

Abstract
Successful treatment of cancer remains a challenge, due to the unique pathophysiology of solid tumors, and the predictable emergence of resistance. Traditional methods for cancer therapy including radiotherapy, chemotherapy, and immunotherapy all have their own limitations. A novel approach is bacteriotherapy, either used alone, or in combination with conventional methods, has shown a positive effect on regression of tumors and inhibition of metastasis. Bacteria‐assisted tumor‐targeted therapy used as therapeutic/gene/drug delivery vehicles has great promise in the treatment of tumors. The use of bacteria only, or in combination with conventional methods was found to be effective in some experimental models of cancer (tumor regression and increased survival rate). In this article, we reviewed the major advantages, challenges, and prospective directions for combinations of bacteria with conventional methods for tumor therapy.
Keywords: bacteriotherapy, cancer, chemotherapy, immunotherapy, radiotherapy, tumoricidal agents
1. INTRODUCTION
Cancer remains one of the most common causes of death throughout the world.1, 2, 3 In 2018, the new cases and deaths of cancer were reported 18.1 and 9.6 million, respectively.4 By 2030, it is expected that there will be ~17 million deaths.1 These statistics emphasize the urgency of finding novel and more effective treatments. The historical treatment options for cancer, including surgery, radiotherapy, chemotherapy, and immunotherapy all have some limitations. Treatment of various cancers is difficult due to the multifactorial physiology (including problems with volume, site, stage, and metastasis of the tumor). Moreover, resistance often emerges to reduce the initial effectiveness of chemotherapy, radiotherapy, and immunotherapy, leading to poor tumor control, and many side effects occur during or after the treatment).5 Alternative or complementary therapies such as gene therapy, diet therapy, photodynamic therapy, insulin potentiating therapy, HAMLET (human alpha‐lactalbumin made lethal to tumor cells), telomerase therapy, hyperthermia therapy, dichloroacetate, non‐invasive RF cancer treatment, and bacteriotherapy have been proposed to improve and increase the effectiveness of conventional cancer therapy.6
The use of therapeutic bacteria is one approach that may be able to overcome some of the limitations of conventional cancer therapy as stated above. Bacteria alone can act as potent antitumor agents. Another remarkable feature of bacteria is their ability to be genetically engineered to alter their ability to synthesize and release specific compounds, and tailor their metabolic pathways. Publications on the use of bacterial anti‐cancer therapies have grown significantly over the past few decades. Therapeutic bacteria can especially target the hypoxic areas of tumors and actively penetrate the tissue, and can allow different strategies such as the secretion of toxins/enzymes including proteases and lipases to be tested. Bacteria can be used as vectors to carry tumoricidal agents and immunotherapeutic agents, thereby destroying tumor cells (Table 1). 7, 8, 9, 10 However, the fight against cancer is not expected to be won any time soon, and so creative efforts to harness the power of bacteria for cancer treatment will still continue. This review covers the use of bacteria as anticancer agents to improve cancer treatment.
Table 1.
The treatment strategy of bacteria in cancer therapy
| Treatment strategy | Type of bacteria, treatment approach | Outcome | Ref. |
|---|---|---|---|
| Immunotherapeutic agents | Streptococcus pyogenes, intentionally infected a cancer patient with erysipelas | Rapid tumor regression | 149 |
| Streptococcus pyogenes, intentionally infected a cancer patient with erysipelas | Regression of cancer | 150 | |
| Bacillus Calmette‐Guerin, injection into patients with tuberculosis | Reduced frequency of cancer | 151 | |
| Clostridium spp, concurrently suffered from gas gangrene in patients with tumor | Tumor regression | 152 | |
| Attenuated Salmonella typhimurium, vaccination of the B16F10 tumor‐bearing mice by derivatives Salmonella typhimurium (SL1344 InvA or SL3261AT InvA | An antitumor effect | 51 | |
| Listeria monocytogenes, vaccination a recombinant Listeria monocytogenes (Lm‐NP) on breast, melanoma, and cervical cancer | Regression growth all types of tumors | 153 | |
| Vectors/spores to carry tumoricidal agents | Clostridium acetobutylicum DSM792, cloned of the construction (pIMP1eglArIL2) of the rIL2 expression/secretion vector into Clostridium acetobutylicum DSM792 | A significant increase in secretory production of biologically active rat interleukin‐2 | 154 |
| Clostridium novyi‐NT, IV injection of C novyi‐NT spores and a single IV dose of liposomal doxorubicin (Doxil) administered into mice bearing colorectal cancer | Elimination of tumors | 126 | |
| C novyi‐NT and C. sporogenes, conjugation of pMTL‐555‐VHH construct of a VHH‐AG2 expressing vector (an anti HIF‐1α) into these bacteria | Rise of delivery of therapeutics agents | 44 | |
| C novyi‐NT, IV administration of HTI‐286, docetaxel, vinorelbime, and MAC‐321 in combination with or without C novyi‐NT spores into mice bearing HTC 116 xenografts | Hemorrhagic necrosis of tumors | 155 | |
| C novyi‐NT spores, IV injection C novyi‐NT spores into CT26 and RENCA tumors in mice and VX2 tumor in rabbits | Relatively treated in mice and rabbits with cancer | 156 | |
| Bifidobacterium longum 105‐A and 108‐A, IV injection of the pBLES100 (constructed by cloning a B longum plasmid and a gene encoding spectinomycin adenyltransferase AAD from Enterococcus faecalis into the E coli vector pBR322) to B16‐F10 melanoma tumor‐bearing mice | Increase in specific gene delivery vectors in the tumor | 74 | |
| Bacterial toxins/enzymes | Salmonella enterica Serovar Typhimurium, oral administrated construction of Salmonella‐based survivin vaccine into BALB/c, colon, DBT, and GL261 glioblastoma ‐bearing mice | Vaccine as an adjuvant against different types of cancer | 157 |
| Streptococci and Serratia marcescens, injection of bacterial concoction derived from heat‐killed streptococcal and Serratia marcescens (Coley's Toxin) into body, sarcomas | A severe erysipelas infection led to the cure of cancer | 158 | |
| E coli BM2‐1 strain, direct inoculum of Cytotoxic Necrotizing Factor‐1 (CNF‐1) to the HEp‐2 cells (exposed to UVB irradiation) | Activation of the Rho GTP‐binding protein and prevent apoptosis in epithelial cells | 159 | |
| Corynebacterium diphtheriae, the incubation of the Vero cells for 1 h in growth medium with different amounts of nicked124 I‐labeled diphtheria toxin (DT) | Inhibition of protein synthesis and subsequent cell lysis and/or induction of apoptosis Vero cells | 160 | |
| Clostridium perfringens, intratumoral injections of either 2, 10 µg of Clostridium perfringens enterotoxin (CPE) in xenografts of T47D breast cancer cells in mice | Rapid and dose‐dependent cytolysis | 161 | |
| Clostridium botulinum, administration of botulinum neurotoxin (BoNTs) into tumors | BoNTs an effect on the tumor microenvironment and more effective destruction of radiotherapy and chemotherapy in cancer cells | 129 | |
| Pseudomonas aeruginosa, IV injection of the chimeric fusion protein interleukin‐4‐Pseudomonas exotoxin (IL4‐PE) into GBM induce in nude mice and, intratumor administration of IL4‐PE in malignant astrocytoma in a phase I clinical trial | A significant antitumor activity | 162 |
2. BACTERIA IN CANCER THERAPY
Cancer is a challenging disease, which requires a multi‐pronged approach for effective treatment.11 The historical role of bacteria as anticancer agents was recognized as long as one century ago (Figure 1). For the first time clinicians used live bacteria (Streptococci and Clostridia) for cancer treatment. Today, genetically modified bacteria are mostly used for this purpose.12, 13, 14, 15, 16 Bacteria can be used in cancer therapy by taking advantage of different strategies (Figure 2). These strategies include native bacterial toxicity, combination with other therapies, bacteria that can control expression of anticancer agents, expression of tumor‐specific antigens, gene transfer, RNA interference, and pro‐drug cleavage.7 The use of whole live, attenuated and/or genetically modified bacteria alone, or in combination with conventional agents has been tested in several experimental models of cancer. The most common bacteria used in this field are the genera Salmonella, Clostridium, Bifidobacterium, Lactobacillus, Escherichia, Pseudomonas, Caulobacter, Listeria, Proteus, and Streptococcus.8, 17, 18, 19, 20 The use of three species of bacteria, Clostridia, Bifidobacteria, and Salmonellae as vectors for delivering or expressing tumor suppressor genes, anti‐angiogenic genes, suicide genes, or tumor‐associated antigens has been tested in animal models bearing various tumors.8, 21, 22, 23, 24 Some clinical trials have already been conducted displaying partial responses, and thus further investigation should be performed in humans.8 Also, modified bacteria can be used for theranostic applications, since they can be detected using magnetic resonance imaging (MRI) or positron emission tomography (PET) as dual therapeutic and diagnostic agents.7, 25, 26, 27, 28, 29, 30, 31
Figure 1.
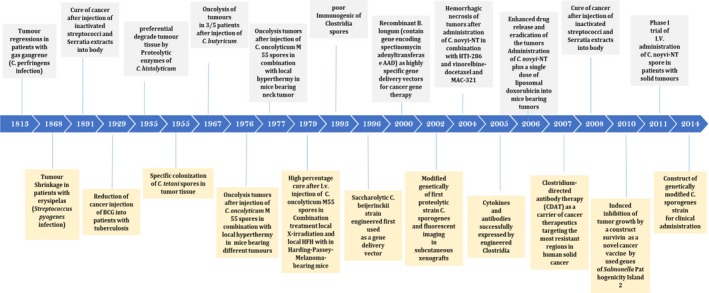
Timeline | The history uses of therapeutic bacteria in oncology
Figure 2.
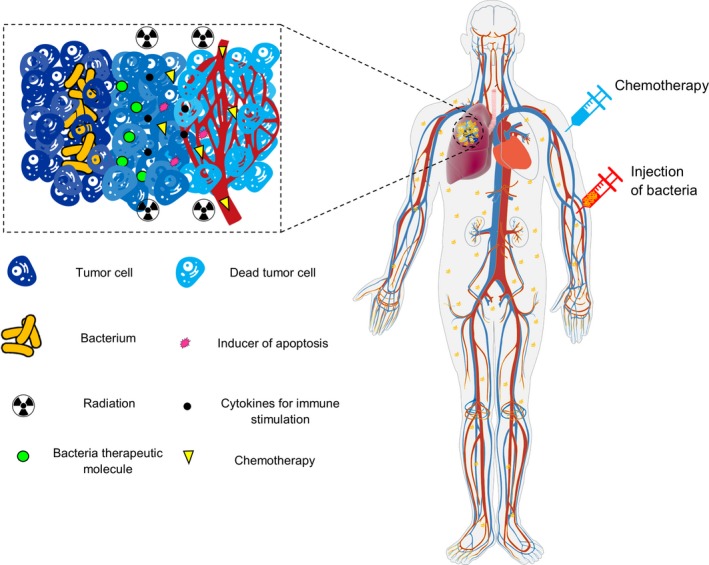
Schematic of therapeutic bacteria strategies against hypoxic tumor adapted from Forbes7
3. GENETICALLY MODIFIED BACTERIA IN CANCER THERAPY
Gene therapy is a forward‐looking alternative approach to cancer therapy. Selective targeting and destruction of tumor cells are the major advantages of gene therapy.8, 12, 32, 33 Genetically modified bacteria may also be able to lower pathogenicity to the host and increase the antitumor efficacy.32, 34 Recently, a number of studies have developed a new approach for cancer therapy using genetically engineered bacteria designed to express reporter genes, cytotoxic protein and/or anticancer agents, and tumor‐specific antigens.10, 34, 35 It has been found that genetically modified bacteria can have a more significant multiplication in tumors than in normal tissues.36, 37 The ability of Salmonella typhimurium serovar VNP20009 and Clostridium butyricum M55 to selectively colonize tumors has allowed them to be used as delivery vectors in mouse tumor models, without severe immune responses or toxic side effects. However, the results of some of these studies were less promising than expected.8, 18, 19, 38, 39, 40 Nevertheless, Clostridia strains (C acetobutylicum and C beijerinckii) can be successfully engineered to express genes encoding specific bacterial enzymes (cytosine deaminase, nitroreductase), or murine tumor necrosis factor alpha (m‐TNFα), producing more promising antitumor effects.41, 42, 43 A number of studies found that bacteria were able to produce antibodies that could bind to hypoxia inducible factor 1α (as a crucial transcription factor with a role in tumor development).7, 44, 45 Clinical trials have demonstrated that the engineered S typhimurium and Clostridium novyi ‐NT expressing HlyE or Stx2 (an acidic pH‐responsive promoter) or recA (a 38 kDa protein essential for the repair and maintenance of DNA) activated the host immune system to express cytokines such as interleukin‐2 (IL‐2), IL‐4, IL‐18, CC chemokine‐21, and consequently led to the regression and necrosis of tumors. 8, 46, 47, 48 These studies suggested that a combination of bacteriotherapy with radiotherapy, immunotherapy, or chemotherapy could be a novel and useful approach to cancer treatment.
4. BACTERIA AS IMMUNOTHERAPEUTIC AGENTS IN CANCER THERAPY
Cancer immunotherapy involves triggering a specific immune response in patients to allow various kinds of host immune cells to attack the cancer cells. It is believed that once the activated host immune cells (mainly tumor antigen‐specific CD8+ and CD4 +T lymphocytes that have been activated and stimulated) can recognize and destroy tumor cells. Bacterial infections (such as those caused by C novyi) can lead to the production of heat shock proteins such as Hsp70 which is released from necrotic cells, and pathogen‐associated molecular patterns (PAMPs) which are released from bacteria.49 Hsp70 causes maturation of dendritic cells, which are the professional antigen‐presenting cells required for the generation of effective antigen‐specific immune responses. PAMPs bind to and activate toll‐like receptors, stimulating up‐regulation of pro‐inflammatory cytokines (eg, IL‐12), and costimulatory molecules (eg, CD40). Subsequently, these mediators cause production of interferon gamma (IFN‐γ) and a Th1‐dependent cell‐mediated response will commence, essentially mediated by CD8+ effector cells.50 CD8+ lymphocytes isolated from C novyi NT‐treated mice can in turn stimulate acquired immunity in a tumor‐specific model. Avogadri et al, have described an interesting approach to melanoma immunotherapy dependent on the ability of intracellular bacteria like S typhimurium to infect host cells.51 They observed that S typhimurium used a type‐3 secretion system (T3SS) to infect tumor cells. Essentially, mutant strains, which were defective in T3SS lost the ability to enter tumor cells, both in vivo and in vitro. Tumor cells infected by Salmonella are not directly destroyed by the S typhimurium; but rather bacterial antigens are presented and become targets for anti‐Salmonella—specific T cells, a process that has not been fully explained. Furumoto et al, have reported that compounds derived from bacteria (CpG oligonucleotides) can be used to activate dendritic cells and cause complete regression of B16F10 melanoma tumors, which are known to be highly immunosuppressive in mice.52
Recombinant Escherichia coli strains have been utilized for over 30 years as a standard tool in molecular biology, and are broadly used for manufacturing recombinant proteins. These strains can also be used for the delivery of tumor antigens into dendritic cells. The simultaneous production of listeriolysin O (LLO; a pore‐forming cytolysin released from Listeria monocytogenes) together with ovalbumin (OVA; a model tumor antigen) in E coli strains led to the MHC class I presentation of the OVA Kb‐restricted epitope, SIINFEKL, after phagocytosis of the microorganisms by macrophages.53 LLO is expressed as a bacterial cytoplasmic protein, and is only released subsequent to the uptake of the L monocytogenes by phagocytosis and degradation within the phagocytic vesicles. LLO then punctures the phagosome, permitting its release into the cytosol together with co‐expressed proteins, for processing and presentation by the MHC class I pathway. A recent study showed that, if applied several times, a combination of PAMPs could eradicate solid tumors in cancer bearing mice.54
5. BACTERIAL TOXINS OR ENZYMES IN CANCER THERAPY
Several pathogenic microorganisms express and release particular protein toxins that serve to suppress the immune response of the infected host. Some of these have been tested to some extent for cancer therapy.55 Commonly, they catalyze the covalent alteration of specific proteins. In that way, they can inhibit the production or release of antibodies and cytokines. Moreover, they can inhibit macrophage migration and disrupt the barrier function of epithelial cells. Often, these toxins are powerful enzymes with high specificity against their cellular substrates, which are frequently signaling molecules. These enzymatic toxins have the capability to alter their substrates in the cytosol after bacteria enter the cells. A few toxins are able to change the function and morphology of the cells, or possibly kill the host cells. Since many of these toxins have well‐known structures, cellular receptors, molecular mechanisms, and uptake pathways, they have been utilized to analyze or influence particular signaling pathways of mammalian cells.55 Bacterial toxins are the most powerful cytotoxins produced by bacteria themselves. Cytolysin A (ClyA; also known as HlyE) is a bacterial enzyme toxin, which works by making pores in eukaryotic cell membranes and triggering caspase‐mediated cell death.10, 47 A few studies have found that treating mice with S typhimurium or E coli strains expressing the ClyA toxin inhibited tumor growth.10, 47 Three of the cytotoxins have been found to belong to the TNFα superfamily: TNF‐related apoptosis‐inducing ligand (TRAI‐l), FAS ligand (FAS‐l), and TNFΑ.7, 56 These proteins selectively cause programmed cell death via death receptor pathways, activating the apoptotic mediators, caspase 3 and caspase 8.56 Recently, in a report of “photo‐controlled bacterial metabolite therapy”, Zheng et al57 developed a biotic/abiotic hybrid system. They combined carbon nitride (C3N4) with an E coli strain that was able to produce nitric oxide (NO). In a mouse model, the C3N4 loaded bacteria were accumulated throughout the tumor, and the treatment resulted in a significant antitumor activity (~80% inhibition of tumor growth). Furthermore, cell cycle inhibitors (e.g. cytolethal distending toxins, CDTs, and the cycle inhibiting factor, Cif) blocked cell division and were proposed to compromise the immune system by impeding the clonal expansion of lymphocytes. Conversely, cell cycle stimulators, for example, the cytotoxic necrotizing factor (CNF) stimulate cell proliferation and interfere with cell differentiation.58, 59 Bacterial toxins that block or stimulate the eukaryotic cell cycle have been called “cyclomodulins”. For instance, CNF is a cell‐cycle stimulator released by some bacteria such as E coli. CNF stimulates the G1‐S cycle transition and increases replication of DNA. However, the overall number of cells does not increase, and the cells become multinucleated instead, possibly through the ability of toxin to prevent cell differentiation and trigger cell apoptosis.60 Cif is involved in enterohemorrhagic E coli and enteropathogenic E coli, while CDTs are produced by several Gram‐negative bacterial species, and Salmonella typhi and Campylobacter jejuni. The antitumor effects of bacterial toxins could be associated with reduced side effects compared to traditional antitumor therapy. These bacterial toxins could be combined with anti‐cancer drugs, or with irradiation to enhance the efficacy of cancer therapy.61
6. BACTERIAL SPORES AND VECTORS AS TUMORICIDAL AGENTS
Live bacterial vectors may be valuable tools for the development of new cancer therapies, which can be added to the collection of existing drugs.62 In one aspect of this novel methodology, bacteria are modified to deprive cancerous cells of oxygen, thereby causing tumor death.
The interest in utilizing bacteria as anticancer agents goes back to the end of the 19th century; however, with the advent of molecular biology, this methodology has also been recently revisited.63, 64 Over the past 50 years, many strains of obligate and facultative anerobic bacteria have been investigated as oncolytic agents owing to their ability to specifically multiply in oxygen‐poor (hypoxic) tissues.65, 66, 67 Early results suggested that anerobic bacteria particularly targeted solid tumors, triggering an inflammatory response inside the tumor followed by tumor regression in up to 30% of cases for instance after administration of bacterial spores.68, 69, 70 Several strains of Bifidobacteria, Salmonella sp,71 E coli,72, 73 and Clostridium 74 have been shown to selectively colonize the hypoxic areas of tumors and destroy tumor cells, thereby providing an additional specific tumor‐targeted therapy.62 Several studies have suggested that bacteria have been engineered to express a cytotoxic protein or reporter gene could be utilized in tumor treatment or tumor imaging.75 These genetically modified bacteria multiply in tumors by up to 1000‐fold higher, than in normal tissues.71 S typhimurium has been utilized as a carrier to transport various converting enzymes and antigens into tumors.76 The genus Clostridium is strictly anerobic and the vast majority of species can produce spores permitting survival but not growth in hypoxic conditions.77 Once conditions are favorable (as in wounds or spoiled meat), these spores can germinate into metabolically active bacterial cells. Furthermore, clostridia utilized in the field are generally susceptible to a wide range of antibiotics, permitting control of their replication at any time.77 Highly hypoxic tissue is usually only found in tumors and is absent in most other organs of the body. This high specificity was shown by Malmgren et al,78 who injected Clostridium spores into tumor‐bearing mice, and showed that only the mice with tumors died from the infection. In view of the fact that necrotic mainly areas exist only inside tumors and not in normal tissues, it was realized that lethal toxin‐free Clostridium novyi NT spores could be very efficient in eradicating established tumors.66, 67, 79 The systemic administration of clostridia spores (which is remarkably well tolerated) can lead to the destruction of tumor cells surrounding necrotic and hypoxic regions that are resistant to conventional therapies.80 The majority of anerobic bacteria tested up to now can form extremely resistant spores that enable them to survive even in oxygen‐rich conditions, but they cannot multiply or grow there. However, when they experience favorable conditions, for example, dead regions within tumors, the spores will germinate and the bacteria will multiply, rendering them ideal to target cancers. Spores of C novyi‐NT (a genetically modified strain without the lethal toxin) have demonstrated antitumor activity without any systemic side effects. In mice receiving an intratumoral injection of C histolyticum spores, a marked lysis of tumor tissues was observed. A similar phenomenon was found in mice injected intravenously with spores of C sporogenes. Additionally, Clostridium was not detected in normal tissues of mice receiving an intravenous injection of bacteria, but only in tumors. Pharmacological and toxicological assessment of mice injected with C novyi‐NT spores suggested that the spores were quickly cleared from the general circulation by the reticuloendothelial system. No systemic toxicity was seen in healthy mice or rabbits even after massive doses. However, in tumor‐bearing mice, toxicity appeared to be associated with the tumor size, and in this case, spores did cause bacterial infection. Bacterial spores have been additionally exploited as delivery agents for cytotoxic peptides, anticancer agents, therapeutic proteins, and as vectors for gene therapy.6, 40, 47
7. COMBINATION OF BACTERIOTHERAPY WITH DIFFERENT APPROACHES IN CANCER THERAPY
The combination of bacteriotherapy with other different types of cancer therapy has shown remarkable potential for both diagnostic and therapeutic applications. Chemotherapy, radiotherapy, and immunotherapy are accepted as the major conventional types of cancer treatment.81 However, conventional strategies can cause numerous complications in patients including induction of cancer cell resistance, systemic toxicity, and immune suppression, not to mention that they have failed to completely eradicate all the cancer cells in most cases. It is known to be challenging to effectively administer radiotherapy or chemotherapeutic agents to the hypoxic and acidic regions in tumors. Incomplete tumor targeting, inadequate tissue penetration, and limited cancer cell toxicity are limitations of most cytotoxic drugs.82 Therefore, viruses or other vectors have been used for selective tumor targeting and cancer therapy. About 30% of cancer‐related deaths are caused by failure of local tumor control, suggesting that improving local control has the potential to improve the survival of one‐third of all cancer patients. Several ways to improve local tumor control are currently under investigation. Promising strategies seem to be those that combine existing therapeutic modalities with new approaches, such as combining ionizing irradiation with gene therapy.83
8. COMBINATION OF BACTERIOTHERAPY WITH RADIOTHERAPY
Radiotherapy remains central among the most effective approaches to treat many different cancers. Even so, damage to normal tissues cannot be completely avoided, representing an important limitation for the efficacy of this cancer treatment approach.84 One of the fundamental explanations for the lack of efficacy of radiotherapy in some solid tumors is the presence of hypoxic (i.e. poorly vascularized) zones that are resistant to radiation. However, this limitation could be an advantage for other approaches, for example, the use of facultative or obligate anerobic bacteria.62 Therefore, the radiotherapeutic doses could be lowered to spare surrounding healthy tissues. Contrasted with another investigative approach, gene therapy, bacteriotherapy has the advantage of avoiding genetic modification of tumor cells, which is rather inefficient and not risk‐free. The combination of radiotherapy with bacteria is a novel active area of investigation. In spite of the fact that there have been few investigations using bacteria to promote radiotherapy, this field may still become a practical approach in clinical radiation oncology. It has been shown that genetically engineered Salmonella bacteria have the desired properties of an antitumor vector. They can selectively replicate within tumors, and can express effector genes such as the herpes simplex thymidine kinase. Salmonella that targets tumors from a distant inoculation site, can mediate tumor growth suppression.85 Lipid A‐altered Salmonella auxotrophs were developed that displayed attenuated toxicity in mice and swine. These mutants demonstrated considerably reduced induction of host TNF‐α (a key mediator in bacteria‐mediated tumor therapy), yet retained the capacity for tumor multiplication and growth suppression, achieving accumulation in tumors of 109 colony forming units (cfu)/g of tumor, which was 1000 times higher than normal tissues in mice.86, 87 The outcome was followed for tumor growth and mouse survival. When the bacteriotherapy was combined with radiation treatment it produced additional antitumor effects. In dose‐response studies with increasing doses of radiation but single dosages of Salmonella, the 2 agents together caused synergistic suppression of tumor growth (greater than additivity).88 The attenuated S typhimurium strain ΔppGpp (guanosine 5'‐diphosphate‐3'‐diphosphate) could be used to kill tumor cells. It was engineered to carry a pBAD plasmid coding for cytotoxic protein cytolysis A (ClyA) and also for bacterial luciferase (Lux) to allow optical imaging.46 ClyA is a 34‐kDa pore‐forming hemolytic protein, which can be produced S typhimurium and E coli without posttranslational modification.89 The ClyA cytotoxicity toward macrophages and mammalian cells induced cell caspase‐mediated cell death. A serious drawback of radiotherapy is that it is not clear whether the radiation affects cytotoxin‐expressing Salmonella that has localized in the tumor. Liu et al37 reported that the combination of radiotherapy and bacterial therapy using engineered S typhimurium ∆ppGpp reduced tumor growth compared with bacterial therapy alone. In addition, the researchers recently showed that the agonist of toll‐like receptor 5 (TLR5), called bacterial flagellin, as well as CBLB502 (its pharmacologically optimized derivative) could protect primates and rodents from gastrointestinal and hematopoietic radiation syndromes induced by total body irradiation.84, 90 In another investigation, Platt et al88 showed that the combination of X‐rays with Salmonella had supra‐additive antitumor effects, with a larger slope of the dose‐response curve. They additionally proposed that at higher radiation doses (25‐50 Gy), the supra‐additive effect was not clear, as this would have needed a full fractional dose‐response. Felgner et al40 recently reported that pre‐exposure to therapeutic bacteria (E coli Symbioflor‐2 or Salmonella SL7207 vector strains) in naive mice and tumor bearing mice that were subsequently immunized, caused a significantly difference in the phenotype of the microenvironment of colonized tumors. Bettegowda et al91 used a combination of spores of C novyi‐NT together with radiation therapy to treat transplanted tumors in mice. Their results indicated that C novyi‐NT spores alone only had very minor therapeutic effects, while the combination resulted in long‐term remission in a significant fraction of animals. In their studies partial and complete responses were found with the combination of external beam therapy or brachytherapy, and a single dose of C novyi‐NT. Nevertheless, they recommended that the combination therapy using conventional doses of radiation could be toxic for organs like the liver. On the other hand, they suggested that a combination of radioactive iodine with C novyi‐NT might enable patients to be treated with lower doses of radiolabeled antibodies, subsequently limiting toxicity to normal tissues, for example, to bone marrow. Nuyts et al32 tested bacteria as a potential gene delivery agent and combined it with radiation therapy. They isolated 2 radiation‐inducible genes of the SOS repair system (recA and recN genes) in Clostridium acetobutylicum DSM792, and confirmed these genes could be activated at a radiation dose of 2 Gy. These results suggested that fractionated radiotherapy could trigger repeated gene induction in bacteria leading to enhanced and prolonged protein expression.32 They suggested that the radio‐responsive recA promoter could increase TNFa production in recombinant clostridia after 2 Gy of irradiation.83 Jiang et al10 assessed the antitumor effects of combining RT with bacteria. In this investigation, E coli carrying pAClyA was injected into CT26‐bearing BALB/c mice, after which they were irradiated with different radiation doses (0, 8, 15, 21 Gy). This research showed that engineered bacteria such as E coli strain K‐12 can produce ClyA to enhance the therapeutic effects of radiation. Furthermore, their findings confirmed that radiotherapy and bacteriolytic therapy could inhibit the development of tumor metastasis. Table 2 summarizes the studies on the use of bacteria after or during radiotherapy as a combination cancer therapy.
Table 2.
Summaries of studies on combination of bacteriotherapy and radiotherapy for cancer therapy
| Strain(s) | Methods | Outcome | Ref |
|---|---|---|---|
| Clostridium novyi‐NT | Injection 3 × 108 spores of C novyi‐NT and irradiation; external beam radiation (0.1 Gy/s, Cs‐137 source), systemic radioimmunotherapy with 500µCi of I‐131‐labeled T84.66 mAb and brachytherapy (10 Gy/day) used of plaques loaded with I‐125 seeds in different transplanted tumors in mice | C novyi‐NT plus external beam radiation led to tumor shrinkage in mice bearing HCT116 tumors | 91 |
| C novyi with brachytherapy led to cure 100% of mice bearing HCT116 and HuCC‐T1 xenografts | |||
| Treated of the xenografts of colorectal cancer LS174T by combination of C novyi‐NT with radioimmunotherapy | |||
| C oncolyticum M55 | Injection C. oncolyticum M 55 spores with local tumor hyperthermy in mice bearing Ehrlich solid carcinoma, Harding‐Pasey‐melanoma and fibrosarcoma | Oncolysis of the tumors | 163 |
| C oncolyticum M55 | IV injection C. oncolyticum M 55 spores in combination with local tumor hyperthermy 2305 NMRI‐mice bearing neck tumors | intensification of the oncolysis tumors after 12 h | 164 |
| C oncolyticum M55 | local X‐irradiation and local HFH with iv spore‐ C. oncolyticum M55 in Harding‐Passey‐Melanoma‐bearing mice | Relatively cure | 165 |
| C acetobutylicum DSM792 | Induction of the recA and recN genes (involved in DNA repair) in Clostridium by radiation 2 Gy | Significant increase in b‐galactosidase activity | 32 |
| C acetobutylicum DSM792 | Induction of the gene involved in recA gene in Clostridium by radiation 2 Gy | Significant increase TNFα | 83 |
| E coli K‐12 | SC injection E coli containing expressing ClyA gene (5 × 107CFU) with 21 Gy radiation murine CT26 colon carcinoma cells | Tumor shrinkage, suppressed metastatic tumor growth and prolonged the survival time | 10 |
| Salmonella YS146 and YS166 | Combination treatment administration of X‐rays 5 to 15Gy with ip or iv injection Salmonella 2 × 105 cfu into mice bearing B16F10 or Cloudman S91 melanomas | Suppression of tumors growth and prolonged mice survival | 88 |
| Salmonella typhimurium SHJ2037 | Combination treatment radiotherapy 21Gy with iv injection S typhimurium (containing of Plasmid construction plasmid pBAD‐RLuc8‐clyA) into mice bearing colon tumor CT26 | Regression of tumors | 37 |
Several innovative approaches have been proposed to reduce radiotherapy‐induced normal tissue damage.92, 93 The application of bacteria could also be used to reduce normal tissue damage during or after RT. Certain strains of bacteria (particularly Lactobacilli and Bifidobacteria) have been proposed to reduce RT side effects.25, 94, 95, 96 The use of probiotics to preserve normal tissue during or after radiotherapy has also been shown in many clinical and preclinical studies.97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108
9. COMBINATION OF BACTERIOTHERAPY WITH CHEMOTHERAPY
Chemotherapy is still the mainstay of treatment for inoperable cancer, despite numerous shortcomings such as inadequate drug concentrations in tumors, occurrence of systemic toxicity (hematological, gastrointestinal, alopecia, heart, and skin toxicity) in many types of cancer, and almost inevitable induction of drug resistance.109, 110, 111, 112 Neutropenia is one of the main manifestations of hematological toxicity. It is well known that due to immunosuppression, neutropenia poses a risk of infectious disease occurring during treatment. In addition, chemotherapy is responsible for gastrointestinal toxicity because of mucosal damage, and altering the natural host microflora.113, 114, 115 It should be noted that tumor cells that remain after chemotherapy frequently show increased aggressiveness, and can enter blood and lymph vessels, thereby increasing the probability of metastasis.116, 117 Therefore the introduction of new approaches is required to increase effectiveness and reduce toxicity in chemotherapy.109 Combining chemotherapy with bacteriotherapy could be one of these new approaches. It should be pointed out that bacteria can specifically target the most hypoxic tumors which are often those most resistant to chemotherapy.118 Bacteria can, not only sensitize tumors to increase the efficiency of chemotherapy, but they can also be exploited as drug/gene delivery vehicles. Bacterial toxins can destroy tumors and can also be used for bacterial‐based cancer vaccines.6, 61 Genetically modified bacteria can be used for selective tumor targeting as well as bacterial gene‐directed enzyme prodrug therapy for cancer.6 Moreover, bacterial endotoxins can also be used to fight cancer particularly in combination with chemotherapy.7 Probiotic bacteria could mitigate the severity of chemotherapy‐induced toxicity, particularly the gastrointestinal side‐effects. VSL‐3 is 1 type of probiotic formula that has been effective in reducing complications (such as diarrhea induced by chemotherapy in rats).119 Whitford et al, showed that Streptococcus thermophilus has beneficial effects on 5‐fluorouracil (5‐FU) complications (intestinal mucositis).120 Bowen et al suggested that, in spite of the overall shortage of hard data, probiotics should be tested for chemotherapy complications.113 In another investigation, 150 patients with colorectal cancer receiving 5‐FU and leucovorin bolus injections plus continuous 5‐FU infusion as postoperative adjuvant therapy, were randomly allocated to receive L rhamnosus GG (1‐2 × 10(10) CFU) and fiber (11 g guar gum) per day, while others did not. The patients receiving Lactobacillus had significantly less severe grades of diarrhea. These participants also had less abdominal discomfort thereby reducing the need for hospital care and lowering of chemotherapy doses.121 Abd El‐Atti et al showed the effectiveness of probiotics to control chemotherapy complications in patients with advanced breast cancer.122 One of the most troubling treatment‐related complications in patients with head and neck cancer is oral mucositis. Some of the recent studies have investigated the positive effects of Lactobacillus brevis CD2 lozenges on the severity and prevalence of mucositis, as well as the tumor resistance to radiotherapy. It was observed that patients who received Lactobacillus during chemotherapy had fewer intestinal problems than the others, resulting in shortening the course of chemotherapy and lower doses.94, 102, 108, 112, 123, 124, 125 Another method to reduce the side effects of chemotherapy is the COBALT strategy (combination bacteriolytic therapy; simultaneous use of C novyi‐NT spores with conventional chemotherapeutic agents). Although COBALT showed meaningful antitumor effects, it could not completely prevent animal deaths.66 Another use of C novyi‐NT is its membrane‐disrupting potential in liposome‐encapsulated drug delivery to tumor cells.126, 127 Nitroreductase (NR) enzymes from different bacterial strains have been investigated in some studies.128 It was shown that the NR enzyme from Haemophilus influenza had promising pharmacokinetic properties and could be utilized in treating tumors in mice. Mice were treated either with C sporogenes alone, or with C sporogenes in combination with NR and CB1954.
A novel study demonstrated the power of botulinum neurotoxin (BoNT) to destroy tumor vessels, allowing greater cancer cell destruction by chemotherapy.129 The most important problem in this process was insufficient tumor lysis. Since all the components of malignant tissue are not completely consumed by bacteria, bacteriotherapy should be combined with chemotherapy.130 Salmonella and Clostridium produce the suicide enzyme cytosine deaminase (CDase) which transforms the pro‐drug 5‐fluorocytosine (5‐FC) to chemotherapeutic 5‐FU.42, 131, 132, 133 Nemunaitis et al in an experimental clinical trial used 5‐FC and recombinant Salmonella expressing CDase, and 66.7% of patients showed a tumor response.18 Another study investigated the ability and efficiency of Salmonella typhimurium VNP20009 in a murine melanoma model combined with different chemotherapy drugs.118 Kasinskas et al,134 discussed the relationship between S typhimurium and the microenvironments of solid tumors. They suggested that the interaction of S typhimurium with the microenvironment regulated the amount and location of bacterial accumulation. Through monitoring these interactions, they proposed that administration of S typhimurium could lead to increased effects of standard chemotherapeutic drugs. Exploiting S choleraesuis as a single tumor‐targeting anticancer agent in tumor‐bearing mice was reported by Lee et al.135 Their study indicated that the combination of S choleraesuis and cisplatin postponed tumor development and increased survival.
10. BACTERIA IN THERANOSTIC APPROACHES
Theranostics describes the use of multifunctional approaches to simultaneously image, monitor and treat tumors, and has recently attracted a great deal of attention.136, 137, 138 One of the common approaches to theranostics is to use drug‐delivery nanovesicles, that also incorporate an imaging component.139 Bacteria can be used in theranostics because they can specifically target tumors and can replicate in tumor cells.6, 139 The ease of genetic manipulation of bacteria allows for the production of attenuated strains with greater safety profiles, and vector systems, thereby allowing for precise tuning and multifunctional capabilities.7, 140 To facilitate the monitoring of migration patterns and to follow the proliferation of these bacteria, scientists use genetically modified bacteria to express reporter genes that allow optical imaging.10 In addition, bacteria can be detected by MRI or PET.7, 10, 140 Sheng‐Nan Jiang et al,10 used an E coli strain K‐12 (MG1655) producing the cytotoxic protein and pore‐forming hemolytic cytolysin A (to kill colon cancer cells) and the bacterial luciferase (Lux) operon (as an in vivo imaging marker). They found that a combination of bacteriotherapy and radiotherapy reduced tumor metastasis and increased the survival rate in mice. Cheng‐Hung Luo developed 2 approaches for cancer theranostics using Bifidobacterium breve and Clostridium difficile to increase the treatment and imaging effectiveness.141 In another study, Zurkiya et al142 applied magnetotactic bacteria (naturally producing magnetosomes) to deliver a gene expression marker (magA) in the human 293FT cell line as a candidate MRI reporter gene. Quispe‐Tintaya et al tested an engineered bacterial strain (virulence‐attenuated live Listeria monocytogenes with bound radioactive antibodies), instead of administering radiotherapy and bacterial tumor therapy independently. They observed that this construct led to primary tumor regression (>60%) and reduced metastases (>90%).143 Some bacteria (Haloarchaea) that produce gas vesicles (~width of 45‐250 nm and length of 100‐600 nm) have been examined as theranostic agents.144, 145 Recently, Shapiro et al146 described a novel diagnostic strategy using gas vesicles (harvested from Anabaena flosaquae and Halobacterium NRC‐1) as ultrasound contrast agents for molecular imaging in mice. In addition, they suggested that these vesicles could be targeted as therapeutic agents and drug or gene delivery vehicles.147
11. CONCLUSIONS AND PROSPECTIVES
The unique pathophysiology of solid tumors causes major obstacles for traditional anticancer therapies. There are advantages and disadvantages in the applications of therapeutic bacteria in cancer therapy.18, 134 Currently, although traditional cancer therapies are still the mainstream treatments, bacteriotherapy has demonstrated remarkable effects, thanks to its high specificity, ability to be controlled post‐administration, and oncolytic capabilities in many in vivo studies.34 Nevertheless, many problems remain for using bacteria in clinical practice as antitumor agents including; bacterial toxicity, DNA instability, limited targeting efficiency, choice of practical and safe bacterial strains, and testing combination with other therapies.7, 80, 148 Hopefully, these obstacles can be overcome by more sophisticated genetic engineering of tailored strains. In the future, genetically modified bacteria will be made more practical for both diagnostic and therapeutic anticancer applications, and to enhance radiotherapy, immunotherapy, or chemotherapy efficacy.
CONFLICT INTEREST
There are no conflicts of interest.
ACKNOWLEDGMENTS
The authors thank all those involved in the laboratory sciences research center, and Golestan Research Center of Gastroenterology and Hepatology of Golestan University of Medical Sciences, Gorgan, Iran who participated in the preparation of the manuscript.
Sedighi M, Zahedi Bialvaei A, Hamblin MR, et al. Therapeutic bacteria to combat cancer; current advances, challenges, and opportunities. Cancer Med. 2019;8:3167–3181. 10.1002/cam4.2148
Funding information
Dr Hamblin was supported by US NIH Grants R01AI050875 and R21AI121700.
REFERENCES
- 1. Thun MJ, DeLancey JO, Center MM, Jemal A, Ward EM. The global burden of cancer: priorities for prevention. Carcinogenesis. 2009;31:100‐110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Bhatt AP, Redinbo MR, Bultman SJ. The role of the microbiome in cancer development and therapy. CA Cancer J Clin. 2017;67:326‐344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Torre L, Bray F, Siegel R, Ferlay J, Lortet‐Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87‐108. [DOI] [PubMed] [Google Scholar]
- 4. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394‐424. [DOI] [PubMed] [Google Scholar]
- 5. Datta Nr, Ordóñez SG, Gaipl Us, et al. Local hyperthermia combined with radiotherapy and‐/or chemotherapy: Recent advances and promises for the future. Cancer Treat Rev. 2015;41:742‐753. [DOI] [PubMed] [Google Scholar]
- 6. Patyar S, Joshi R, Byrav DP, Prakash A, Medhi B, Das B. Bacteria in cancer therapy: a novel experimental strategy. J Biomed Sci. 2010;17:21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Forbes NS. Engineering the perfect (bacterial) cancer therapy. Nat Rev Cancer. 2010;10:785‐794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Nallar SC, Xu D‐Q, Kalvakolanu DV. Bacteria and genetically modified bacteria as cancer therapeutics: Current advances and challenges. Cytokine. 2017;89:160‐172. [DOI] [PubMed] [Google Scholar]
- 9. Wang K, Kievit FM, Zhang M. Nanoparticles for cancer gene therapy: Recent advances, challenges, and strategies. Pharmacol Res. 2016;114:56‐66. [DOI] [PubMed] [Google Scholar]
- 10. Jiang S‐N, Phan TX, Nam T‐K, et al. Inhibition of tumor growth and metastasis by a combination of Escherichia coli–mediated cytolytic therapy and radiotherapy. Mol Ther. 2010;18:635‐642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Parsonnet J. Bacterial infection as a cause of cancer. Environ Health Perspect. 1995;103:263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Mengesha A, Dubois L, Chiu RK, et al. Potential and limitations of bacterial‐mediated cancer therapy. Front Biosci. 2007;12:3880‐3891. [DOI] [PubMed] [Google Scholar]
- 13. Hajitou A, Trepel M, Lilley CE, et al. A hybrid vector for ligand‐directed tumor targeting and molecular imaging. Cell. 2006;125:385‐398. [DOI] [PubMed] [Google Scholar]
- 14. Kramer MG, Masner M, Ferreira FA, Hoffman RM. Bacterial therapy of cancer: promises, limitations, and insights for future directions. Front Microbiol. 2018;9:16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Kamta J, Chaar M, Ande A, Altomare DA, Ait‐Oudhia S. Advancing cancer therapy with present and emerging immuno‐oncology approaches. Front Oncol. 2017;7:64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Murphy C, Rettedal E, Lehouritis P, Devoy C, Tangney M. Intratumoural production of TNFα by bacteria mediates cancer therapy. PloS one. 2017;12:e0180034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Heimann DM, Rosenberg SA. Continuous intravenous administration of live genetically modified salmonella typhimurium in patients with metastatic melanoma. J Immunother. 2003;26:179‐180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Nemunaitis J, Cunningham C, Senzer N, et al. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene Ther. 2003;10:737‐744. [DOI] [PubMed] [Google Scholar]
- 19. Thamm DH, Kurzman ID, King I, et al. Systemic administration of an attenuated, tumor‐targeting Salmonella typhimurium to dogs with spontaneous neoplasia: phase I evaluation. Clin Cancer Res. 2005;11:4827‐4834. [DOI] [PubMed] [Google Scholar]
- 20. Song S, Vuai MS, Zhong M. The role of bacteria in cancer therapy–enemies in the past, but allies at present. Infect Agent Cancer. 2018;13:9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Wang C, Hu Q, Shen H‐M. Pharmacological inhibitors of autophagy as novel cancer therapeutic agents. Pharmacol Res. 2016;105:164‐175. [DOI] [PubMed] [Google Scholar]
- 22. Jia LJ, Wei DP, Sun QM, Huang Y, Wu Q, Hua ZC. Oral delivery of tumor‐targeting Salmonella for cancer therapy in murine tumor models. Cancer Sci. 2007;98:1107‐1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Nguyen VH, Min J‐J. Salmonella‐mediated cancer therapy: roles and potential. Nucl Med Mol Imaging. 2017;51:118‐126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Felgner S, Kocijancic D, Frahm M, et al. Engineered Salmonella enterica serovar Typhimurium overcomes limitations of anti‐bacterial immunity in bacteria‐mediated tumor therapy. OncoImmunology. 2018;7:e1382791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Kouhsari E, Ghadimi‐Daresajini A, Abdollahi H, et al. The potential roles of bacteria to improve radiation treatment outcome. Clin Transl Oncol. 2017;19:1‐13. [DOI] [PubMed] [Google Scholar]
- 26. Yu YA, Shabahang S, Timiryasova TM, et al. Visualization of tumors and metastases in live animals with bacteria and vaccinia virus encoding light‐emitting proteins. Nat Biotechnol. 2004;22:313‐320. [DOI] [PubMed] [Google Scholar]
- 27. Zhao M, Yang M, Li X‐m, et al. Tumor‐targeting bacterial therapy with amino acid auxotrophs of GFP‐expressing Salmonella typhimurium . Proc Natl Acad Sci USA. 2005;102:755‐760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hoffman RM, Zhao M. Whole‐body imaging of bacterial infection and antibiotic response. Nat Protoc. 2006;1:2988‐2994. [DOI] [PubMed] [Google Scholar]
- 29. Benoit Mr, Mayer D, Barak Y, et al. Visualizing implanted tumors in mice with magnetic resonance imaging using magnetotactic bacteria. Clin Cancer Res. 2009;15:5170‐5177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Tjuvajev J, Blasberg R, Luo X, Zheng L, King I, Bermudes D. Salmonella‐based tumor‐targeted cancer therapy: tumor amplified protein expression therapy (TAPET™) for diagnostic imaging. J Control Release. 2001;74:313‐315. [DOI] [PubMed] [Google Scholar]
- 31. Binder DC, Wainwright DA. The boosting potential of bacteria in cancer immunotherapy. Trends Mol Med. 2017;23:580‐582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Nuyts S, Van Mellaert L, Theys J, Landuyt W, Lambin P, Anné J. The use of radiation‐induced bacterial promoters in anaerobic conditions: a means to control gene expression in clostridium‐mediated therapy for cancer. Radiat Res. 2001;155:716‐723. [DOI] [PubMed] [Google Scholar]
- 33. Ginn SL, Amaya AK, Alexander IE, Edelstein M, Abedi MR. Gene therapy clinical trials worldwide to 2017: an update. J Gene Med. 2018;20:e3015. [DOI] [PubMed] [Google Scholar]
- 34. Zhang S, Chen Y, Wang J, Tang F, Miao T, Li M. Role of nontoxigenic Clostridium novyi in solid tumor therapy. Rev Med Microbiol. 2014;25:71‐76. [Google Scholar]
- 35. Zhou S, Gravekamp C, Bermudes D, Liu K. Tumour‐targeting bacteria engineered to fight cancer. Nat Rev Cancer. 2018;1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Dang L, Bettegowda C, Kinzler KW, Vogelstein B. Combination bacteriolytic therapy for the treatment of tumors. United States patent. US 8,007,782. 2011. [DOI] [PMC free article] [PubMed]
- 37. Liu X, Jiang S, Piao L, Yuan F. Radiotherapy combined with an engineered Salmonella typhimurium inhibits tumor growth in a mouse model of colon cancer. Exp Anim. 2016;65:413‐418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Toso JF, Gill VJ, Hwu P, et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J Clin Oncol. 2002;20:142‐152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Schlechte H, Elbe B. Recombinant plasmid DNA variation of Clostridium oncolyticum—model experiments of cancerostatic gene transfer. Zentralbl Bakteriol Mikrobiol Hyg A. 1988;268:347‐356. [DOI] [PubMed] [Google Scholar]
- 40. Felgner S, Pawar V, Kocijancic D, Erhardt M, Weiss S. Tumour‐targeting bacteria‐based cancer therapies for increased specificity and improved outcome. Microb Biotechnol. 2017;10:1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Lemmon MA, Bu Z, Ladbury JE, et al. Two EGF molecules contribute additively to stabilization of the EGFR dimer. EMBO J. 1997;16:281‐294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Theys J, Landuyt W, Nuyts S, et al. Specific targeting of cytosine deaminase to solid tumors by. Cancer Gene Ther. 2001;8:294‐297. [DOI] [PubMed] [Google Scholar]
- 43. Theys J, Nuyts S, Landuyt W, et al. Stable Escherichia coli‐Clostridium acetobutylicum shuttle vector for secretion of murine tumor necrosis factor alpha. Appl Environ Microbiol. 1999;65:4295‐4300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Groot AJ, Mengesha A, van der Wall E, van Diest PJ, Theys J, Vooijs M. Functional antibodies produced by oncolytic clostridia. Biochem Biophys Res Commun. 2007;364:985‐989. [DOI] [PubMed] [Google Scholar]
- 45. Jahanban‐Esfahlan R, de la Guardia M, Ahmadi D, Yousefi B. Modulating tumor hypoxia by nanomedicine for effective cancer therapy. J Cell Physiol. 2018;233:2019‐2031. [DOI] [PubMed] [Google Scholar]
- 46. Nguyen VH, Kim H‐S, Ha J‐M, Hong Y, Choy HE, Min J‐J. Genetically engineered Salmonella typhimurium as an imageable therapeutic probe for cancer. Cancer Res. 2010;70:18‐23. [DOI] [PubMed] [Google Scholar]
- 47. Ryan Rm, Green J, Williams PJ, et al. Bacterial delivery of a novel cytolysin to hypoxic areas of solid tumors. Gene Ther. 2009;16:329. [DOI] [PubMed] [Google Scholar]
- 48. Flentie K, Kocher B, Gammon ST, Novack DV, McKinney JS, Piwnica‐Worms D. A bioluminescent transposon reporter‐trap identifies tumor‐specific microenvironment‐induced promoters in Salmonella for conditional bacterial‐based tumor therapy. Cancer Discov. 2012;2:624‐637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Gelman AE, Turka LA. Autoimmunity heats up. Nature Med. 2003;9:1465‐1466. [DOI] [PubMed] [Google Scholar]
- 50. Kay A. Allergy and allergic diseases. N Engl J Med. 2001;344:30‐37. [DOI] [PubMed] [Google Scholar]
- 51. Avogadri F, Martinoli C, Petrovska L, et al. Cancer immunotherapy based on killing of Salmonella‐infected tumor cells. Cancer Res. 2005;65:3920‐3927. [DOI] [PubMed] [Google Scholar]
- 52. Furumoto K, Soares L, Engleman EG, Merad M. Induction of potent antitumor immunity by in situ targeting of intratumoral DCs. J Clin Invest. 2004;113:774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Higgins DE, Shastri N, Portnoy DA. Delivery of protein to the cytosol of macrophages using Escherichia coli K‐12. Mol Microbiol. 1999;31:1631‐1641. [DOI] [PubMed] [Google Scholar]
- 54. Reuter U, Oettmeier R, Hobohm U. Safety of therapeutic fever induction in cancer patients using approved PAMP drugs. Transl Oncol. 2018;11:330‐337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Zahaf N‐I, Schmidt G. Bacterial toxins for cancer therapy. Toxins. 2017;9:236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Ganai S, Arenas R, Forbes N. Tumour‐targeted delivery of TRAIL using Salmonella typhimurium enhances breast cancer survival in mice. Br J Cancer. 2009;101:1683‐1691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Zheng D‐W, Chen Y, Li Z‐H, et al. Optically‐controlled bacterial metabolite for cancer therapy. Nat Commun. 2018;9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Nougayrède J‐P, Taieb F, De Rycke J, Oswald E. Cyclomodulins: bacterial effectors that modulate the eukaryotic cell cycle. Trends Microbiol. 2005;13:103‐110. [DOI] [PubMed] [Google Scholar]
- 59. Maroccia Z, Loizzo S, Travaglione S, Frank C, Fabbri A, Fiorentini C. New therapeutics from nature: the odd case of the bacterial cytotoxic necrotizing factor 1. Biomed Pharmacother. 2018;101:929‐937. [DOI] [PubMed] [Google Scholar]
- 60. Oswald E, Sugai M, Labigne A, et al. Cytotoxic necrotizing factor type 2 produced by virulent Escherichia coli modifies the small GTP‐binding proteins Rho involved in assembly of actin stress fibers. Proc Natl Acad Sci. 1994;91:3814‐3818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Carswell E, Old LJ, Kassel R, Green S, Fiore N, An WB. endotoxin‐induced serum factor that causes necrosis of tumors. Proc Natl Acad Sci. 1975;72:3666‐3670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Gardlik R, Fruehauf JH. Bacterial vectors and delivery systems in cancer therapy. IDrugs. 2010;13:701‐706. [PubMed] [Google Scholar]
- 63. Bermudes D, Zheng L, King IC. Live bacteria as anticancer agents and tumor‐selective protein delivery vectors. Curr Opin Drug Discov Devel. 2002;5:194‐199. [PubMed] [Google Scholar]
- 64. Nuyts S, Theys J, Landuyt W, Lambin P, Anné J. Increasing specificity of anti‐tumor therapy: cytotoxic protein delivery by non‐pathogenic clostridia under regulation of radio‐induced promoters. Anticancer Res. 2001;21:857‐861. [PubMed] [Google Scholar]
- 65. Marth E, Möse JR. Oncolysis by Clostridium oncolyticum M55 and subsequent enzymatic determination of sialic acid in serum. Zentralbl Bakteriol Mikrobiol Hyg A. 1987;265:33‐44. [DOI] [PubMed] [Google Scholar]
- 66. Dang LH, Bettegowda C, Huso Dl, Kinzler Kw, Vogelstein B. Combination bacteriolytic therapy for the treatment of experimental tumors. Proc Natl Acad Sci. 2001;98:15155‐15160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Taniguchi S, Fujimori M, Sasaki T, et al. Targeting solid tumors with non‐pathogenic obligate anaerobic bacteria. Cancer Sci. 2010;101:1925‐1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Urbanska AM, Zhang X, Prakash S. Bioengineered colorectal cancer drugs: orally delivered anti‐inflammatory agents. Cell Biochem Biophys. 2015;72:757‐769. [DOI] [PubMed] [Google Scholar]
- 69. Yazawa K, Fujimori M, Nakamura T, et al. Bifidobacterium longum as a delivery system for gene therapy of chemically induced rat mammary tumors. Breast Cancer Res Treat. 2001;66:165‐170. [DOI] [PubMed] [Google Scholar]
- 70. Diaz LA, Cheong I, Foss CA, et al. Pharmacologic and toxicologic evaluation of C. novyi‐NT spores. Toxicol Sci. 2005;88:562‐575. [DOI] [PubMed] [Google Scholar]
- 71. Low KB, Ittensohn M, Le T, et al. Lipid A mutant Salmonella with suppressed virulence and TNFalpha induction retain tumor‐targeting in vivo. Nat Biotechnol. 1999;17:37‐41. [DOI] [PubMed] [Google Scholar]
- 72. Min J‐J, Kim H‐J, Park JH, et al. Noninvasive real‐time imaging of tumors and metastases using tumor‐targeting light‐emitting Escherichia coli. Mol Imaging Biol. 2008;10:54‐61. [DOI] [PubMed] [Google Scholar]
- 73. Min JJ, Nguyen VH, Kim HJ, Hong Y, Choy HE. Quantitative bioluminescence imaging of tumor‐targeting bacteria in living animals. Nat Protoc. 2008;3:629‐636. [DOI] [PubMed] [Google Scholar]
- 74. Agrawal N, Bettegowda C, Cheong I, et al. Bacteriolytic therapy can generate a potent immune response against experimental tumors. Proc Natl Acad Sci. 2004;101:15172‐15177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Dang L, Bettegowda C, Kinzler KW, Vogelstein B. Combination bacteriolytic therapy for the treatment of tumors. Google Patents. 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Yam C, Zhao M, Hayashi K, et al. Monotherapy with a tumor‐targeting mutant of S. typhimurium controls liver metastasis in a mouse model of pancreatic cancer. The Journal of surgical research. 2010;164:248–255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Theys J, Lambin P. Clostridium to treat cancer: dream or reality? Ann Transl Med. 2015;3:S21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Malmgren RA, Flanigan CC. Localization of the vegetative form of Clostridium tetani in mouse tumors following intravenous spore administration. Cancer Res. 1955;15:473‐478. [PubMed] [Google Scholar]
- 79. Kizaka‐Kondoh S, Inoue M, Harada H, Hiraoka M. Tumor hypoxia: a target for selective cancer therapy. Cancer Sci. 2003;94:1021‐1028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Van Mellaert L, Barbé S, Anné J. Clostridium spores as anti‐tumour agents. Trends Microbiol. 2006;14:190‐196. [DOI] [PubMed] [Google Scholar]
- 81. Harrison LB, Chadha M, Hill RJ, Hu K, Shasha D. Impact of tumor hypoxia and anemia on radiation therapy outcomes. Oncologist. 2002;7:492‐508. [DOI] [PubMed] [Google Scholar]
- 82. Wachsberger P, Burd R, Dicker AP. Tumor response to ionizing radiation combined with antiangiogenesis or vascular targeting agents. Explor Mech Interact. 2003;9:1957‐1971. [PubMed] [Google Scholar]
- 83. Nuyts S, Van Mellaert L, Theys J, et al. Radio‐responsive recA promoter significantly increases TNF [alpha] production in recombinant clostridia after 2 Gy irradiation. Gene Ther. 2001;8:1197. [DOI] [PubMed] [Google Scholar]
- 84. Burdelya LG, Gleiberman AS, Toshkov I, et al. Toll‐like receptor 5 agonist protects mice from dermatitis and oral mucositis caused by local radiation: implications for head‐and‐neck cancer radiotherapy. Inter J Radiat Oncol Biol Phys. 2012;83:228‐234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Pawelek JM, Low KB, Bermudes D. Tumor‐targeted Salmonella as a novel anticancer vector. Cancer Res. 1997;57:4537‐4544. [PubMed] [Google Scholar]
- 86. Low KB, Ittensohn M, Le T, et al. Lipid A mutant Salmonella with suppressed virulence and TNFα induction retain tumor‐targeting in vivo. Nat Biotechnol. 1999;17:37‐41. [DOI] [PubMed] [Google Scholar]
- 87. Bermudes D, Low B, Pawelek JM. Tumor‐targeted Salmonella: strain development and expression of the HSV‐tK effector gene. Gene Ther Cancer;2000:419‐436. [DOI] [PubMed] [Google Scholar]
- 88. Platt J, Sodi S, Kelley M, et al. Antitumour effects of genetically engineered Salmonella in combination with radiation. Eur J Cancer. 2000;36:2397‐2402. [DOI] [PubMed] [Google Scholar]
- 89. Wai SN, Lindmark B, Söderblom T, et al. Vesicle‐mediated export and assembly of pore‐forming oligomers of the enterobacterial ClyA cytotoxin. Cell. 2003;115:25‐35. [DOI] [PubMed] [Google Scholar]
- 90. Burdelya Lg, Krivokrysenko Vi, Tallant Tc, et al. An agonist of toll‐like receptor 5 has radioprotective activity in mouse and primate models. Science. 2008;320:226‐230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Bettegowda C, Dang Lh, Abrams R, et al. Overcoming the hypoxic barrier to radiation therapy with anaerobic bacteria. Proc Natl Acade Sci. 2003;100:15083‐15088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Abdollahi H. Beneficial effects of cellular autofluorescence following ionization radiation: hypothetical approaches for radiation protection and enhancing radiotherapy effectiveness. Med Hypotheses. 2015;84:194‐198. [DOI] [PubMed] [Google Scholar]
- 93. Brizel DM, Wasserman TH, Henke M, et al. Phase III randomized trial of amifostine as a radioprotector in head and neck cancer. J Clin Oncol. 2000;18:3339‐3345. [DOI] [PubMed] [Google Scholar]
- 94. Abdollahi H. Probiotic‐based protection of normal tissues during radiotherapy. Nutrition. 2014;30:495. [DOI] [PubMed] [Google Scholar]
- 95. Khademi S, Abdollahi H. Application of hydrogen producing microorganisms in radiotherapy: an idea. Iran J Public Health. 2014;43:1018‐1019. [PMC free article] [PubMed] [Google Scholar]
- 96. Abdollahi H, Atashzar M, Amini M. The potential use of biogas producing microorganisms in radiation protection. J Med Hypotheses Idea. 2015;9:67‐71. [Google Scholar]
- 97. Delia P, Sansotta G, Donato V, et al. Prevention of radiation‐induced diarrhea with the use of VSL# 3, a new high‐potency probiotic preparation. Am J Gastroenterol. 2002;97:2150. [DOI] [PubMed] [Google Scholar]
- 98. Demirer S, Aydıntug S, Aslım B, et al. Effects of probiotics on radiation‐induced intestinal injury in rats. Nutrition. 2006;22:179‐186. [DOI] [PubMed] [Google Scholar]
- 99. Giralt J, Regadera JP, Verges R, et al. Effects of probiotic Lactobacillus casei DN‐114 001 in prevention of radiation‐induced diarrhea: results from multicenter, randomized, placebo‐controlled nutritional trial. Int J Radiat Oncol Biol Phys. 2008;71:1213‐1219. [DOI] [PubMed] [Google Scholar]
- 100. Delia P, Sansotta G, Donato V, et al. Use of probiotics for prevention of radiation‐induced diarrhea. WJG. 2007;13:912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Chitapanarux I, Chitapanarux T, Traisathit P, Kudumpee S, Tharavichitkul E, Lorvidhaya V. Randomized controlled trial of live lactobacillus acidophilus plus bifidobacterium bifidum in prophylaxis of diarrhea during radiotherapy in cervical cancer patients. Radiat Oncol. 2010;5:31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Sharma A, Rath G, Chaudhary S, Thakar A, Mohanti BK, Bahadur S. Lactobacillus brevis CD2 lozenges reduce radiation‐and chemotherapy‐induced mucositis in patients with head and neck cancer: a randomized double‐blind placebo‐controlled study. Eur J Cancer. 2012;48:875‐881. [DOI] [PubMed] [Google Scholar]
- 103. Weill FS, Cela EM, Paz ML, Ferrari A, Leoni J, Maglio D. Lipoteichoic acid from Lactobacillus rhamnosus GG as an oral photoprotective agent against UV‐induced carcinogenesis. Br J Nutr. 2013;109:457‐466. [DOI] [PubMed] [Google Scholar]
- 104. Ki Y, Kim W, Nam J, et al. Probiotics for rectal volume variation during radiation therapy for prostate cancer. Inter J Radiat Oncol Biol Phys. 2013;87:646‐650. [DOI] [PubMed] [Google Scholar]
- 105. shao F, Xin F‐z, Yang C‐G, et al. The impact of microbial immune enteral nutrition on the patients with acute radiation enteritis in bowel function and immune status. Cell Biochem Biophys. 2014;69:357‐361. [DOI] [PubMed] [Google Scholar]
- 106. Ki Y, Kim W, Cho H, Ahn K, Choi Y, Kim D. The effect of probiotics for preventing radiation‐induced morphological changes in intestinal mucosa of rats. J Korean Med Sci. 2014;29:1372‐1378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. He J‐Y, Wang W‐Z, Qi H‐Z, Ma Y, He S‐Y. Use of recombinant Lactobacillus sakei for the prevention and treatment of radiation‐induced enteritis. Med Hypotheses. 2018;119:37‐40. [DOI] [PubMed] [Google Scholar]
- 108. Chang C‐W, Liu C‐Y, Lee H‐C, et al. Lactobacillus casei variety rhamnosus probiotic preventively attenuates 5‐fluorouracil/oxaliplatin‐induced intestinal injury in a syngeneic colorectal cancer model. Front Microbiol. 2018;9:983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Xu G, McLeod HL. Strategies for enzyme/prodrug cancer therapy. Clin Cancer Res. 2001;7:3314‐3324. [PubMed] [Google Scholar]
- 110. Evrard A, Cuq P, Ciccolini J, Vian L, Cano J. Increased cytotoxicity and bystander effect of 5‐fluorouracil and 5′‐deoxy‐5‐fluorouridine in human colorectal cancer cells transfected with thymidine phosphorylase. Br J Cancer. 1999;80:1726‐1733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Springer CJ, Niculescu‐Duvaz I. Prodrug‐activating systems in suicide gene therapy. J Clin Invest. 2000;105:1161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Ho CL, Tan HQ, Chua KJ, et al. Engineered commensal microbes for diet‐mediated colorectal‐cancer chemoprevention. Nat Biomed Eng. 2018;2:27. [DOI] [PubMed] [Google Scholar]
- 113. Mego M, Holec V, Drgona L, Hainova K, Ciernikova S, Zajac V. Probiotic bacteria in cancer patients undergoing chemotherapy and radiation therapy. Complement Ther Med. 2013;21:712‐723. [DOI] [PubMed] [Google Scholar]
- 114. Klastersky J. A review of chemoprophylaxis and therapy of bacterial infections in neutropenic patients. Diagn Microbiol Infect Dis. 1989;12:201‐207. [DOI] [PubMed] [Google Scholar]
- 115. Marshall JC. Gastrointestinal flora and its alterations in critical illness. Curr Opin Crit Care. 1999;5:119. [DOI] [PubMed] [Google Scholar]
- 116. Davis AJ, Tannock IF. Repopulation of tumour cells between cycles of chemotherapy: a neglected factor. Lancet Oncol. 2000;1:86‐93. [DOI] [PubMed] [Google Scholar]
- 117. Davis AJ, Tannock IF. Tumor physiology and resistance to chemotherapy: repopulation and drug penetration. Clin Relev Resist Cancer Chemother. 2002;1‐26. [DOI] [PubMed] [Google Scholar]
- 118. Jia L‐J, Wei D‐P, Sun Q‐M, et al. Tumor‐targeting Salmonella typhimurium improves cyclophosphamide chemotherapy at maximum tolerated dose and low‐dose metronomic regimens in a murine melanoma model. Int J Cancer. 2007;121:666‐674. [DOI] [PubMed] [Google Scholar]
- 119. Bowen JM, Stringer AM, Gibson RJ, Yeoh AS, Hannam S, Keefe DM. VSL# 3 probiotic treatment reduces chemotherapy‐induced diarrhoea and weight loss. Cancer Biol Ther. 2007;6:1445‐1450. [DOI] [PubMed] [Google Scholar]
- 120. Whitford EJ, Cummins AG, Butler RN, et al. Effects of Streptococcus thermophilus TH‐4 on intestinal mucositis induced by the chemotherapeutic agent, 5‐Fluorouracil (5‐FU). Cancer Biol Ther. 2009;8:505‐511. [PubMed] [Google Scholar]
- 121. Österlund P, Ruotsalainen T, Korpela R, et al. Lactobacillus supplementation for diarrhoea related to chemotherapy of colorectal cancer: a randomised study. Br J Cancer. 2007;97:1028‐1034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. El‐Atti SA, Wasicek K, Mark S, Hegazi R. Use of probiotics in the management of chemotherapy‐induced diarrhea: a case study. J Parenter Enteral Nutr. 2009;33:569‐570. [DOI] [PubMed] [Google Scholar]
- 123. Ali R, Zaharuddin L, Chan S‐N, Wong Z, Ngiu CS, Mokhtar NM. Sa1838‐The clinical and circulating inflammatory cytokines effects of probiotic containing Lactobacillus and Bifidobacterium strains in patients with colorectal cancer: a randomized double blind controlled trial. Gastroenterology. 2018;154:S‐414. [Google Scholar]
- 124. Thomsen M, Vitetta L. Adjunctive treatments for the prevention of chemotherapy‐and radiotherapy‐induced mucositis. Integr Cancer Ther. 2018;17:1027‐1047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Wei D, Heus P, van de Wetering FT, van Tienhoven G, Verleye L, Scholten RJ. Probiotics for the prevention or treatment of chemotherapy‐or radiotherapy‐related diarrhoea in people with cancer. Cochrane Database Syst Rev. 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Cheong I, Huang X, Bettegowda C, et al. A bacterial protein enhances the release and efficacy of liposomal cancer drugs. Science. 2006;314:1308‐1311. [DOI] [PubMed] [Google Scholar]
- 127. Cheong I, Zhou S. Tumor‐specific liposomal drug release mediated by liposomase. Methods Enzymol. 2009;465:251‐265. [DOI] [PubMed] [Google Scholar]
- 128. Theys J, Pennington O, Dubois L, et al. Repeated cycles of Clostridium‐directed enzyme prodrug therapy result in sustained antitumour effects in vivo. Br J Cancer. 2006;95:1212‐1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Ansiaux R, Gallez B. Use of botulinum toxins in cancer therapy. Expert Opin Invest Drugs. 2007;16:209‐218. [DOI] [PubMed] [Google Scholar]
- 130. Hatefi A, Canine BF. Perspectives in vector development for systemic cancer gene therapy. Gene Ther Mol Biol. 2009;13:15. [PMC free article] [PubMed] [Google Scholar]
- 131. Mei S, Theys J, Landuyt W, Anne J, Lambin P. Optimization of tumor‐targeted gene delivery by engineered attenuated Salmonella typhimurium . Anticancer Res. 2002;22:3261‐3266. [PubMed] [Google Scholar]
- 132. Dresselaers T, Theys J, Nuyts S, et al. Non‐invasive 19F MR spectroscopy of 5‐fluorocytosine to 5‐fluorouracil conversion by recombinant Salmonella in tumours. Br J Cancer. 2003;89:1796‐1801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Dubois L, Dresselaers T, Landuyt W, et al. Efficacy of gene therapy‐delivered cytosine deaminase is determined by enzymatic activity but not expression. Br J Cancer. 2007;96:758‐761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Kasinskas RW, Forbes NS. Salmonella typhimurium specifically chemotax and proliferate in heterogeneous tumor tissue in vitro. Biotechnol Bioeng. 2006;94:710‐721. [DOI] [PubMed] [Google Scholar]
- 135. Lee C‐H, Wu C‐L, Tai Y‐S, Shiau A‐L. Systemic administration of attenuated Salmonella choleraesuis in combination with cisplatin for cancer therapy. Mol Ther. 2005;11:707‐716. [DOI] [PubMed] [Google Scholar]
- 136. Ahmed N, Fessi H, Elaissari A. Theranostic applications of nanoparticles in cancer. Drug discov Today. 2012;17:928‐934. [DOI] [PubMed] [Google Scholar]
- 137. Xie J, Lee S, Chen X. Nanoparticle‐based theranostic agents. Adv Drug Deliv Rev. 2010;62:1064‐1079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018;16:214. [DOI] [PubMed] [Google Scholar]
- 139. Tietze R, Zaloga J, Unterweger H, et al. Magnetic nanoparticle‐based drug delivery for cancer therapy. Biochem Biophys Res Commun. 2015;468:463‐470. [DOI] [PubMed] [Google Scholar]
- 140. Zheng JH, Min J‐J. Targeted Cancer Therapy Using Engineered Salmonella typhimurium. Chonnam Med J. 2016;52:173‐184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Luo C‐H, Huang C‐T, Su C‐H, Yeh C‐S. Bacteria‐mediated hypoxia‐specific delivery of nanoparticles for tumors imaging and therapy. Nano Lett. 2016;16:3493‐3499. [DOI] [PubMed] [Google Scholar]
- 142. Zurkiya O, Chan AW, Hu X. MagA is sufficient for producing magnetic nanoparticles in mammalian cells, making it an MRI reporter. Magn Reson Med. 2008;59:1225‐1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Quispe‐Tintaya W, Chandra D, Jahangir A, et al. Nontoxic radioactive Listeriaat is a highly effective therapy against metastatic pancreatic cancer. Proc Natl Acad Sci. 2013;110:8668‐8673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Pfeifer F. Distribution, formation and regulation of gas vesicles. Nat Rev Microbiol. 2012;10:705‐715. [DOI] [PubMed] [Google Scholar]
- 145. Oren A. The function of gas vesicles in halophilic archaea and bacteria: theories and experimental evidence. Life. 2012;3:1‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Shapiro MG, Goodwill PW, Neogy A, et al. Biogenic gas nanostructures as ultrasonic molecular reporters. Nat Nanotechnol. 2014;9:311‐316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Ibsen S, Schutt CE, Esener S. Microbubble‐mediated ultrasound therapy: a review of its potential in cancer treatment. Drug des Devel Ther. 2013;7:375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Wei MQ, Ellem KA, Dunn P, West MJ, Bai CX, Vogelstein B. Facultative or obligate anaerobic bacteria have the potential for multimodality therapy of solid tumours. Eur J Cancer. 2007;43:490‐496. [DOI] [PubMed] [Google Scholar]
- 149. Fehleisen F. Ueber die Züchtung der Erysipelkokken auf künstlichem Nährboden und ihre Übertragbarkeit auf den Menschen. Dtsch Med Wochenschr. 1882;8:553‐554. [Google Scholar]
- 150. Busch W. Aus der Sitzung der medicinischen Section vom 13 November 1867. Berl Klin Wochenschr. 1868;5:137. [Google Scholar]
- 151. Pearl R. Cancer and Tuberculosis. Am J Hyg. 1929;9:97‐159. [Google Scholar]
- 152. Minton NP. Clostridia in cancer therapy. Nat Rev Microbiol. 2003;1:237‐242. [DOI] [PubMed] [Google Scholar]
- 153. Wood LM, Guirnalda PD, Seavey MM, Paterson Y. Cancer immunotherapy using Listeria monocytogenes and listerial virulence factors. Immunol Res. 2008;42:233‐245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Barbé S, Van Mellaert L, Theys J, et al. Secretory production of biologically active rat interleukin‐2 by Clostridium acetobutylicum DSM792 as a tool for anti‐tumor treatment. FEMS Microbiol Lett. 2005;246:67‐73. [DOI] [PubMed] [Google Scholar]
- 155. Dang LH, Bettegowda C, Agrawal N, et al. Targeting vascular and avascular compartments of tumors with C. novyi‐NT and anti‐microtubule agents. Cancer Biol Ther. 2004;3:326‐337. [DOI] [PubMed] [Google Scholar]
- 156. Agrawal N, Bettegowda C, Cheong I, et al. Bacteriolytic therapy can generate a potent immune response against experimental tumors. Proc Natl Acad Sci USA. 2004;101:15172‐15177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157. Xiong G, Husseiny MI, Song L, et al. Novel cancer vaccine based on genes of Salmonella pathogenicity island 2. Int J Cancer. 2010;126:2622‐2634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Colet WB. Late results of the treatment of inoperable sarcoma by the mixed toxins of erysipelas and bacillus prodigiosus. Trans Southern Surg Gynecol Ass. 1906;18:197. [Google Scholar]
- 159. Fiorentini C, Matarrese P, Straface E, et al. Toxin‐induced activation of Rho GTP‐binding protein increases Bcl‐2 expression and influences mitochondrial homeostasis. Exp Cell Res. 1998;242:341‐350. [DOI] [PubMed] [Google Scholar]
- 160. Falnes PØ, Ariansen S, Sandvig K, Olsnes S. Requirement for prolonged action in the cytosol for optimal protein synthesis inhibition by diphtheria toxin. J Biol Chem. 2000;275:4363‐4368. [DOI] [PubMed] [Google Scholar]
- 161. Kominsky SL, Vali M, Korz D, et al. Clostridium perfringens enterotoxin elicits rapid and specific cytolysis of breast carcinoma cells mediated through tight junction proteins claudin 3 and 4. Am J Pathol. 2004;164:1627‐1633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Puri RK. Development of a recombinant interleukin‐4‐Pseudomonas exotoxin for therapy of glioblastoma. Toxicologic Pathol. 1999;27:53‐57. [DOI] [PubMed] [Google Scholar]
- 163. Dietzel F, Gericke D, König W. Tumor hyperthermia using high frequency for increase of oncolysis by Clostridium butyricum (M 55). Strahlentherapie. 1976;152:537‐541. [PubMed] [Google Scholar]
- 164. Dietzel F, Gericke D. Intensification of the oncolysis by clostridia by means of radio‐frequency hyperthermy in experiments on animals–dependence on dosage and on intervals (author's transl). Strahlentherapie. 1977;153:263‐266. [PubMed] [Google Scholar]
- 165. Gericke D, Dietzel F, König W, Rüster I, Schumacher L. Further progress with oncolysis due to apathogenic clostridia. Zentralbl Bakteriol Orig A. 1979;243:102‐112. [PubMed] [Google Scholar]