Figure 4. NPC2 binding to symbiont-produced sterols via immunoprecipitation-lipidomics, and differential stability of non-canonical NPC2 at varying pH.
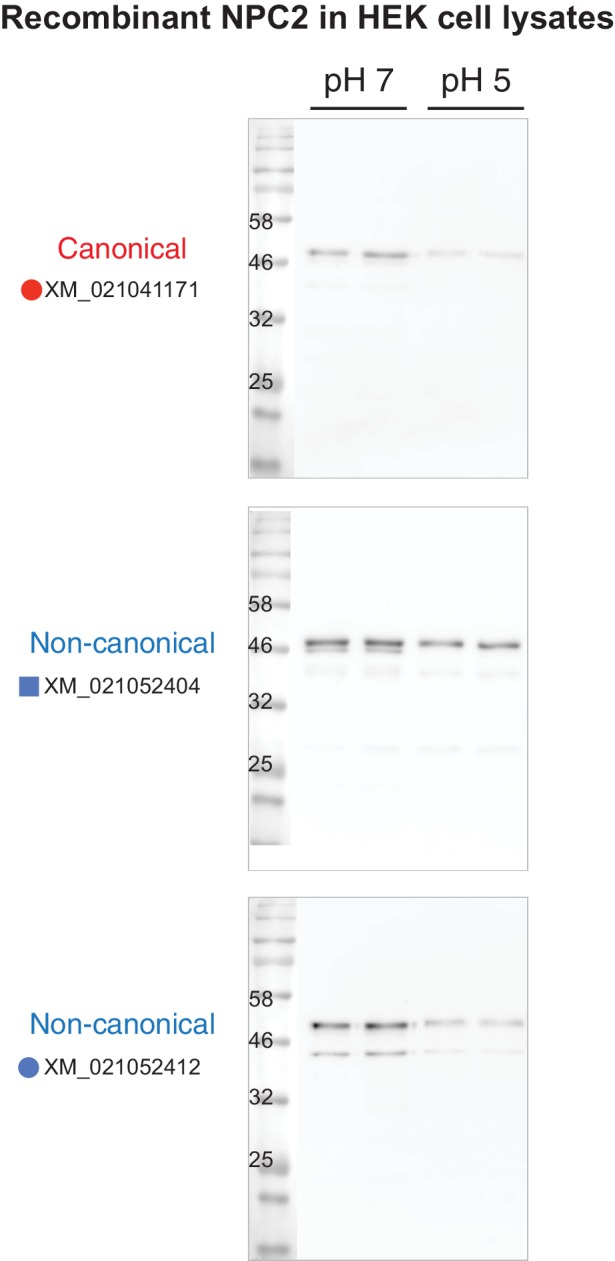
(A) Quantification of bound lipids in the eluates following IP of recombinant canonical and non-canonical NPC2:crmCherry:HA and negative control crmCherry:HA at pH 7 (Figure 4—figure supplement 1). Sterols comprising <1.5% were omitted for clarity. crmCherry, lysosome-stable cleavage-resistant mCherry (Huang et al., 2014) . Average values ± SD (error bars). Except ß-sitosterol, each NPC2 was significantly different to crmCherry negative control (Student’s t-test, p<0.01). (B) Relative proportions of NPC2-bound sterols and the corresponding symbiont extract at pH 5 and 7. Average values ± SD (error bars) (C) Immunoprecipitation (IP) of native non-canonical NPC2 from Aiptasia and quantification of eluted bound sterols. Control, identical reaction omitting antibody. Average values ± SEM (error bars). Statistical comparisons to control (Student’s t-test: *p<0.05, •p<0.09). (D) NPC2 gene expression by qPCR in the various Aiptasia/Symbiodiniaceae host/symbiont combinations in Figure 1B. (E) Recombinant NPC2s detected by mCherry antibody in the soluble fractions of HEK cell lysates. Lysate preparations were identical except for buffer pH; equivalent volumes loaded per lane. (F) Quantification of protein abundances from Western blots in E. Average values ± SEM (error bars). Student’s t-test, *p<0.05. (G) We propose a model in which symbiotic anthozoans have evolved non-canonical NPC2 homologues that are spatiotemporally regulated to specifically respond to symbiosis, including through adaptation to the acidic environment of the symbiosome, the lysosomal-like organelle in which symbionts reside. NPC2 proteins bind and transport symbiont-produced sterols, and such trafficking is essential for cellular homeostasis of the sterol-auxotrophic hosts.

Figure 4—figure supplement 1. Immunoprecipitation-lipidomics: protein inputs and immunoprecipitation (IP) elutions.

Figure 4—figure supplement 2. Soluble recombinant NPC2s in HEK 293T cell lysates.