent risk of all FRI episodes and the risk of FRI by specific pathogens, using Kaplan-Meier plots and log-rank tests. However, multivariable analysis was not performed because of sample size limitations when restricting the event of interest to specific pathogens. All analyses used Stata 15 (Stata; College Station, TX), with 2-tailed P values of < .05 considered statistically significant.
RESULTS
Demographic Characteristics of Participants
The study included 6138 FRI episodes among 5677 participants (enrolled over approximately 5 years; Table 1), with a median of 33 days (interquartile range [IQR], 16–56 days) from enlistment to the first FRI episode and a median of 2 days (IQR, 1–3 days) from illness onset to the consultation. As inclusion in the study was based on having an FRI, the longer 17-week (2677 participants [47.2%]) and 19-week (1357 [23.9%]) BMT intake types contributed more participants, followed by the 9-week (1276 [22.5%]) and other (n = 367, 6.5%) intake types. The median age was 20.1 years (IQR, 19.0–20.9 years), and 99.9% were male. A total of 83.6% received influenza vaccine in the past 1 year (with only 113 receiving this before enlistment); 17.3% (981) received MIV only, 65.5% (3719) received TIV only, and 0.8% (47) received both MIV and TIV.
Table 1.
Characteristics of All Participants and Participants With ≥2 Febrile Respiratory Illness (FRI) Episodes
| Characteristic | All Participants, No. | Participants With ≥2 FRI Episodes, No. (%) | OR for Having ≥2 FRI Episodes (95% CI) | P a |
|---|---|---|---|---|
| Study year(s) | ||||
| 2009/2010 | 1510 | 127 (8.4) | Reference | <.001 |
| 2011 | 1315 | 140 (10.6) | 1.30 (1.01–1.67) | |
| 2012 | 1160 | 65 (5.6) | .65 (.47–.88) | |
| 2013 | 841 | 54 (6.4) | .75 (.54–1.04) | |
| 2014 | 851 | 37 (4.3) | .49 (.34–.72) | |
| Type of BMT intake | ||||
| 19-week | 1357 | 164 (12.1) | Reference | <.001 |
| 17-week | 2677 | 228 (8.5) | .68 (.55–.84) | |
| 9-week | 1276 | 24 (1.9) | .14 (.09–.22) | |
| Otherb | 367 | 7 (1.9) | .14 (.07–.30) | |
| Sex | ||||
| Male | 5672 | 423 (7.5) | Reference | 1.000 |
| Female | 5 | 0 (0.0) | … | |
| Ethnicity | ||||
| Chinese | 4362 | 304 (7.0) | Reference | .036 |
| Malay | 780 | 73 (9.4) | 1.38 (1.05–1.80) | |
| Indian | 322 | 32 (9.9) | 1.47 (1.00–2.16) | |
| Other | 213 | 14 (6.6) | .94 (.54–1.63) | |
| Smoking history | ||||
| Never | 4277 | 285 (6.7) | Reference | <.001 |
| Former | 147 | 18 (12.2) | 1.95 (1.18–3.25) | |
| Current | 1253 | 120 (9.6) | 1.48 (1.19–1.85) | |
| History of asthma | ||||
| No | 4340 | 326 (7.5) | Reference | .812 |
| Yes | 1337 | 97 (7.3) | .08 (.07–.09) | |
| Influenza vaccinationc | ||||
| No recent influenza vaccination | 930 | 97 (10.4) | Reference | .003 |
| MIV for A(H1N1)pdm09 only | 981 | 73 (7.4) | .69 (.50–.95) | |
| TIV only | 3719 | 250 (6.7) | .62 (.48–.79) | |
| Both MIV and TIV | 47 | 3 (6.4) | .59 (.18–1.92) |
Abbreviations: A(H1N1)pdm09, 2009 pandemic influenza A(H1N1) virus; BMT, basic military training; CI, confidence interval; MIV, monovalent influenza vaccine; OR, odds ratio; TIV, trivalent influenza vaccine.
aBy the Fisher exact test, comparing participants with 1 FRI episode to those with >1 FRI episode.
bA mix of shorter BMT intake types.
cUp to 1 year before first FRI episode (those reporting vaccination >1 year ago are considered unvaccinated). Participants can receive either or both vaccine formulations.
Four hundred twenty-three participants (7.6%) had ≥2 FRI episodes. The median time between consecutive episodes was 34 days (IQR, 21–55 days). While 12.1% in the 19-week BMT intake type had ≥2 FRI episodes, this was unsurprisingly less common for the shorter 17-week (8.5%; odds ratio [OR], 0.68 [95% confidence interval {CI}, .55–.84]), 9-week (1.9%; OR, 0.14 [95% CI, .09–.22]), and other (1.9%; OR, 0.14 [95% CI, .07–.30]) intake types. Other factors associated with having ≥2 FRI episodes were Malay and Indian ethnicity (versus Chinese) and being ex- or current smokers (versus nonsmokers). Compared with participants who not recently vaccinated, among whom 10.4% had ≥2 FRI episodes, 7.4% (OR, 0.69; 95% CI, .50–.95), 6.7% (OR, 0.62; 95% CI, .48–.79), and 6.4% (OR, 0.59; 95% CI, .18–1.92) among those who recently received MIV only, TIV only, or both MIV and TIV, respectively.
Distribution of Viruses in Initial and Subsequent FRI Episodes
A total of 3487 FRI episodes (56.8%) were positive for ≥1 virus included in the panel (Table 2). ERV was the most common, in 1449 (23.6%) FRI episodes, followed by FluV (in 870 [14.2%], including 138, 137, and 568 positive for A(H1N1)pdm09, influenza A(H3N2) virus, and influenza B virus, respectively), AdV (in 677 [11.0%], including 121 and 505 positive for AdV-B and AdV-E, respectively), and CoV (in 458 [7.5%], with the majority [282] positive for CoV-OC43). hMPV, hPIV, and RSV were less frequently observed (in 181, 254, and 30 FRI episodes, respectively). Three hundred ninety-four episodes (6.4%) were simultaneously positive for agents from ≥2 virus groups.
Table 2.
Distribution of Viruses in Initial Febrile Respiratory Illness (FRI) Episodes and Number and Proportion with Subsequent Episodes
| Viral Etiology of Subsequent Episodes, No. (%) of Initial Episodes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Viral Etiology of Initial Episode | Initial Episodes, No. | Subsequent Episodes, No. (%) of Initial Episodes | ≥1 Panel Virus | AdV | CoV | ERV | hMPV | FluV | hPIV | RSV |
| Overall | 6138 | 461 (7.5) | 230 (3.7) | 61 (1.0) | 50 (0.8) | 84 (1.4) | 16 (0.3) | 34 (0.6) | 18 (0.3) | 3 (0.0) |
| No panel virus detected | 2651 | 245 (9.2) | 106 (4.0) | 35 (1.3) | 17 (0.6) | 38 (1.4) | 10 (0.4) | 16 (0.6) | 5 (0.2) | 1 (0.0) |
| ≥1 panel virus | 3487 | 216 (6.2) | 124 (3.6) | 26 (0.7) | 33 (0.9) | 46 (1.3) | 6 (0.2) | 18 (0.5) | 13 (0.4) | 2 (0.1) |
| AdV | 677 | 13 (1.9) | 6 (0.9) | 2 (0.3)a | 1 (0.1) | 3 (0.4) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| CoV | 458 | 44 (9.6) | 29 (6.3) | 5 (1.1) | 7 (1.5)a | 11 (2.4) | 3 (0.7) | 2 (0.4) | 4 (0.9) | 0 (0.0) |
| ERV | 1449 | 99 (6.8) | 58 (4.0) | 15 (1.0) | 16 (1.1) | 24 (1.7)a | 3 (0.2) | 8 (0.6) | 4 (0.3) | 2 (0.1) |
| hMPV | 181 | 13 (7.2) | 8 (4.4) | 1 (0.6) | 2 (1.1) | 4 (2.2) | 0 (0.0)a | 1 (0.6) | 1 (0.6) | 0 (0.0) |
| FluV | 870 | 46 (5.3) | 24 (2.8) | 4 (0.5) | 8 (0.9) | 8 (0.9) | 0 (0.0) | 5 (0.6)a | 3 (0.3) | 0 (0.0) |
| hPIV | 254 | 16 (6.3) | 9 (3.5) | 1 (0.4) | 2 (0.8) | 3 (1.2) | 0 (0.0) | 3 (1.2) | 2 (0.8)a | 0 (0.0) |
| RSV | 30 | 3 (10.0) | 2 (6.7) | 0 (0.0) | 0 (0.0) | 1 (3.3) | 0 (0.0) | 1 (3.3) | 0 (0.0) | 0 (0.0)a |
Abbreviations: AdV, adenovirus; CoV, coronavirus; ERV, enterovirus/rhinovirus; FluV, influenza virus; hMPV, human metapneumovirus; hPIV, human parainfluenza virus; RSV, respiratory syncytial virus.
aThe initial and subsequent episode involve the same virus group.
FRI episodes with an identified virus were less likely than those without an identified virus to be followed by a subsequent FRI episode (6.2% vs 9.2%, respectively; P < .001), and subsequent FRI episodes with viruses from the same group were rare (<2% across all virus groups).
Temporal Distribution of FRI Cases, by Calendar Month and BMT Phase
From December 2009 through December 2014, there was an average of 100 FRI cases per month, but the number of cases fluctuated widely. While AdV infections were concentrated between June 2011 and February 2013 (Figure 2A), FluV infections mostly occurred before November 2010, with a sharp decline following routine administration of TIV. However, considerable influenza A(H3N2) virus and influenza B virus activity was observed from April 2013 through October 2014 (Figure 2B). When analyzed by time from enlistment, ERV infections were concentrated in earlier BMT phases, while CoV and AdV infections circulated mostly during weeks 3–8 and weeks 5–12 after enlistment, respectively (Figure 2C). FluV circulation had 2 peaks, at weeks 1–2 and then again at weeks 5–8 after enlistment (Figure 2D); the former pattern was observed after and the latter before routine TIV administration (data not shown).
Figure 2.

A and C, Distribution of febrile respiratory illness (FRI) episodes overall and those due to enterovirus/rhinovirus, adenovirus, and coronavirus, by calendar time (A) and week after basis military training (BMT) enlistment (C). B and D, Distribution of FRI episodes due to any influenza virus, 2009 pandemic influenza A(H1N1) virus (A[H1N1]pdm09), influenza A(H3N2) virus, and influenza B virus, by calendar time (B) and week after BMT enlistment (D). Gray shading denotes the period when trivalent influenza vaccine was routinely administered to incoming recruits.
Prior Infection With AdV or FluV Tends to Protect Against Subsequent FRI episode
Among FRI episodes with an identified virus, the proportion that were not followed by a FRI episode decreased more gradually than that among FRI episodes without an identified virus (Figure 3A). Relative protection against a subsequent FRI episode was also observed for AdV- positive versus AdV-negative episodes (Figure 3B) and likewise for FluV infections (Figure 3C); all 3 results were statistically significant at P < .001. However, no significant relationships were observed for CoV, ERV, hMPV, hPIV, and RSV (Supplementary Figure 1).
Figure 3.

Kaplan-Meier plots of the risk of a subsequent febrile respiratory illness (FRI) episode over time, by virological status of the previous FRI episode. A, Exposure status for any panel virus. B, Exposure status for adenovirus. C, Exposure status for influenza virus. P values were calculated by log-rank tests to compare observed and expected events among participants who tested positive or negative for the respective virus group.
Model 1, which accounted for the hierarchical data structure and other covariates, gave similar results (Table 3). Relative to episodes without an identified virus, only AdV (hazard ratio [HR], 0.24; 95% CI, .14–.44) and FluV (HR, 0.52; 95% CI, .38–.73) infections conferred significant protection against subsequent FRI episodes. AdV and FluV infections remained significantly protective, after adjustment for FRI incidence by time from enlistment (model 2) and after addition of a variable that stratified subintervals on whether they ended <4 weeks before or ≥4 weeks after the initial FRI episode (model 3). Interestingly, the hazard of a subsequent FRI episode was significantly lower in subintervals ending <4 weeks after the initial episode (vs those ending ≥4 weeks after the initial episode; HR, 0.43; 95% CI, .30–.61). Moreover, there was evidence for interaction with FluV (Pinteraction = .085); stratified HRs for FluV infection in subintervals ending <4 weeks and ≥4 weeks after the initial FRI episode were 0.23 (95% CI, .08–.63; P = .005) and 0.58 (95% CI, .38–.87, P = .009), respectively.
Table 3.
Crude Incidence Rate and Results of Survival Analyses of Associations Between Prior Infection and Protection Against Subsequent Febrile Respiratory Illness (FRI) Among Major Basic Military Training (BMT) Intake Types
| Model 1b,c | Model 2b,d | Model 3b,e | ||||
|---|---|---|---|---|---|---|
| Variablea | HR (95% CI) | P | HR (95% CI) | P | HR (95% CI) | P |
| Virus in prior episode (vs no virus identified) | ||||||
| AdV | .24 (.14–.44) | <.001 | .27 (.15–.47) | <.001 | .24 (.12–.47) | <.001 |
| CoV | 1.07 (.77–1.49) | .685 | 1.12 (.81–1.54) | .501 | 1.35 (.91–2.02) | .136 |
| ERV | .91 (.71–1.15) | .423 | .90 (.71–1.14) | .397 | .89 (.66–1.18) | .410 |
| hMPV | .91 (.52–1.59) | .734 | .97 (.57–1.67) | .915 | 1.01 (.51–2.00) | .983 |
| FluV | .52 (.38–.73) | <.001 | .54 (.38–.74) | <.001 | .50 (.34–.74) | .001 |
| hPIV | .98 (.58–1.64) | .930 | 1.02 (.62–1.69) | .938 | 1.09 (.59–2.00) | .780 |
| RSV | 1.80 (.52–6.22) | .351 | 1.94 (.58–6.43) | .281 | 2.96 (.71–12.31) | .136 |
| FRI incidence rate by time from enlistment (per 1000 person-days) | Omitted | 1.44 (1.16–1.78) | .001 | 1.87 (1.41–2.49) | <.001 | |
| Subinterval ending <4 wk after prior episode (vs ≥4 wks) | … | Omitted | .43 (.30–.61) | <.001 | ||
| Type of BMT intake (vs 19 wk) | ||||||
| 17 wk | .83 (.63–1.09) | .181 | .83 (.64–1.09) | .175 | .81 (.59–1.12) | .205 |
| 9 wk | .43 (.27–.70) | .001 | .42 (.26–.68) | <.001 | .46 (.26–.81) | .007 |
| Otherf | 1.31 (.57–3.01) | .524 | Omitted | Omitted | ||
Abbreviations: AdV, adenovirus; CI, confidence interval; CoV, coronavirus; ERV, enterovirus/rhinovirus; FluV, influenza virus; HR, hazard ratio; hMPV, human metapneumovirus; hPIV, human parainfluenza virus; RSV, respiratory syncytial virus.
aOnly variables with results significant at P < .05 are shown in the table.
bAll models were adjusted for study year, type of BMT intake, age, ethnicity, smoking history, history of asthma, and influenza vaccination.
cNot adjusted for FRI incidence rate by time from enlistment or for timing of interval relative to the prior consultation episode. All episodes are included as exposures and events of interest. All BMT types are included.
dAdjusted for FRI incidence rate by time from enlistment, expressed as FRI episodes per 1000 enlistee days (as defined and illustrated in methods and Figure 1B), but not for timing of interval relative to the prior consultation episode. All episodes are included as exposures and events of interest. Only the 19-week, 17-week, and 9-week BMT types are included.
eAdjusted for the FRI incidence rate by the time since enlistment, expressed as the number of FRI episodes per 1000 enlistee-days (as defined and illustrated in Methods and Figure 1B), and for whether the interval assessed was <4 weeks or ≥4 weeks after the prior episode. The initial episode is the exposure of interest, and the second episode is the event of interest. Only the 19-week, 17-week, and 9-week BMT types are included.
fA mix of shorter BMT intake types.
Supplementary Table 1 corroborates these findings through an alternative approach using crude FRI incidence rates based on the underlying BMT population at risk. Following AdV and FluV infection, incidence rates for subsequent FRI episodes were consistently lower than the average for all recruits in the 19-, 17-, and 9-week BMT intake types. Moreover, compared with the full course duration, analyses censored 8 and 6 weeks after enlistment showed stronger protective effects from prior FluV infection. Likewise, incidence rate ratios stratified by BMT intake type revealed increased protection by prior FluV infection with shorter course durations (Table S2).
Prior Infection With AdV or FluV Tends to Protect Against Subsequent Infection With Specific Viruses
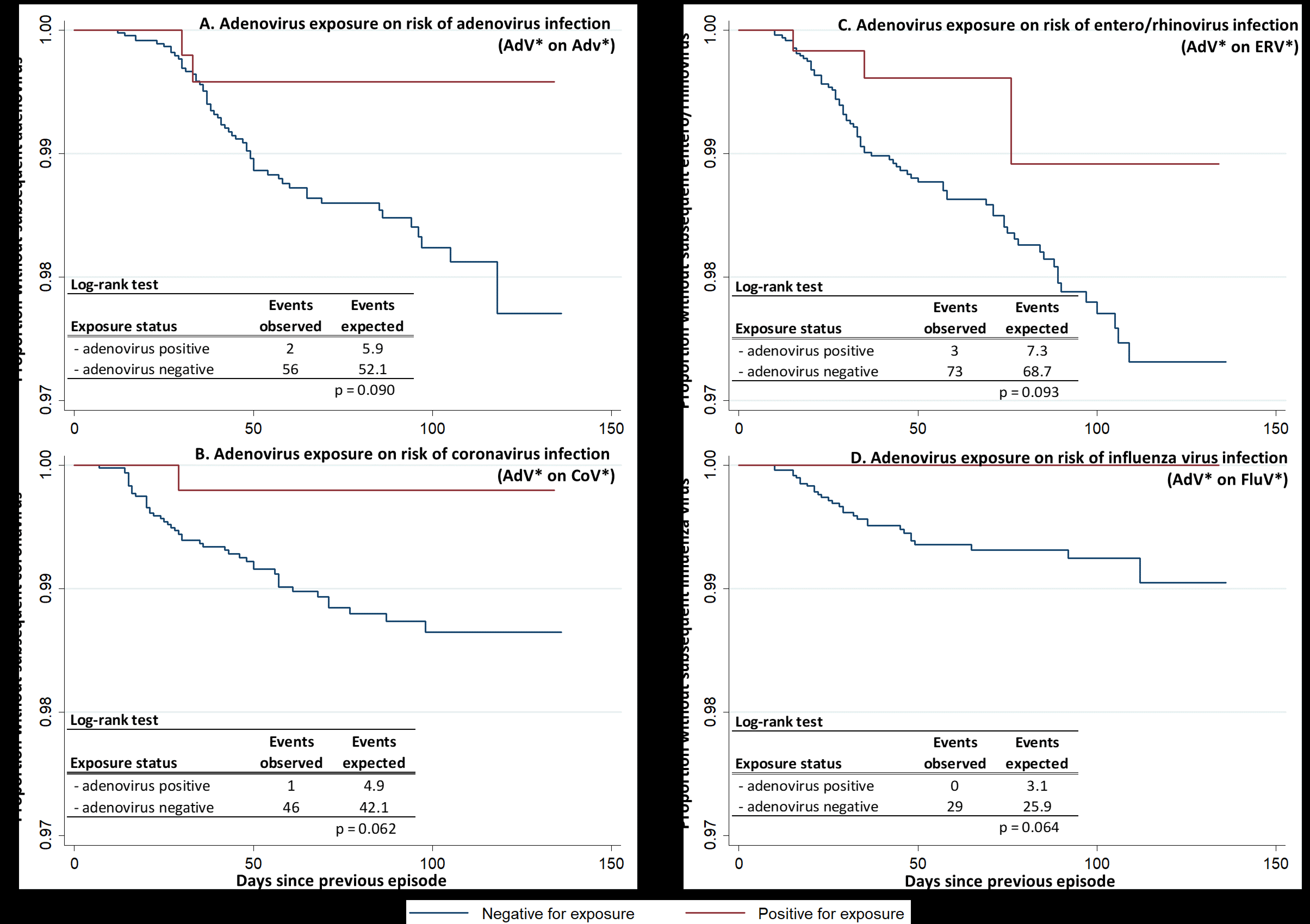
AdV-positive episodes tended to protect against subsequent infection with AdV (P = .090) and also 3 other virus groups commonly encountered in BMT, namely CoV, ERV, and FluV (P = .062, .093, and .064 respectively; Supplementary Figure 2). FluV–positive FRI episodes significantly reduced the risk of subsequent AdV infection (P = .044; Supplementary Figure 3A). Cross-protection between FluV episodes was not apparent in the full data set (Supplementary Figure 3B). However, before routine administration of TIV (ie, on or before 4 October 2010), FluV–positive episodes tended to protect against subsequent FluV infection in enlistees who had not recently received TIV (P = .081; Supplementary Figure 3C). None of the other virus groups conferred significant group-specific protection (Supplementary Table 3).
DISCUSSION
Our study provides evidence on how having a FRI episode due to respiratory viruses from certain categories reduced the risk of a subsequent FRI episode during basic military training relative to having a FRI episode that was not due to those respiratory viruses. Among the viruses, protective effects were strongest following AdV and FluV episodes. AdV infections conferred protection against subsequent infection with AdV and cross-protection to other groups of commonly circulating viruses in BMT (CoV, ERV and FluV), while FluV infection conferred significant protection against subsequent AdV infection. The observation of influenza’s protection against subsequent episodes of influenza (mostly due to other types/subtypes in our data, in the period prior to routine TIV administration) is also consistent with reports by other human and animal model studies [6, 20, 21].
Previous time-series analyses have demonstrated viral interference after accounting for seasonal factors, suggesting that epidemics of influenza virus and rhinovirus infection tend to shift the timing of epidemics for other viruses [8]. Others have used case-control–type analyses of codetection data to demonstrate that rhinovirus infection, as an exposure, was inversely associated with the probability of observing adenovirus [12] and influenza virus infections, with the latter 2 viral infections framed as the outcomes of interest [12, 13]. However, in a study of military recruits in which adenovirus was the dominant FRI agent, Wang et al used a case-control analysis to demonstrate the reverse phenomenon—that adenovirus infection as an exposure reduced the odds of rhinovirus infection as the outcome of interest [11]. However, they noted how another interpretation of their results is that rhinovirus infection reduced the risk of subsequent adenovirus infection. The opposing framing of outcomes and exposures in these codetection studies highlights issues in interpreting results from such cross-sectional and case-control designs, in which the sequence of infections cannot be ascertained. What distinguishes this work is our use of a cohort study design by which we can identify the temporality and sequence of initial and subsequent FRI episodes, which the previous studies could not do. Observations positive for a specific virus group are essentially being compared against those negative for all viruses we tested for. Therefore, this analyses does not assess whether, for example, infections by the ERV group confers any protection relative to individuals who did not recently have any viral infection. Supplementary Tables 1 and 2 suggest that, relative to enlisted recruits for the main BMT intake types, incidence rates for subsequent FRI episodes were higher over the full course duration and for 19-week BMT intake types, respectively. This applied to all virus groups except AdV and FluV groups and was possibly due to confounding by common risk factors for repeated FRIs at the individual level [15]. However, in the shortest, 9-week BMT type and when censoring the data at shorter periods after enlistment, incidence rates were reduced for most virus groups relative to the average rates for recruits following enlistment. For FRI episodes as a whole, incidence rate ratios were 0.49 (95% CI, .29–.77; P < .001) in the data censored 4 weeks after enlistment (Supplementary Table 1). These observations therefore suggest that viral causes of FRI other than AdV and FluV may also confer short, time-limited protection. They also corroborate findings from Table 3 on how a reduced hazard of a subsequent FRI episode occurs in the first 4 weeks following the initial FRI episode, relative to latter periods, and how FluV is more strongly protective during the former period. The difference between the viruses may thus be one of degree, with most viruses conferring some weak, time-limited protection, with the stronger effects from adenovirus and influenza virus becoming apparent when aggregating data over the full course duration. One mechanism consistent with such short-lived cross-protection is the triggering of interferon-stimulated genes and cytokines, causing nearby cells in the respiratory tract to enter an antiviral state [22, 23]. Alternatively, transient cross-protection could result from infection-induced behavioral changes, such as improved hand hygiene following an FRI and a short duration of reduced exposure to new pathogens while being furloughed on home leave. However, this on its own is likely inadequate to explain the extent of cross-protection observed with adenovirus and influenza virus infections relative to other FRI episodes. Other proposed mechanisms, such as competition for the same receptor-binding sites and intracellular host machinery for replication [24], and an unfavorable physiological state of the host, such as high body temperatures upon an initial infection [11], are more applicable to protection against coinfections but not, as demonstrated here, against sequential infection.
Our findings have several implications. They provide an explanation for what has previously been suspected on the basis of time-series analyses, such as how influenza epidemics potentially delay RSV epidemics [8, 25]. They also add to the body of evidence motivating the search for the causative mechanisms for cross-protection among viral infections while providing additional clues. Given the relative ranking of virus families in conferring cross-protection, we suggest focusing the search for mechanisms on common factors induced by both adenovirus and influenza virus. Notably, our previous studies suggest that infections with these 2 infections may be more severe, because they are more likely to present with febrile illness [26] and higher temperatures (≥38°C) than infections due to other common circulating virus families [16]. This again supports a cytokine-mediated antiviral effect. Finding a means to induce such cross-protection without severe side effects could potentially lead to interventions that may be particularly useful in the context of exposure to dangerous viruses for which there is no specific vaccine or antiviral agent [27].
However, our study has some limitations. Our study population largely comprises healthy young adults, and key circulating viruses in this group differs somewhat from those observed in the general population in Singapore [28]. Also, we excluded respiratory illness episodes that did not meet our inclusion criteria of having a temperature ≥37.5°C. We also acknowledge that our capture of data on FRI episodes is likely incomplete, as consultations outside office hours and over the weekend (when trainees are on home leave) were missed by our surveillance program. However, we argue that restricting our study to febrile cases and the incomplete capture of data are biases toward the null and thus do not negate our main findings on the relative cross-protection against subsequent FRI episodes conferred by adenovirus and influenza virus infections.
In summary, our study demonstrated broad-based viral interference in a population of military recruits. Infections from the adenovirus and influenza virus families conferred significant cross-protective effects against subsequent FRI episodes relative to other circulating viruses. The duration of these cross-protective effects extend beyond those previously demonstrated in studies of viral coinfections. Our study points the way for research into the underlying mechanisms for broad-based antiviral activity in the human host, which deserves greater study.
Supplementary Data
Supplementary materials are available at The Journal of Infectious Diseases online. Consisting of data provided by the authors to benefit the reader, the posted materials are not copyedited and are the sole responsibility of the authors, so questions or comments should be addressed to the corresponding author.
{kind=link}
{kind=link}
{kind=link}
Notes
Acknowledgments. We thank our colleagues whose work on the project, along with our own, made this article possible; and Benjamin Tay, for assisting with extraction of additional data during manuscript revision.
Financial support. This work was supported by the Ministry of Education (tier 1 grant R-608-000-132-112 to C. X. P. C.) and by the Centre for Infectious Disease Epidemiology and Research, Saw Swee Hock School of Public Health (funded by the Singapore Ministry of Defence).
Potential conflicts of interest. All authors: No reported conflicts of interest. All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
References
- 1. McKinney HH. Mosaic diseases in the Canary Islands, West Africa and Gibraltar. J Agric Res 1929; 39:577–8. [Google Scholar]
- 2. Henle W. Interference phenomena between animal viruses; a review. J Immunol 1950; 64:203–36. [PubMed] [Google Scholar]
- 3. Isaacs A, Lindenmann J. Classics in oncology: virus interference: I. the interferon. CA Cancer J Clin 1988; 38:280–90. [PubMed] [Google Scholar]
- 4. Chan KF, Carolan LA, Korenkov D, et al. . Investigating viral interference between influenza a virus and human respiratory syncytial virus in a ferret model of infection. J Infect Dis 2018; 218:406–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Costa-Hurtado M, Afonso CL, Miller PJ, et al. . Virus interference between H7N2 low pathogenic avian influenza virus and lentogenic Newcastle disease virus in experimental co-infections in chickens and turkeys. Vet Res 2014; 45:1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Laurie KL, Guarnaccia TA, Carolan LA, et al. . Interval between infections and viral hierarchy are determinants of viral interference following influenza virus infection in a ferret model. J Infect Dis 2015; 212:1701–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Yang L, Chan KH, Suen LKP, et al. . Impact of the 2009 H1N1 pandemic on age-specific epidemic curves of other respiratory viruses: a comparison of pre-pandemic, pandemic and post-pandemic periods in a subtropical city. PLoS One 2015; 10:e0125447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Asten L van, Bijkerk P, Fanoy E, et al. . Early occurrence of influenza A epidemics coincided with changes in occurrence of other respiratory virus infections. Influenza Other Respir Viruses 2016; 10:14–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Zheng X, Song Z, Li Y, Zhang J, Wang X-L. Possible interference between seasonal epidemics of influenza and other respiratory viruses in Hong Kong, 2014–2017. BMC Infect Dis 2017; 17:772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Linde A, Rotzén-Ostlund M, Zweygberg-Wirgart B, Rubinova S, Brytting M. Does viral interference affect spread of influenza? Euro Surveill Bull 2009; 14(40). [PubMed] [Google Scholar]
- 11. Wang Z, Malanoski AP, Lin B, et al. . Broad spectrum respiratory pathogen analysis of throat swabs from military recruits reveals interference between rhinoviruses and adenoviruses. Microb Ecol 2010; 59:623–34. [DOI] [PubMed] [Google Scholar]
- 12. Greer RM, McErlean P, Arden KE, et al. . Do rhinoviruses reduce the probability of viral co-detection during acute respiratory tract infections? J Clin Virol 2009; 45:10–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Casalegno JS, Ottmann M, Duchamp MB, et al. . Rhinoviruses delayed the circulation of the pandemic influenza A (H1N1) 2009 virus in France. Clin Microbiol Infect 2010; 16:326–9. [DOI] [PubMed] [Google Scholar]
- 14. Ho HP, Zhao X, Pang J, et al. . Effectiveness of seasonal influenza vaccinations against laboratory-confirmed influenza-associated infections among Singapore military personnel in 2010–2013. Influenza Other Respir Viruses 2014; 8:557–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Pang J, Jin J, Loh JP, et al. . Risk factors for febrile respiratory illness and mono-viral infections in a semi-closed military environment: a case-control study. BMC Infect Dis 2015; 15:288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Tan XQ, Zhao X, Lee VJ, et al. . Respiratory viral pathogens among Singapore military servicemen 2009–2012: epidemiology and clinical characteristics. BMC Infect Dis 2014; 14:204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Hsu JP, Zhao X, Chen MI-C, et al. . Rate of decline of antibody titers to pandemic influenza A (H1N1-2009) by hemagglutination inhibition and virus microneutralization assays in a cohort of seroconverting adults in Singapore. BMC Infect Dis 2014; 14:414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Young B, Sadarangani S, Jiang L, Wilder-Smith A, Chen MI. Duration of influenza vaccine effectiveness: a systematic review, meta-analysis, and meta-regression of test-negative design case-control studies. J Infect Dis 2018; 217:731–41. [DOI] [PubMed] [Google Scholar]
- 19. Young B, Zhao X, Cook AR, Parry CM, Wilder-Smith A, I-Cheng MC. Do antibody responses to the influenza vaccine persist year-round in the elderly? A systematic review and meta-analysis. Vaccine 2017; 35:212–21. [DOI] [PubMed] [Google Scholar]
- 20. Goh EH, Jiang L, Hsu JP, et al. . Epidemiology and relative severity of influenza subtypes in Singapore in the post-pandemic period from 2009 to 2010. Clin Infect Dis 2017; 65:1905–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Schultz-Cherry S. Viral interference: the case of influenza viruses. J Infect Dis 2015; 212:1690–1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Takeda K, Akira S. Toll-like receptors. Curr Protoc Immunol 2007; Chapter 14:Unit 14.12. [DOI] [PubMed] [Google Scholar]
- 23. Khaitov MR, Laza-Stanca V, Edwards MR, et al. . Respiratory virus induction of alpha-, beta- and lambda-interferons in bronchial epithelial cells and peripheral blood mononuclear cells. Allergy 2009; 64:375–86. [DOI] [PubMed] [Google Scholar]
- 24. Pinky L, Dobrovolny HM. Coinfections of the respiratory tract: viral competition for resources. PLoS One 2016; 11:e0155589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Anestad G. Interference between outbreaks of respiratory syncytial virus and influenza virus infection. Lancet 1982; 1:502. [DOI] [PubMed] [Google Scholar]
- 26. Lau Y-F, Koh W-HV, Kan C, et al. . Epidemiologic analysis of respiratory viral infections among Singapore military servicemen in 2016. BMC Infect Dis 2018; 18:123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Beilharz MW, Cummins MJ, Bennett AL, Cummins JM. Oromucosal administration of interferon to humans. Pharmaceuticals (Basel) 2010; 3:323–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Jiang L, Lee VJ, Cui L, et al. . Detection of viral respiratory pathogens in mild and severe acute respiratory infections in Singapore. Sci Rep 2017; 7:42963. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.