

Abstract
Treatment with progesterone protects the male and female brain against damage after middle cerebral artery occlusion (MCAO). However, in both sexes, the brain contains significant amounts of endogenous progesterone. It is not known whether endogenously produced progesterone enhances the resistance of the brain to ischemic insult. Here, we used steroid profiling by gas chromatography-tandem mass spectrometry (GC-MS/MS) for exploring adaptive and sex-specific changes in brain levels of progesterone and its metabolites after MCAO. We show that, in the male mouse brain, progesterone is mainly metabolized via 5α-reduction leading to 5α-dihydroprogesterone (5α-DHP), also a progesterone receptor (PR) agonist ligand in neural cells, then to 3α,5α-tetrahydroprogesterone (3α,5α-THP). In the female mouse brain, levels of 5α-DHP and 3α,5α-THP are lower and levels of 20α-DHP are higher than in males. After MCAO, levels of progesterone and 5α-DHP are upregulated rapidly to pregnancy-like levels in the male but not in the female brain. To assess whether endogenous progesterone and 5α-DHP contribute to the resistance of neural cells to ischemic damage, we inactivated PR selectively in the CNS. Deletion of PR in the brain reduced its resistance to MCAO, resulting in increased infarct volumes and neurological deficits in both sexes. Importantly, endogenous PR ligands continue to protect the brain of aging mice. These results uncover the unexpected importance of endogenous progesterone and its metabolites in cerebroprotection. They also reveal that the female reproductive hormone progesterone is an endogenous cerebroprotective neurosteroid in both sexes.
SIGNIFICANCE STATEMENT The brain responds to injury with protective signaling and has a remarkable capacity to protect itself. We show here that, in response to ischemic stroke, levels of progesterone and its neuroactive metabolite 5α-dihydroprogesterone are upregulated rapidly in the male mouse brain but not in the female brain. An important role of endogenous progesterone in cerebroprotection was demonstrated by the conditional inactivation of its receptor in neural cells. These results show the importance of endogenous progesterone, its metabolites, and neural progesterone receptors in acute cerebroprotection after stroke. This new concept could be exploited therapeutically by taking into account the progesterone status of patients and by supplementing and reinforcing endogenous progesterone signaling for attaining its full cerebroprotective potential.
Keywords: aging, endogenous, neuroprotection, progesterone receptors, sex differences, steroids
Introduction
The very rapid loss of brain tissue after the onset of ischemic stroke represents a major therapeutic challenge (Saver, 2006). To date, the only acute treatments for ischemic stroke are thrombolysis with tissue plasminogen activator and removal of the thrombus with a retriever stent (Vivien et al., 2011; Campbell et al., 2015). These reperfusion therapies would benefit from the combined use of a cerebroprotective agent, which may prolong the survival of neural cells and extend the very short 4.5 h time window to reperfusion. In addition, a cerebroprotective agent may limit tissue damage resulting from reperfusion (Donnan et al., 2011; Fisher, 2011).
Treatment with exogenous progesterone has been demonstrated to protect the male and female rodent brain against ischemic damage after middle cerebral artery occlusion (MCAO) (Sayeed and Stein, 2009; Gibson et al., 2011). Notably, progesterone treatment shows long-lasting cerebroprotective efficacy with a 6 h window of opportunity (Gibson et al., 2011; Wali et al., 2014). However, it is unknown whether endogenous progesterone plays a role in cerebroprotective mechanisms. In adult males and females, neural cells are indeed exposed to significant levels of progesterone either derived from the ovaries in females or from the adrenal glands in both sexes (Schumacher et al., 2014). Moreover, progesterone is a “neurosteroid” synthesized de novo within the brain (Baulieu, 1997).
The aim of the present study was to assess whether endogenous progesterone and its metabolites are involved in acute self-protection of the male and female mouse brain against ischemic damage. This is an important problem because there is an ongoing interest in taking advantage of endogenous cerebroprotective mechanisms for therapeutic purposes (Iadecola and Anrather, 2011; Mergenthaler and Dirnagl, 2011). However, assessment of the cerebroprotective actions of progesterone requires taking into account its extensive metabolism in the brain (Schumacher et al., 2014). Therefore, progesterone is readily converted to 5α-dihydroprogesterone (5α-DHP) by neural cells (Celotti et al., 1992). In these cells, both progesterone and 5α-DHP activate gene transcription via progesterone receptors (PRs) (Rupprecht et al., 1993) (see Fig. 1). The PRs are ligand-activated transcription factors, but they also interact with membrane-associated kinases outside of the nucleus (Lange et al., 2007; Grimm et al., 2016). We have demonstrated recently that PRs play a key role in the cerebroprotective actions of progesterone after MCAO (Liu et al., 2012). The progesterone metabolite 5α-DHP is further converted to 3α,5α-tetrahydroprogesterone (3α,5α-THP), also well known as allopregnanolone, which does not bind to PR but functions as a positive modulator of GABAA receptors, resulting in the inhibition of neuronal activity (Fig. 1) (Hosie et al., 2006; Lambert et al., 2009). The conversion to 20α-dihydroprogesterone (20α-DHP) is an additional metabolic pathway of progesterone. Because 20α-DHP has low affinity for PRs and no progestogenic activity, the 20α reduction is generally considered as an inactivation pathway of progesterone (Fig. 1). However, progesterone and 20α-DHP are interconvertible and 20α-DHP may thus serve as a reservoir for progesterone and its biologically active metabolites (Hillisch et al., 2003; Williams et al., 2012).
Figure 1.

Schematic presentation of the single biosynthetic pathway starting with cholesterol and leading to progesterone, its metabolites, and their main signaling mechanisms. Enzymes are shown as numbers in italics next to the arrows indicating metabolic steps. Cholesterol is converted to pregnenolone inside the steroidogenic mitochondria by cytochrome P450scc (1). Carbons are numbered as shown. Pregnenolone is then converted to progesterone by the 3β-hydroxysteroid dehydrogenases (2) and progesterone to 5α-dihydroprogesterone (5α-DHP) by the 5α-reductases (3). Both progesterone and 5α-DHP bind to intracellular PRs, which regulate gene transcription inside the nucleus, but also interact with membrane-associated kinases in extranuclear compartments. The conversion of 5α-DHP to 3α,5α-THP (allopregnanolone) by the 3α-hydroxysteroid dehydrogenase (4) is a reversible reaction. In contrast to progesterone and 5α-DHP, 3α,5α-THP does not bind to PR, but is a positive modulator of GABAA receptors. The interconversion of 20-keto-pregnanes to 20α-hydroxy-pregnanes, considered as biologically inactive, is also reversible and catalyzed by a 20α-hydroxysteroid dehydrogenase (5).
We used sensitive and specific gas chromatography-tandem mass spectrometry (GC-MS/MS) for establishing steroid profiles in the brain and plasma of intact and ischemied male and female C57BL/6 mice. We found that levels of 5α-DHP and 3α,5α-THP were higher whereas levels of 20α-DHP were lower in the male brain than in the female brain. During the first 6 h after MCAO, there was a rapid and major increase in the levels of progesterone and 5α-DHP in the male brain, but, surprisingly, not in the female brain. To determine whether endogenous PR-active progesterone and 5α-DHP are involved in the acute resistance of the brain to ischemic damage, we inactivated PRs selectively in neural cells. Lack of neural PRs increased the vulnerability of the brains of both male and female young and aging mice.
Materials and Methods
Ethical statement
Procedures concerning animal care and use were performed in accordance with national guidelines (authorization 94-345 to R.G., animal facility approval 94-043-13), French ethical laws (Act 87-848 and Act 2013-11), and European Communities Council Directive (86/609/EEC) and have been approved by the ethical committee of French Ministry of Higher Education and Research C2EA-26 (project 2014_029). All experiments were performed following the ARRIVE guidelines (www.nc3rs.org.uk).
Generation of mice with conditional invalidation of PR gene in the nervous system
Generation of PR+/loxP mice.
PR+/loxP mice were generated by the Mouse Clinical Institute (MCI, Illkirch, France). Exon 2 of the PR genomic locus was targeted to generate a conditional allele. The modified PR allele PRloxP in which exon 2 was flanked by loxP sites was generated by homologous recombination in embryonic stem cells. Exon 2 encodes the first zinc finger of the DNA-binding domain of the receptor (Hill et al., 2012). The PRloxP/loxP mouse was crossed with a mouse expressing the Cre recombinase driven by the promoter and the CNS-specific enhancer of rat Nestin (Nes). In this mouse, the Cre recombinase is selectively expressed in CNS neurons, astrocytes, and oligodendrocytes, but not in microglial cells (Tronche et al., 1999). We used this Nestin-Cre mouse strain because of its well characterized efficiency of Cre-mediated recombination, resulting in the almost complete deletion of floxed steroid receptor genes in the brain (Tronche et al., 1999; Raskin et al., 2009).
Breeding colony establishment.
Two breeding colonies, PRloxP/loxP mice and PRNesCre mice (C57BL/6 background), were established in our animal facility (U1195 Inserm, Kremlin-Bicêtre). Male and female heterozygotes PR+/loxP mice were crossed. Pups were screened and PRloxP/loxP mice were selected. Male and female PRloxP/loxP mice were crossed to establish a PRloxP/loxP colony. We also generated a transgenic colony of PRNesCre mice that selectively lack PRs in the neural cells using the Cre-lox strategy. Male mice PR+/+ Tg (NesCre) were mated with female PRloxP/+ mice to obtain PR+/loxP Tg (NesCre) mice. Male PR+/loxP Tg (NesCre mice) were crossed with female PRloxP/loxP. Male PRloxP/loxP Tg (NesCre) were selected and crossed with female PRloxP/loxP to establish the breeding colony of PRloxP/loxP NesCre, which will be referred to as PRNesCre mice.
Mice were weaned at 21 d of age and each mouse was identified and genotyped by PCR using genomic DNA from tail to assess the presence of loxP sites and Cre recombinase using specific primers. Screening for LoxP sites was as follows: F1: 5′-AACATGGAAGGATGCACCTGTTCCC-3′; R1: 5′-GCCTTGAACTCATGAATTCCCAGCTTC-3′; F2: 5′-CCTTCCCTCTCATTGTCATG GAGAC-3′; R2: 5′-AGAGAATCTCTCGCCAGTGTCATGG-3′. Screening for Nes Cre transgene was as follows: F3: 5′-GCGGTCTGGCAGTAAAAACTATC-3′; R3: 5′-GTGAAACAGCATTGCT GTCACTT-3′. The internal positive control for PCR amplification was as follows: F4: 5′-CTAGGCCACAGAATT GAAAGATCT-3′; R4: 5′-GTAGGTGGAAATTCTAGCATCATCC-3′). Mice were group housed in a temperature-controlled room (22°C) and photoperiod (12 h light/12 h dark; lights on from 8:00 A.M. to 8:00 P.M.), with food and water available ad libitum.
Transient MCAO model
Transient cerebral ischemia was performed under ketamine (50 mg/kg) and xylazine hydrochloride (6 mg/kg) anesthesia. Body temperature of the mice was monitored by a rectal probe and maintained constant at 36–38°C during surgery with the aid of a homeothermic blanket control unit (Harvard Apparatus). Cerebral blood flow was monitored by laser Doppler flowmetry (Moor Instruments) 5 min before and 5 min after MCAO. A >70% drop of blood flow was considered a successful MCAO and mice that did not reach the required decrease were excluded from the study.
MCAO was performed for 30 min as described previously (Gibson and Murphy, 2004). Briefly, the left common carotid artery was isolated. A nylon monofilament (83 μm diameter; Drennan) coated with thermo-melting glue (4 mm long, 190 μm diameter) was introduced through an arteriotomy performed in the common carotid artery and advanced into the internal carotid artery to occlude the origin of the middle cerebral artery. The filament was withdrawn 30 min after occlusion to allow reperfusion.
Sham-operated mice underwent the same surgical procedures except that no filament was inserted. After reperfusion, the skin was adapted with a surgical suture. Mice received 0.5 ml of NaCl 0.9% subcutaneously. Animals were placed carefully in a heated cage to control body temperature and given moistened, soft pellets in a Petri dish placed in the cages to encourage eating.
Collecting brains and plasma for steroid profiling by GC-MS/MS
Mice were housed in a temperature-controlled room on a 12 h light/12 h dark cycle (lights on from 8:00 A.M. to 8:00 P.M.). For plasma and brain sampling, mice were handled gently to minimize stress and were quickly decapitated by rapid cervical dislocation without anesthesia. The brain was dissected out and the hemispheres were separated and quickly frozen on dry ice and stored at −80°C until steroid level analysis by GC-MS/MS.
Experimental groups
Set 1.
Set 1 was designed to evaluate the effect of ischemia on steroid levels. C57BL/6 young adult male and female mice (3 months old) were purchased from Janvier and were included in the experimental set after 1 week of acclimatization. Female mice were used at their diestrus phase. The estrus cycle was monitored between 9:00 and 10:00 A.M. by the examination of vaginal smears in a large group of intact young adult females. The adopted nomenclature was proestrus, estrus, metestrus, and diestrus according to the percentage of different cell types present as follows: proestrus (a majority of nucleated epithelial cells), estrus (a majority of cornified epithelial cells), metestrus (cornified epithelial cells and leukocytes), and diestrus (a majority of leukocytes) (Byers et al., 2012). The estrus cycle stage was checked in all females and those in diestrus were selected to constitute experimental groups of that day.
Twelve mice (6 males and 6 females) were used as controls to provide reference values for brain steroid levels in intact C57BL/6 mice. A total of 100 C57BL/6 mice (50 males and 50 females) were subjected to MCAO for 30 min. Sets of 20 mice (10 males and 10 females) were killed at 1, 2, 4, 6, and 24 h after MCAO.
Set 2.
Set 2 was designed to evaluate the effects of neural PR deletion on steroid levels in both sexes. Young adult male PRloxP/loxP and PRNesCre mice on a C57BL/6 background (3–4 months old, 26–30 g, n = 6 per group) and young adult female PRloxP/loxP and PRNesCre mice (3–4 months old, 21–26 g, diestrus phase, n = 6 per group) were used. The estrus cycle stage was checked in all females and those in diestrus were selected to constitute experimental groups of that day.
Set 3.
Set 3 was designed to evaluate the effect of neural PR deletion and MCAO on steroid levels. Young adult male PRloxP/loxP and PRNesCre mice (3–4 months old, 26–30 g, n = 9 per group), young adult female PRloxP/loxP and PRNesCre mice (3–4 months old, 21–26 g, diestrus phase, n = 9 per group) were subjected to 30 min of MCAO (Gibson and Murphy, 2004) or sham operation.
Steroid measurements by GC-MS/MS
Steroids were extracted from plasma and the left hemisphere (180–240 mg) of brain with methanol and the following internal standards were added to the extracts for steroid quantification: 2 ng of 2H6-5α-DHP (CDN Isotopes) for the analysis of 5α/β-dihydroprogesterone (5α/β-DHP); 2 ng of 13C3-testosterone (Isoscience) for the analysis of testosterone; 2 ng of 13C3-androstenedione for the analysis of androstenedione; 2 ng of 13C3-progesterone for the analysis of progesterone; 2 ng of 13C3-17β-estradiol for the analysis of 17β-estradiol, 2H13-deoxycorticosterone (DOC) for the analysis of DOC; 2 ng of 19 nor-PROG for the analysis of 20α-DHP and 5α/β-dihydrodeoxycorticosterone; 2 ng of epietiocholanolone (Steraloids) for the analysis of α-DHT; 3α5α-tetrahydrotestosterone, pregnenolone, 20α-dihydropregnenolone, 3α/β5α/β-tetrahydroprogesterone, 5α20α-tetrahydroprogesterone, 3α/β5α/β-hexahydroprogesterone, and 3α5α/β-tetrahydrodeoxycorticosterone, and 2H8-corticosterone for the analysis of corticosterone. Samples were purified and fractionated by solid-phase extraction with the recycling procedure (Liere et al., 2004). Briefly, the extracts were dissolved in 1 ml of methanol and applied to the C18 cartridge (500 mg, 6 ml; International Sorbent Technology), followed by 5 ml of methanol/H2O (85/15). The flow-through containing the free steroids was collected and dried. After a previous reconditioning of the same cartridge with 5 ml of H2O, the dried samples were dissolved in methanol/H2O (2/8) and reapplied. The cartridge was then washed with 5 ml of H2O and 5 ml of methanol/H2O (1/1) and unconjugated steroids were eluted with 5 ml of methanol/H2O (9/1).
The unconjugated steroid-containing fraction was then filtered and further purified and fractionated by HPLC. The HPLC system was composed of a WPS-3000SL analytical autosampler and a LPG-3400SD quaternary pump gradient coupled with a SR-3000 fraction collector (Thermo Fisher Scientific). The HPLC separation was achieved with a Lichrosorb Diol column (25 cm, 4.6 mm, 5 μm) and a thermostatic block at 30°C. The column was equilibrated in a solvent system of 90% hexane and 10% of a mixture composed of hexane/isopropanol (85/15). Elution was performed at a flo -rate of 1 ml/min, first 90% hexane and 10% of hexane/isopropanol (85/15) for 8 min, then with a linear gradient to 100% of hexane/isopropanol (85/15) in 2 min. This mobile phase was kept constant for 10 min and a linear gradient to 100% methanol was applied. The column was washed with methanol for 15 min.
Three fractions were collected from the HPLC system: 5α/β-DHPROG was eluted in the first HPLC fraction (3–13 min) and was silylated with 50 μl of a mixture N-methyl-N-trimethylsilyltrifluoroacetamide/ammonium iodide/dithioerythritol (1000:2:5 vol/w/w) for 15 min at 70°C. The second fraction (13–25 min) containing pregnenolone, 20α-dihydropregnenolone, progesterone, deoxycorticosterone, testosterone, and their reduced metabolites and 17β-estradiol was derivatized with 25 μl of heptafluorobutyric anhydride (HFBA) and 25 μl of anhydrous acetone for 1 h 20°C. Corticosterone was eluted in the third fraction (25–33 min) and derivatized with 25 μl of HFBA and 25 μl of anhydrous acetone for 1 h at 80°C. All fractions were dried under a stream of N2 and resuspended in hexane for GC-MS/MS analysis.
GC-MS/MS analysis of the brain extracts was performed using an AI 1310 autosampler, a Trace 1310 GC, and a TSQ 8000 MS/MS (Thermo Fisher Scientific) using argon as the collision gas. Injection was performed in the splitless mode at 250°C (1 min of splitless time) and the temperature of the GC oven was initially maintained at 50°C for 1 min and ramped between 50°C and 200°C at 20°C/min, then ramped to 300°C at 5°C/min, and finally ramped to 350°C at 30°C/min. The helium carrier gas flow was maintained constant at 1 ml/min during the analysis. The transfer line and ionization chamber temperatures were 330°C and 180°C, respectively. Electron impact ionization was used for mass spectrometry with ionization energy of 70 eV. GC-MS/MS signals were evaluated using a computer workstation by means of the software Excalibur, release 3.0 (Thermo Fisher Scientific). Identification of steroids was supported by their retention time and according to two or three transitions. Quantification was performed according to the transition giving the more abundant product ion (Table 1) with a previously established calibration curve.
Table 1.
GC-MS/MS parameters for steroid identification and quantification in multiple reaction monitoring
| Steroid (mol wt) | Derivatized steroid (mol wt) | Retention time (min) | Transition (m/z→m/z) | Collision energy (eV) |
|---|---|---|---|---|
| Pregnenolone (316) | Pregnenolone-3-HFB (512) | 21.24 | 298→283 | 8 |
| P (314) | P-3-HFB (510) | 21.34 | 510→425 | 14 |
| 5α-DHP (316) | 5α-DHP-3,20-TMS2 (460) | 18.63 | 460→425 | 12 |
| 5β-DHP (316) | 5β-DHP-3,20-TMS2 (460) | 17.30 | 460→445 | 10 |
| 3α5α-THP (318) | 3α5α-THP-3-HFB (514) | 20.23 | 429→215 | 8 |
| 3β5α-THP (318) | 3β5α -THP-3-HFB (514) | 21.62 | 429→215 | 8 |
| 3α5β-THP (318) | 3α5β-THP-3-HFB (514) | 20.39 | 496→481 | 12 |
| 20α-DHP (316) | 20α-DHP-3,20-HFB2 (708) | 18.48 | 708→693 | 10 |
| 5α20α-THP (318) | 5α20α-THP-3-HFB (514) | 22.93 | 514→499 | 8 |
| 3α5α20α-HHP (320) | 3α5α20α-HHP-3,20-HFB2 (712) | 17.40 | 712→429 | 8 |
| 3β5α20α-HHP (320) | 3β5α20α-HHP-3,20-HFB2 (712) | 18.75 | 697→483 | 10 |
| 3α5β20α-HHP (320) | 3α5β20α-HHP-3,20-HFB2 (712) | 17.59 | 712→429 | 8 |
| 20α-DHPREG (318) | 20α-DHPREG-3,20-HFB2 (710) | 18.00 | 496→211 | 10 |
| Testosterone (288) | Testosterone-3,17-HFB2 (680) | 15.98 | 680→320 | 8 |
| 5α-DHT (290) | 5α-DHT-17-HFB (486) | 20.16 | 486→414 | 10 |
| 3α5α-THT (292) | 3α5α-THT-3,17-HFB2 (684) | 15.00 | 470→455 | 8 |
| 17β-estradiol (272) | 17β-estradiol-3,l7-HFB2 (664) | 16.60 | 664→237 | 10 |
| DOC (330) | DOC-3,21-HFB2 (722) | 21.80 | 722→707 | 10 |
| 5α-DHDOC (332) | 5α-DHDOC-3,21-HFB (528) | 25.55 | 301→273 | 8 |
| 5β-DHDOC (332) | 5β-DHDOC-3,21-HFB (528) | 25.23 | 301→255 | 8 |
| 3α5α-THDOC (334) | 3α5α-THDOC-3,17-HFB2 (726) | 20.69 | 499→257 | 8 |
| 3α5β-THDOC (334) | 3α5β-THDOC-3,17-HFB2 (726) | 20.83 | 499→257 | 10 |
| Corticosterone (346) | Corticosterone-3,21-HFB2-H2O (720) | 20.31 | 705→355 | 10 |
Steroids were identified according to their retention time and their specific transition. Transitions were performed by selecting a precursor ion of a targeted steroid by the first mass spectrometer. This precursor ion was then fragmented at a collision energy chosen to give rise to a maximal signal of the product ion analyzed by the second mass spectrometer. The quantification was determined by the transition giving the most abundant product ion. P, Progesterone; DHP, dihydroprogesterone; THP, tetrahydroprogesterone; HHP, hexahydroprogesterone; DHPREG, dihydropregnenolone; DHT, dihydrotestosterone; THT, tetrahydrotestosterone; DOC, 11-deoxycorticosterone; DHDOC, dihydro-11-deoxycorticosterone; THDOC, tetrahydro-11-deoxycorticosterone; HFB, heptafluorobutyrate; TMS, trimethylsilyl.
The analytical protocol has been validated for all the targeted steroids using extracts of 200 mg of from a pool of male mice brain. The evaluation included the limit of detection, linearity, accuracy, intra-assay, and inter-assay precision (Table 2). The limit of detection was determined as the lowest amount of compounds that can be measured by GC-MS/MS with a signal-to-noise ratio >3 and ranged from 0.5 to 20 pg/g. The linearity was assessed by analyzing increasing amounts of mouse brain extracts (20, 50, 100, and 200 mg) in triplicate. The linearity was satisfactory for all the steroids with a coefficient of correlation ranging from 0.992 to 0.999. The accuracy of the assay was evaluated by determining the analytical recovery, which was defined as C/(C0 + S) × 100(%), where C is the concentration of the steroid in the spiked brain extract (100 mg), C0 is the concentration of a steroid in the unspiked brain extract (100 mg), and S is the spiked concentration. The precision of the intra-assays and inter-assays were evaluated by analyzing 5 replicates of 200 mg of brain extracts on 1 d and over 4 d, respectively.
Table 2.
Validation of the GC-MS/MS analytical procedure in mouse brain
| Steroid (mol wt) | Detection limit (pg/g) | Accuracy (%) | Intra-assay variation (%) | Inter-assay variation (%) |
|---|---|---|---|---|
| Pregnenolone (316) | 2 | 102.9 | 5.1 | 5.5 |
| P (314) | 20 | 105.9 | 2.2 | 3.6 |
| 5α-DHP (316) | 5 | 101.3 | 3.2 | 4.8 |
| 5β-DHP (316) | 2 | 103.4 | 3.6 | 6.8 |
| 3α5α-THP (318) | 10 | 97.5 | 5.4 | 7.4 |
| 3β5α-THP (318) | 0.5 | 99.6 | 4.5 | 5.6 |
| 3α5β-THP (318) | 10 | 105.1 | 6.2 | 6.5 |
| 20α-DHP (316) | 10 | 103.6 | 8.2 | 9.2 |
| 5α20α-THP (318) | 1 | 106.2 | 4.3 | 5.1 |
| 3α5α20α-HHP (320) | 5 | 98.4 | 3.2 | 3.5 |
| 3β5α20α-HHP (320) | 5 | 99.6 | 2.9 | 3.0 |
| 3α5β20α-HHP (320) | 5 | 95.3 | 2.6 | 2.7 |
| 20α-DHPREG (318) | 1 | 107.1 | 1.5 | 2.5 |
| Testosterone (288) | 1 | 99.8 | 1.1 | 1.8 |
| 5α-DHT (290) | 2 | 101.4 | 3.9 | 4.3 |
| 3α5α-THT (292) | 1 | 102.6 | 3.6 | 4.9 |
| 17β-estradiol (272) | 1 | 96.4 | 1.3 | 1.9 |
| DOC (330) | 5 | 98.2 | 1.5 | 1.7 |
| 5α-DHDOC (332) | 5 | 102.6 | 5.4 | 5.8 |
| 5β-DHDOC (332) | 5 | 104.5 | 6.1 | 6.9 |
| 3α5α-THDOC (334) | 2 | 10.10.4 | 4.9 | 6.1 |
| 3α5β-THDOC (334) | 2 | 98.2 | 3.4 | 4.0 |
| Corticosterone (346) | 20 | 97.6 | 2.5 | 3.0 |
The analytical protocol has been validated for all targeted steroids using 200 mg extracts from a pool of male mouse brains. The limit of detection was determined as the lowest amount of compounds that could be measured by GC-MS/MS with a signal-to-noise ratio >3. The linearity was assessed by analyzing increasing amounts of mouse brain extracts (20, 50, 100, and 200 mg) in triplicate with a coefficient of correlation ranging from 0.992 to 0.999. The accuracy of the assay was evaluated by determining the analytical recovery, which was defined as C/(C0 + S) × 100(%), where C is the concentration of the steroid in the spiked brain extract (200 mg), C0 is the concentration of a steroid in the unspiked brain extract (200 mg), and S is the spiked concentration. The precision of the intra-assays and inter-assays were evaluated by analyzing 5 replicates of 200 mg brain extracts on 1 d and over 4 d, respectively.
Abbreviations of steroids are as for Table 1.
Quantitative RT-PCR analysis of PR mRNA expression
Young adult male and female (4 months old) wild-type (WT) PR+/+ (C57BL/6), PRloxP/loxP, and PRNesCre mice (n = 4 per group) were used. Brains minus cerebellum were dissected out and separated into three regions: hypothalamus, cortex, and subcortical regions (all the remaining tissue). Peripheral tissues expressing PRs (http://www.nursa.org) were also collected: aorta, kidney, and thymus from males and uterus, ovaries, and mammary glands from females. Samples were quickly frozen on dry ice and stored at −80°C. Total RNA was isolated from tissue samples using the RNeasy Lipid Tissue Mini Kit (Qiagen) following the manufacturer's instructions. cDNA templates were synthesized using a SuperScript II ribonuclease H reverse transcriptase kit (Invitrogen). Primers for PR were chosen in exon 2 of PR gene. Cyclophilin 3 (Cyc3) was chosen as the housekeeping gene. Primers were designed using Primer Express version 3.0, Applied Biosystems as follows: PR F: 5′-TTACCTGT GGGAGCTGCAAG-3′; PR R: 5′-ATGCCAGCCTGACAACACTT-3′; Cyc3 F: 5′-CCTGGCT GGGTGAGCATGGC-3′; Cyc3 R: 5′-CAGTGGCTTGTCCCGGCTGT-3′). The relative PR mRNA expression was determined by real-time quantitative PCR using the ABI Prism 7300 sequence detection system (Applied Biosystems) and 2−▵▵CT method (Livak and Schmittgen, 2001). Correlation coefficient (r2) and efficiency (Ex) of PCR amplification were calculated before quantification (Peinnequin et al., 2004) as follows PR: Ex = 0.90; r2 = 0.999; Cyc3: Ex = 0.96; r2 = 0.997.
Immunofluorescence and confocal analyses of PR protein expression
Male and female (3 months old) PRloxP/loxP and PRNesCre mice (n = 3 per group) were injected with estradiol benzoate in sesame oil (10 μg/kg/d, s.c., 2 d). This dose and duration of treatment have been shown to induce PR expression (Brown et al., 1987). Animals were anesthetized with ketamine and perfused through the heart with PBS followed by 4% paraformaldehyde. Brains and uteri were dissected out and postfixed in the same fixative overnight. Tissue samples were cut into 50 μm slices using Vibratome and stored in PBS azide 0.05% at 4°C. To detect PR protein, immunofluorescence analysis was performed on free-floating sections using a rabbit polyclonal antibody (A0098; DAKO) directed against the DNA-binding domain of the human PR. This antibody was raised against a peptide sequence (aa 533–547) present in both the A and B isoforms of PRs (Traish and Wotiz, 1990). Slices were washed three times in PBS 1×; pH 7.6 (5 min each), incubated in PBS 1× containing 3% normal goat serum (NGS, Sigma-Aldrich) and 0.5% Triton (20 min at room temperature, RT), and then incubated with anti-PR antibody 1/500 in PBS 1× containing 1% NGS (72 h, at 4°C). Sections were rinsed in PBS 1× and incubated for 2 h at RT with the fluorescent secondary antibody (goat anti-rabbit IgG Alexa Fluor 488, A11008; Invitrogen) diluted at 1:200 in PBS 1×. Control of immunostaining was shown by substitution of the antibody by nonimmune serum on adjacent sections. In this case, no staining was observed. Sections were then rinsed with PBS 1× three times (5 min each) and mounted on slices using PBS/gelatin/azide/fluoromount. Labeling was visualized using a confocal Zeiss LSM 510 microscope. Sequences were acquired with green (488 nm) excitation.
Outcomes at 6 h after transient MCAO
Young adult male PRloxP/loxP and PRNesCre mice (3–4 months old, 26–30 g, n = 9 per group), young adult female PRloxP/loxP and PRNesCre mice (3–4 months old, 21–26 g, diestrus phase, n = 9 per group), and aging PRloxP/loxP and PRNesCre male and female mice (12–14 months old, n = 10 per group) were subjected to 30 min MCAO. Three died during surgery and a further four mice were excluded because of a <70% drop in blood flow. Analysis was performed at 6 h after MCAO.
Infarct volumes (TTC staining).
After cervical dislocation, brains were removed immediately and cut into 8 × 1 mm coronal sections using a MacIlwain tissue chopper (Mickle Laboratory Engineering). To quantify the ischemic damage, slices were rapidly immersed into a 2% 2,3,5-triphenyltetrazolium chloride (TTC; Sigma-Aldrich) solution for 15–20 min at room temperature (Türeyen et al., 2004). The brain slices were scanned and the area of damaged unstained brain tissues was measured on the posterior surface of each section using ImageJ 1.46f software. To correct for brain swelling, infarct volume of each mouse was multiplied by the ratio of the volume of intact contralateral hemisphere to ischemic ipsilateral hemisphere at the same level. Total infarct volume of damaged tissue, expressed in cubic millimeters, was calculated by linear integration of the corrected lesion areas (Golanov and Reis, 1995).
Neurological deficits (six-point neurological severity score).
Neurological deficits of each mouse were evaluated as described previously (Gibson et al., 2011). An expanded 6-point neurologic severity score with the following scale was used: 0, normal function without deficits; 1, flexion and torsion of the contralateral forelimb when held by tail; 2, circling to the ipsilateral side, but normal posture at rest; 3, circling to the ipsilateral side; 4, rolling to the ipsilateral side; and 5, leaning to the ipsilateral side without spontaneous motor activity. The mean of each group was calculated.
Immunofluorescence analysis of neurons and microglia.
After TTC staining, coronal brain slices (+1 mm, bregma and −1 mm) were selected. Brain sections were fixed with 4% PFA and cryoprotected in a series of 10%, 20%, and 30% sucrose for 24 h per solution at 4°C. Tissues were frozen in optimal cutting temperature medium (Sakura-Fineteck) and stored at −20°C. The frozen tissues were cut on the cryostat (HM 560; Thermo Fisher Scientific) to obtain serial coronal brain sections (14 μm). The cryostat sections were washed with PBS 1× and blocked in 3% NGS for 20 min at room temperature and then incubated with the primary antibodies: rabbit anti-NeuN polyclonal antibody (1/500; Millipore) for the staining of neuronal nuclei and rabbit anti-Iba1 antibody (1/100; WAKO), a marker for detecting microglia/macrophages, overnight at 4°C. After rinsing in PBS 1×, sections were incubated with secondary antibody (goat anti-rabbit) diluted 1:200 in PBS 1× for 2 h. After 3 consecutive washes in PBS 1×, the sections were stained for 15 min in 0.1 μg/ml DAPI. The slices were washed in PBS 1× 3 times (5 min each), dried, and coverslipped using Fluormount G (Southern Biotechnology). To show the specificity of the immunostaining, negative controls were processed using 3% NGS in place of primary antibody. Immunofluorescence labeling was visualized using a Zeiss Axioimager microscope A1.
Photographs were obtained with objective (20×) in the cortex and subcortex. Two sections/mouse were analyzed from each group. NeuN+ cells and Iba1+ cells per area of 0.38 mm2 per section were counted. For each mouse, the total number of immune-positive cells in the 2 sections corresponding to an area of 0.76 mm2 was obtained. Data are expressed as means ± SEM of NeuN+ or Iba1+ cells/mm2.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 7 software. The data are expressed as mean ± SEM and n refers to the number of animals per experimental group. Statistical tests included one-way ANOVA and two-way ANOVA followed by Newman–Keuls multiple-comparisons tests between groups. Kruskal–Wallis analysis and Mann–Whitney tests were used for neurological deficits. The correlation of infarct volume and neurological deficits was analyzed by Spearman's correlation analysis. Statistical significance was set at p < 0.05.
Results
Steroid profiling in brain and plasma of young adult male and female mice
We first established steroid profiles by GC-MS/MS for brain and plasma of 3-month-old male and female C57BL/6 mice to determine reference values (Fig. 2, Table 3). Females were at their diestrus stage according to vaginal cytology. Results for pregnenolone and progesterone and its major metabolites are shown in Figure 2. Brain levels of pregnenolone were similar in both sexes and ∼30 times higher than in plasma, consistent with a de novo synthesis by neural cells (Fig. 2A). Levels of progesterone did not differ between sexes in brain, but tended to be higher in the plasma of females. However, in comparison with pregnenolone, brain levels of progesterone were low (<1 ng/g; Fig. 2B). Progesterone is indeed rapidly metabolized in the brain and its measure thus does not provide sufficient information about the dynamics of pregnanes.
Figure 2.
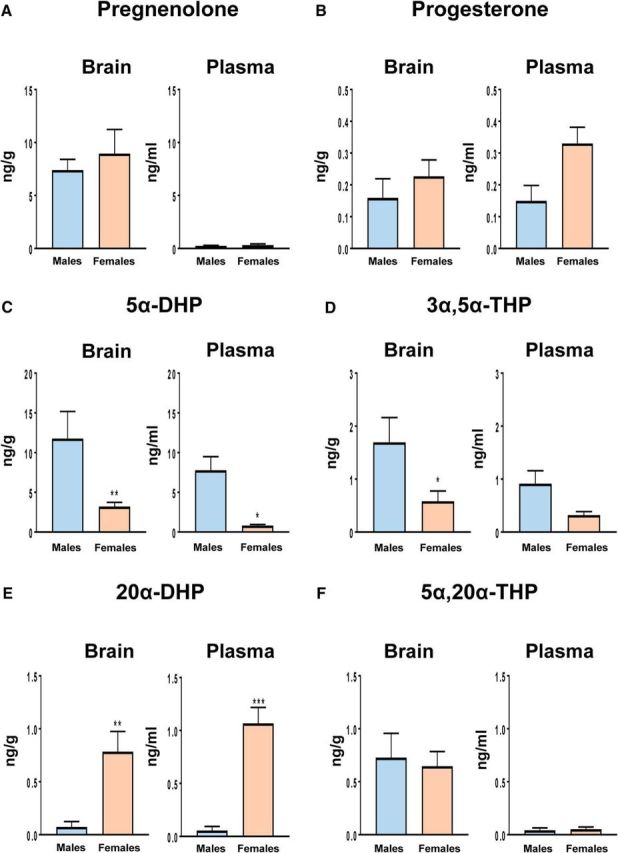
Levels of pregnenolone and progesterone and its major metabolites were analyzed by GC-MS/MS in the left brain hemisphere and plasma of young adult males and diestrus females (for an extended steroid profiling, see Table 3). A, Levels of pregnenolone were higher in the brains of males and females than in plasma (effect of tissue: F(1,29) = 27.5, p < 0.0001). B, Levels of progesterone were similar in brain and plasma of both sexes (effect of tissue: p = 0.4). In plasma, levels of progesterone were higher in females than in males (effect of sex: F(1,25) = 4.74, p < 0.05). C, Levels of 5α-DHP were higher in males than in females for brain and plasma (effect of sex: F(1,27) = 20.6, p < 0.001). Levels of 5α-DHP tended to be higher in brain compared with plasma (effect of tissue: F(1,27) = 3.51, p = 0.072). D, Brain and plasma levels of 3α,5α-THP were higher in males compared with females (effect of sex: F(1,27) = 8.71, p < 0.01). Moreover, levels of 3α,5α-THP tended to be higher in brain compared with plasma (effect of tissue: F(1,27) = 3.26, p = 0.082). E, In contrast to the 5α-reduced 20-keto-pregnanes, levels of 20α-DHP were higher in brain and plasma of females (effect of sex: F(1,25) = 33.1, p < 0.0001). Levels of 20α-DHP did not differ between brain and plasma. F, In the brain, 20α-DHP was converted to 5α,20α-THP. In plasma, levels of 5α,20α-THP were very low (40–50 pg/ml; effect of tissue: F(1,24) = 19.0, p < 0.001). Results are presented as means ± SEM (n = 5–6 per group) and were analyzed by two-way ANOVA (sex × tissue) followed by Newman–Keuls multiple-comparisons tests. ***p < 0.001, **p < 0.01, *p < 0.05 compared with males or as indicated.
Table 3.
Steroid profiling in brain (left hemisphere) and plasma of young adult C57BL6 mice of both sexes
| Steroid | Left hemisphere (ng/g) |
Plasma (ng/ml) |
||
|---|---|---|---|---|
| Males | Females | Males | Females | |
| 5β-DHP | 0.09 ± 0.08 | 0.14 ± 0.08 | 0.02 ± 0.01 | 0.02 ± 0.01 |
| 3β5α-THP | 0.36 ± 0.11 | 0.09 ± 0.04** | 0.01 ± 0.003 | 0.02 ± 0.004 |
| 3α5β-THP | 0.09 ± 0.03 | 0.12 ± 0.07 | 0.07 ± 0.02 | 0.33 ± 0.07* |
| 3α5α20α-HHP | 0.39 ± 0.15 | 0.68 ± 0.15 | 0.18 ± 0.10 | 0.32 ± 0.07 |
| 3β5α20α-HHP | 0.03 ± 0.02 | 0.04 ± 0.02 | 0.01 ± 0.003 | 0.04 ± 0.01 |
| 3α5β20α-HHP | 0.01 ± 0.003 | 0.07 ± 0.04 | 0.03 ± 0.02 | 0.06 ± 0.02 |
| 20α-DHPREG | 0.13 ± 0.02 | 0.14 ± 0.03 | 0.01 ± 0.005 | 0.02 ± 0.01 |
| DOC | 0.02 ± 0.01 | 0.11 ± 0.03 | 0.28 ± 0.13 | 0.20 ± 0.07 |
| 5α-DHDOC | 0.23 ± 0.04 | 0.06 ± 0.01** | 0.20 ± 0.06 | 0.09 ± 0.03* |
| 5β-DHDOC | 0.01 ± 0.01 | 0.21 ± 0.08 | 0.02 ± 0.0002 | 0.21 ± 0.07 |
| 3α5α-THDOC | 0.11 ± 0.03 | 0.02 ± 0.01* | 0.11 ± 0.05 | 0.03 ± 0.01 |
| 3α5β-THDOC | 0.01 ± 0.01 | 0.15 ± 0.06 | 0.01 ± 0.01 | 0.39 ± 0.17 |
| Corticosterone | 3.76 ± 1.00 | 1.40 ± 0.33 | 10.2 ± 4.71 | 21.5 ± 5.99* |
| Testosterone | 1.21 ± 0.39 | 0.02 ± 0.01 | 2.16 ± 0.87 | 0.01 ± 0.003* |
| 5α-DHT | 0.19 ± 0.06 | 0.02 ± 0.01** | 0.08 ± 0.02 | 0.01 ± 0.002 |
| 3α5α-THT | 0.16 ± 0.05 | ND | 0.25 ± 0.06 | ND |
| Estradiol | 0.04 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.03 ± 0.01 |
Results are presented as means ± SEM (n = 6–10 per group) and were analyzed by two-way ANOVA (tissue × sex) followed by Newman–Keuls multiple-comparisons tests.
**p < 0.01,
*p < 0.05 compared with males.
Abbreviations of steroids are as for Table 1. ND, Not detected.
The analysis of progesterone metabolites by GC-MS/MS indeed provided a completely different and unexpected picture, with marked differences between brain and plasma and between sexes. In the male brain, levels of 5α-DHP were 75 times higher than those of progesterone, reaching concentrations of 12 ng/g (∼40 nm; Fig. 2C). Importantly, brain levels of 5α-DHP were ∼4 times higher in males than in females and may thus be considered a major male brain steroid. However, plasma levels of 5α-DHP were also elevated in males, suggesting an important contribution of the adrenal glands (Fig. 2C).
Consistently, brain levels of the 5α-DHP metabolite 3α,5α-THP were higher in males than in females (Fig. 2D). Interestingly, brain levels of 3α,5α-tetrahydrodeoxycorticosterone (3α,5α-THDOC), another potent GABAA receptor modulator derived from the metabolism of adrenal DOC (Sarkar et al., 2011), were also significantly higher in brain of males compared with females (Table 3).
Therefore, steroid profiling by GC-MS/MS revealed the surprising fact that the male mouse brain contains more neuroactive metabolites of progesterone than the female brain. In contrast, brain and plasma levels of 20α-DHP were, respectively, >10 or 20 times higher in females compared with males (Fig. 2E). The reversible conversion of progesterone to 20α-DHP may represent a reservoir of brain pregnanes in females. Additional steroids analyzed in brain and plasma of both sexes are shown in Table 3. Compared with the important 5α-reduction of progesterone in males, the formation of 5α-reduced metabolites of testosterone was very low (Table 3).
Steroid profiling after MCAO in brain and plasma of male and female mice
To gain insight into changes of steroid levels in response to ischemic injury, we performed steroid profiling by GC-MS/MS in the left ischemic hemisphere and in plasma of male and diestrus female C57BL/6 mice at 1, 2, 4, 6, or 24 h after transient MCAO. The left middle cerebral artery was occluded for 30 min, followed by reperfusion (Fig. 3, Table 4).
Figure 3.
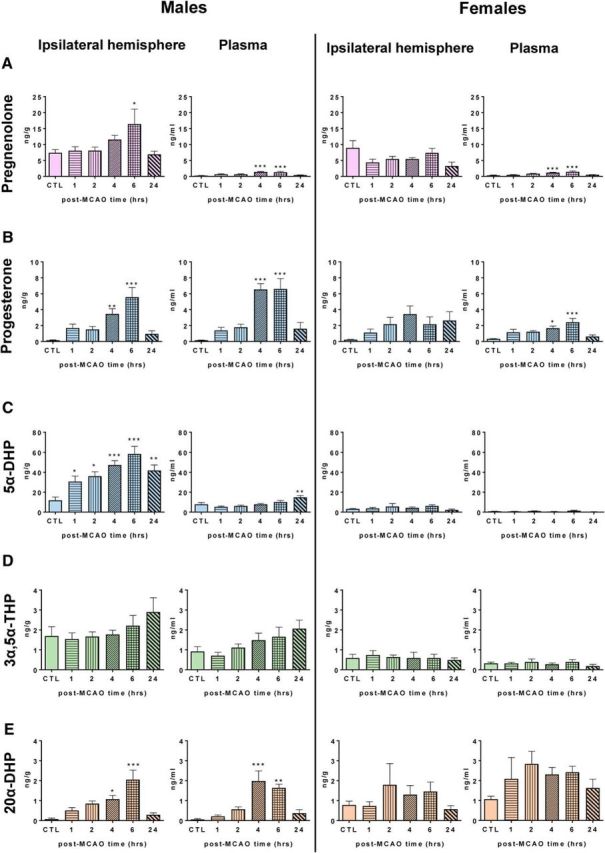
Levels of pregnenolone and progesterone and its major metabolites were analyzed by GC-MS/MS in the ipsilateral (left, ischemic) hemisphere and in plasma of young adult male and diestrus female mice at different times (hours) after MCAO (for an extended steroid profiling, see Table 4). A, In males, levels of pregnenolone were increased significantly in the ipsilateral hemisphere at 6 h after MCAO (F(5,46) = 3.0, p < 0.05) and slightly in plasma at 4 and 6 h (F(5,37) = 8.3, p < 0.001). As in males, plasma levels of pregnenolone were weakly but significantly increased in females at 4 and 6 h (F(5,28) = 10.7, p < 0.001). Analysis by 2-way ANOVA (sex × time) indeed revealed a significant effect of sex (F(5,81) = 11.7, p < 0.001). B, In males, progesterone was significantly increased at 4 and 6 h after MCAO both in the ipsilateral hemisphere (F(5,46) = 9.67, p < 0.0001) and in plasma (F(5,35) = 11.4, p < 0.0001). Analysis by 2-way ANOVA (sex × time) of brain progesterone levels showed a significant effect of time (F(5,80) = 7.94, p < 0.0001) and a significant interaction between time and sex (F(5,80) = 3.20, p < 0.05). Similarly, 2-way ANOVA analysis of plasma progesterone levels revealed a significant effect of time (F(5,60) = 12.4, p < 0.0001), a significant interaction between time and sex (F(5,60) = 4.64, p < 0.001), and also an effect of sex (F(1,60) = 17.8, p < 0.0001). C, In the male ipsilateral hemisphere, levels of 5α-DHP increased continuously between 1 and 6 h after MCAO, and they were still elevated at 24 h (F(5,45) = 7.33 p < 0.0001). Analysis by 2-way ANOVA (sex × time) of brain 5α-DHP levels showed a significant effect of time (F(5,79) = 7.05, p < 0.0001), of sex (F(1,79) = 158.7, p < 0.0001) and a significant interaction between time and sex (F(5,79) = 5.80, p < 0.0001). D, Levels of 3α,5α-THP in brain and plasma were higher in males compared with females. This observation was supported by 2-way ANOVA (sex × time) for brain (effect of sex: F(1,80) = 33.7, p < 0.0001) and plasma (effect of sex: F(1,62) = 29.1, p < 0.0001). E, In males, levels of 20α-DHP were increased significantly in brain (F(5,42) = 9.0, p < 0.0001) and in plasma (F(5,34) = 9.60, p < 0.0001) at 4 and 6 h after MCAO. No significant changes over time were observed in females. A two-way ANOVA (sex × time) revealed a significant effect of sex for plasma levels of 20α-DHP (F(1,61) = 30.5, p < 0.0001). Results are presented as means ± SEM (n = 5–10 per group) and were analyzed by one-way ANOVA followed by Newman–Keuls multiple-comparisons tests. ***p < 0.001, **p < 0.01, *p < 0.05 compared with the control (CTL).
Table 4.
Steroid profiling in brain (ipsilateral hemisphere) and plasma of young adult C57BL/6 mice of both sexes: endpoint measures are at 1, 2, 4, 6, and 24 h after MCAO
| Steroid | Sex | Time post-MCAO (h) |
|||||
|---|---|---|---|---|---|---|---|
| CTL | 1 | 2 | 4 | 6 | 24 | ||
| Ipsilateral hemisphere (ng/g) | |||||||
| 5β-DHP | M | 0.09 ± 0.08 | 0.07 ± 0.01 | 0.09 ± 0.03 | 0.09 ± 0.04 | 0.11 ± 0.04 | 0.06 ± 0.01 |
| F | 0.14 ± 0.08 | 0.04 ± 0.02 | 0.04 ± 0.01 | 0.04 ± 0.02 | 0.11 ± 0.03 | 0.03 ± 0.01 | |
| 3β5α-THP | M | 0.36 ± 0.11 | 0.44 ± 0.08 | 0.41 ± 0.05 | 0.33 ± 0.06 | 0.43 ± 0.10 | 0.61 ± 0.14 |
| F | 0.09 ± 0.04 | 0.01 ± 0.004 | 0.09 ± 0.05 | 0.07 ± 0.02 | 0.09 ± 0.04 | 0.14 ± 0.02$ | |
| 3α5β-THP | M | 0.09 ± 0.03 | 0.06 ± 0.01 | 0.08 ± 0.03 | 0.10 ± 0.04 | 0.10 ± 0.03 | 0.06 ± 0.01 |
| F | 0.12 ± 0.07 | 0.10 ± 0.03 | 0.19 ± 0.07 | 0.47 ± 0.22 | 0.71 ± 0.32$ | 0.47 ± 0.26 | |
| 3α5α20α-HHP | M | 0.39 ± 0.15 | 0.98 ± 0.34 | 0.51 ± 0.07 | 0.46 ± 0.07 | 0.69 ± 0.17 | 0.66 ± 0.17 |
| F | 0.68 ± 0.15 | 0.36 ± 0.11 | 0.62 ± 0.25 | 0.28 ± 0.10 | 0.21 ± 0.04 | 0.14 ± 0.05 | |
| 3β5α20α-HHP | M | 0.03 ± 0.02 | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.05 ± 0.02 | 0.04 ± 0.01 |
| F | 0.04 ± 0.02 | 0.05 ± 0.02 | 0.04 ± 0.02 | 0.05 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | |
| 3α5β20α-HHP | M | 0.01 ± 0.003 | 0.005 ± 0.002 | ND | ND | ND | ND |
| F | 0.07 ± 0.04 | 0.01 ± 0.002 | 0.02 ± 0.004 | 0.03 ± 0.01 | 0.10 ± 0.05 | 0.001 ± 0.00005 | |
| 20α-DHPREG | M | 0.13 ± 0.02 | 0.09 ± 0.03 | 0.09 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.02 |
| F | 0.14 ± 0.03 | 0.18 ± 0.03 | 0.21 ± 0.08 | 0.59 ± 0.50$$ | 0.21 ± 0.05 | 0.11 ± 0.03 | |
| DOC | M | 0.02 ± 0.01 | 2.68 ± 0.69 | 4.56 ± 0.48* | 7.90 ± 1.56*** | 7.30 ± 1.57*** | 1.34 ± 0.40 |
| F | 0.11 ± 0.03 | 4.47 ± 1.49** | 3.27 ± 0.51 | 4.79 ± 0.71* | 5.93 ± 1.41*** | 0.81 ± 0.27 | |
| 5α-DHDOC | M | 0.23 ± 0.04 | 1.16 ± 0.25 | 1.20 ± 0.18 | 1.96 ± 0.30* | 1.90 ± 0.61* | 1.48 ± 0.33 |
| F | 0.06 ± 0.01 | 0.49 ± 0.18 | 0.75 ± 0.23* | 0.99 ± 0.26** | 0.90 ± 0.15** | 0.35 ± 0.10 | |
| 5β-DHDOC | M | 0.01 ± 0.01 | 0.01 ± 0.001 | 0.01 ± 0.002 | 0.07 ± 0.03 | 0.15 ± 0.09 | 0.06 ± 0.03 |
| F | 0.21 ± 0.08 | 2.63 ± 1.21*$$ | 3.46 ± 0.66**$$$ | 2.43 ± 0.27$$ | 3.45 ± 0.70**$$$ | 0.95 ± 0.33 | |
| 3α5α-THDOC | M | 0.11 ± 0.03 | 0.37 ± 0.01 | 0.33 ± 0.07 | 0.35 ± 0.06 | 0.39 ± 0.14 | 0.61 ± 0.19 |
| F | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.13 ± 0.02*** | 0.01 ± 0.002$$ | |
| 3α5β-THDOC | M | 0.01 ± 0.01 | 0.03 ± 0.01 | 0.003 ± 0.0003 | 0.05 ± 0.02 | 0.08 ± 0.05 | 0.02 ± 0.01 |
| F | 0.15 ± 0.06 | 0.99 ± 0.84 | 1.44 ± 0.78 | 0.75 ± 0.41 | 7.71 ± 3.36*$$$ | 0.58 ± 0.54 | |
| Corticosterone | M | 3.76 ± 1.00 | 27.6 ± 6.24 | 69.0 ± 21.8 | 75.7 ± 23.3 | 214.6 ± 41.2*** | 67.0 ± 23.5 |
| F | 1.40 ± 0.33 | 17. 0 ± 6.85 | 49.2 ± 13.6** | 76.1 ± 20.2*** | 68.6 ± 10.6***$$$ | 20.4 ± 8.61 | |
| Testosterone | M | 1.21 ± 0.39 | 0.22 ± 0.06*** | 0.15 ± 0.05*** | 0.14 ± 0.03*** | 0.08 ± 0.01*** | 0.06 ± 0.01*** |
| F | 0.02 ± 0.01$$$ | 0.04 ± 0.02 | 0.01 ± 0.01 | 0.03 ± 0.03 | 0.02 ± 0.005 | 0.01 ± 0.01 | |
| 5α-DHT | M | 0.19 ± 0.06 | 0.06 ± 0.02*** | 0.02 ± 0.01*** | 0.03 ± 0.01*** | 0.02 ± 0.01*** | 0.02 ± 0.001*** |
| F | 0.02 ± 0.01$$$ | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.002 ± 0.001 | 0.01 ± 0.004 | 0.002 ± 0.001 | |
| 3α5α-THT | M | 0.16 ± 0.05 | 0.05 ± 0.01*** | 0.02 ± 0.02*** | ND | 0.003 ± 0.002*** | 0.01 ± 0.01*** |
| F | ND | ND | ND | ND | ND | ND | |
| Estradiol | M | 0.04 ± 0.01 | 0.05 ± 0.02 | 0.04 ± 0.02 | 0.06 ± 0.01 | 0.04 ± 0.02 | 0.02 ± 0.01 |
| F | 0.02 ± 0.01 | 0.01 ± 0.002 | 0.02 ± 0.01 | 0.01 ± 0.01 | 0.001 ± 0.0002 | 0.02 ± 0.01 | |
| Plasma (ng/ml) | |||||||
| 5β-DHP | M | 0.02 ± 0.01 | ND | ND | ND | ND | ND |
| F | 0.02 ± 0.01 | 0.12 ± 0.01***$$$ | 0.12 ± 0.01***$$$ | 0.11 ± 0.02***$$$ | 0.17 ± 0.03***$$$ | 0.09 ± 0.02**$$$ | |
| 3β5α-THP | M | 0.01 ± 0.003 | 0.03 ± 0.01 | 0.05 ± 0.02 | 0.04 ± 0.01 | 0.07 ± 0.02 | 0.04 ± 0.02 |
| F | 0.02 ± 0.004 | 0.01 ± 0.002 | 0.02 ± 0.002 | 0.01 ± 0.005 | 0.02 ± 0.01 | 0.02 ± 0.01 | |
| 3α5β-THP | M | 0.07 ± 0.02 | 0.10 ± 0.03 | 0.07 ± 0.02 | 0.06 ± 0.01 | 0.10 ± 0.02 | 0.09 ± 0.04 |
| F | 0.33 ± 0.07 | 0.22 ± 0.09 | 0.57 ± 0.06 | 0.74 ± 0.09$$ | 1.03 ± 0.38*$$$ | 0.25 ± 0.09 | |
| 3α5α20α-HHP | M | 0.18 ± 0.10 | 0.14 ± 0.05 | 0.35 ± 0.05 | 0.54 ± 0.11 | 0.52 ± 0.12 | 0.34 ± 0.07 |
| F | 0.32 ± 0.07 | 0.26 ± 0.03 | 0.54 ± 0.11 | 0.35 ± 0.03 | 0.27 ± 0.07 | 0.22 ± 0.04 | |
| 3β5α20α-HHP | M | 0.01 ± 0.003 | 0.02 ± 0.004 | 0.04 ± 0.02 | 0.05 ± 0.02 | 0.05 ± 0.02 | 0.06 ± 0.01 |
| F | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.10 ± 0.03 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | |
| 3α5β20α-HHP | M | 0.03 ± 0.02 | 0.01 ± 0.003 | 0.004 ± 0.002 | 0.02 ± 0.01 | 0.01 ± 0.005 | 0.01 ± 0.004 |
| F | 0.06 ± 0.02 | 0.07 ± 0.02 | 0.15 ± 0.04$$ | 0.23 ± 0.02**$$$ | 0.25 ± 0.06**$$$ | 0.07 ± 0.02 | |
| 20α-DHPREG | M | 0.01 ± 0.005 | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.03 ± 0.002 | 0.02 ± 0.004 | 0.02 ± 0.01 |
| F | 0.02 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.03 ± 0.002 | 0.02 ± 0.01 | 0.02 ± 0.01 | |
| DOC | M | 0.28 ± 0.13 | 1.42 ± 0.36 | 2.27 ± 0.68 | 2.78 ± 0.63 | 4.83 ± 1.04*** | 0.84 ± 0.47 |
| F | 0.20 ± 0.07 | 1.31 ± 0.45 | 2.07 ± 0.52 | 4.08 ± 0.93*** | 3.26 ± 0.66*** | 0.81 ± 0.27 | |
| 5α-DHDOC | M | 0.20 ± 0.06 | 0.35 ± 0.09 | 0.34 ± 0.15 | 0.50 ± 0.14 | 0.39 ± 0.16 | 0.43 ± 0.13 |
| F | 0.09 ± 0.03 | 0.04 ± 0.02 | 0.05 ± 0.02 | 0.18 ± 0.02 | 0.18 ± 0.06 | 0.08 ± 0.02 | |
| 5β-DHDOC | M | 0.02 ± 0.0002 | 0.03 ± 0.01 | 0.02 ± 0.003 | 0.03 ± 0.02 | 0.01 ± 0.002 | 0.06 ± 0.03 |
| F | 0.21 ± 0.07 | 1.64 ± 0.21**$$$ | 1.79 ± 0.19**$$$ | 2.89 ± 0.17***$$$ | 2.54 ± 0.47***$$$ | 1.01 ± 0.39*$$$ | |
| 3α5α-THDOC | M | 0.11 ± 0.05 | 0.26 ± 0.09 | 0.39 ± 0.22 | 0.51 ± 0.23 | 0.38 ± 0.20 | 0.41 ± 0.19 |
| F | 0.02 ± 0.01 | 0.05 ± 0.01 | 0.09 ± 0.03 | 0.24 ± 0.07*** | 0.17 ± 0.03* | 0.05 ± 0.01 | |
| 3α5β-THDOC | M | 0.01 ± 0.01 | 0.04 ± 0.01 | 0.07 ± 0.06 | 0.10 ± 0.03 | 0.03 ± 0.01 | 0.05 ± 0.03 |
| F | 0.39 ± 0.17 | 6.95 ± 1.43 | 13.9 ± 0.94*$$ | 29.6 ± 3.11***$$$ | 21.8 ± 7.52***$$$ | 4.58 ± 2.10 | |
| Corticosterone | M | 10.2 ± 4.71 | 109.2 ± 13.9** | 139.3 ± 21.9*** | 215.6 ± 33.6*** | 213.2 ± 12.2*** | 67.6 ± 18.1* |
| F | 21.5 ± 5.99 | 60.5 ± 6.80 | 92.0 ± 4.13 | 182.4 ± 41.3** | 191.2 ± 42.2** | 66.4 ± 31.6 | |
| Testosterone | M | 2.16 ± 0.87 | 0.34 ± 0.07*** | 0.15 ± 0.04*** | 0.14 ± 0.02*** | 0.11 ± 0.02*** | 0.05 ± 0.01*** |
| F | 0.01 ± 0.003$$$ | 0.02 ± 0.01 | 0.01 ± 0.01 | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.003 | |
| 5α-DHT | M | 0.08 ± 0.02 | 0.02 ± 0.01 | 0.04 ± 0.03 | 0.06 ± 0.03 | 0.06 ± 0.03 | 0.08 ± 0.04 |
| F | 0.01 ± 0.002 | ND | 0.02 ± 0.01 | 0.004 ± 0.002 | 0.01 ± 0.004 | 0.004 ± 0.003 | |
| 3α5α-THT | M | 0.25 ± 0.06 | 0.17 ± 0.07 | 0.06 ± 0.02* | 0.05 ± 0.003* | 0.03 ± 0.01** | ND |
| F | ND | ND | ND | ND | ND | ND | |
| Estradiol | M | 0.02 ± 0.01 | 0.02 ± 0.003 | 0.03 ± 0.002 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 |
| F | 0.03 ± 0.01 | 0.08 ± 0.03 | 0.02 ± 0.01 | 0.06 ± 0.02 | 0.01 ± 0.01 | 0.04 ± 0.02 | |
Plasma and brain levels of steroids are expressed as ng/ml for plasma or ng/g for brain (mean ± SEM; n = 4–10 per group). Statistical analysis: one way ANOVA followed by Newman–Keuls multiple-comparisons tests.
***p < 0.001,
**p < 0.01,
*p < 0.05 versus CTL (controls) without MCAO and by two-way ANOVA (sex × time) followed by Newman–Keuls multiple-comparisons tests. Significance: $$$p < 0.001, $$p < 0.01, $p < 0.05 females versus males.
Abbreviations of steroids are as for Table 1. ND, Not detected.
In both males and females, brain levels of pregnenolone were ∼5–10 times higher in brain compared with plasma, confirming results of the previous experiment (Fig. 3A). In the ipsilateral hemisphere, levels of pregnenolone were increased significantly at 6 h after MCAO and returned to control values after 24 h in males. There was no such change with time in females. In both sexes, the low plasma levels of pregnenolone were slightly, but significantly, increased in a transient manner at 4 and 6 h (Fig. 3A).
In males, brain levels of progesterone were increased at 4 and 6 h after MCAO, paralleled by a similar increase in plasma levels, which probably reflects activation of the adrenal glands (Fig. 3B). A similar trend was observed in females, but without reaching statistical significance for the brain. At 6 h, brain and plasma levels of progesterone were lower in females compared with males.
The magnitude of brain-specific changes in endogenous progesterone after MCAO may have been partially masked by its rapid and strong 5α-reduction (Poletti et al., 1998). Indeed, the most marked differences between tissues and sexes were observed for 5α-DHP. In the male ipsilateral hemisphere, levels of 5α-DHP raised continuously between 1 and 6 h after MCAO (Fig. 3C). Maximal and very elevated brain levels of 5α-DHP were reached at 6 h. In plasma, levels of 5α-DHP remained much lower, with a slight increase at 24 h. Levels of 5α-DHP remained elevated in the male brain as late as 24 h after MCAO. In contrast, levels of 5α-DHP remained low in brain (≤5 ng/g) and plasma (≤1 ng/ml) of females (Fig. 3C).
The GABAA receptor active progesterone metabolite 3α,5α-THP did not show significant changes with time after MCAO in both sexes either in brain or in plasma. However, levels of 3α,5α-THP in brain and plasma were higher in males compared with females (Fig. 3D). In males, levels of 20α-DHP were increased significantly in brain and plasma at 4 and 6 h after MCAO. No significant changes over time for the constantly elevated levels of 20α-DHP in females (Fig. 3E).
Summing the levels of progesterone and 5α-DHP well illustrates the rapid and marked increase in PR-active steroids in male brain after MCAO and the noticeable sex difference. Both pregnanes reached maximal levels of nearly 70 ng/g (∼220 nm) at 6 h after MCAO in the male brain (Fig. 4A), which is comparable to their pregnancy plasma levels in mice (Murr et al., 1974). In males, plasma levels of progesterone + 5α-DHP were ∼6 times lower than brain levels, and only showed a small increase at 6 and 24 h (Fig. 4B). In contrast to males, summed levels of progesterone + 5α-DHP remained low after MCAO in the female brain and plasma (Fig. 4C,D).
Figure 4.

Rapid and marked increase in the summed levels of the PR-active steroids progesterone and 5α-DHP after MCAO in the ipsilateral hemisphere of males. A, In the male brain, progesterone + 5α-DHP increased progressively between 1 and 6 h after MCAO and remained elevated at 24 h (F(5,45) = 7.87, p < 0.0001). B, In the plasma of males, pooled levels of progesterone + 5α-DHP remained low, but showed a small increase over time (F(5,37) = 6.48, p < 0.001). C, In contrast to males, levels of progesterone + 5α-DHP remained very low in the female brain and were not affected by MCAO. D, There was a slight, albeit significant increase in female plasma levels of progesterone + 5α-DHP at 4 and 6 h (F(5,28) = 13.1, p < 0.0001).
Additional steroid metabolomic data for brain and plasma, providing reference data, are presented in Table 4. In contrast to 5α-DHP, levels of 5β-DHP were very low in brain and plasma of both sexes (∼10 pg per g or ml). Because the molecular structure of 5β-reduced steroids is not planar but bent, they are considered as biologically inactive and to be involved in hormone clearance (Chen and Penning, 2014). However, some 5β-reduced metabolites of progesterone have neuroactive properties. Therefore, 3α,5β-tetrahydroprogesterone (3α,5β-THP, pregnanolone) is like 3α,5α-THP (allopregnanolone) a positive allosteric modulator of GABAA receptors, both having the same potency (Belelli et al., 1996). Interestingly, whereas brain and plasma levels of 3α,5α-THP were higher in males (Fig. 3D), those of 3α,5β-THP were higher in females at 6 h after MCAO (Table 4). Levels of 3α,5β,20α-hexahydroprogesterone were also higher in females than in males, but the biological significance of the hexahydroprogesterone isomers remains to be explored.
Remarkable were sex differences in the levels of neuroactive steroids derived from the adrenal glands. Plasma levels of corticosterone, the principal glucocorticoid in mice and rats, increased from 10 to 20 ng/ml in controls to ∼200 ng/mg at 6 h after MCAO in both sexes (effect of time: F(5,47) = 18.3, p < 0.0001; Table 4). However, in the brain, a different picture was observed. Although levels of corticosterone increased significantly in the ischemic hemisphere of both sexes (effect of time: F(5,76) = 13.2, p < 0.0001), they remained lower than in plasma until 4 h after MCAO. Only at 6 h did brain levels of corticosterone reach plasma levels in males, whereas in females, brain levels of corticosterone remained lower (effect of sex: F(1,76) = 10.5, p = 0.002; interaction between time and sex (F(5,76) = 4.31, p = 0.002). The female brain may thus protect itself against the steadily increasing, very high levels of adrenal glucocorticoids. After 24 h, levels of the adrenal steroids returned to lower levels but remained above control levels.
In the plasma, conversion of the corticosterone precursor DOC to its 5β-reduced metabolites 5β-DHDOC (effect of sex: F(1,59) = 244.7, p < 0.0001) and 3α,5β-THDOC (effect of sex: F(1,50) = 80.7, p < 0.0001) was much higher in females than in males (Table 4). In the circulation, the sum of both 5β-reduced metabolites reached 32 ng/ml in females at 4 h, which were ∼250 times the levels measured in males. In the brain, levels of both metabolites were also higher in females than in males (5β-DHDOC: effect of sex: F(1,77) = 83.5, p < 0.0001; 3α,5β-THDOC: effect of sex: F(1,67) = 6.69, p < 0.05; Table 4). Therefore, females produce significantly higher amounts of GABAA receptor active 5β-reduced metabolites of progesterone and DOC, respectively, 3α,5β-THP, and, by structural analogy, 3α,5β-THDOC (Gunn et al., 2015).
In contrast to the glucocorticoids, levels of testosterone decreased after the ischemic injury in plasma (effect of time: F(5,59) = 5.38, p < 0.001) and brain (effect of time: F(5,73) = 5.89, p < 0.001) of males (Table 4). Moreover, there was a significant decrease in male brain 5α-DHT levels (effect of time: F(5,67) = 5.41, p < 0.001). This decrease in the 5α-reduction of testosterone thus differs from the marked increase in the formation of 5α-reduced metabolites of progesterone in males (Fig. 3C,D). As expected (Nilsson et al., 2015), low brain concentrations of estradiol were measured and no significant changes over time were observed after MCAO.
Selective deletion of intracellular PRs in the CNS
We hypothesized that the rapid and important increase in brain levels of progesterone and 5α-DHP may be part of spontaneous cerebroprotective responses. To gain an insight into the potential role of these endogenous PR ligands in the acute resistance of brain tissues against ischemic damage, we inactivated PR expression selectively in neural cells using the Cre-LoxP site-specific recombination system to generate a new transgenic mice line PRNesCre.
Specificity and efficiency of neural PR gene invalidation via Cre recombination were determined by quantitative RT-PCR (qPCR) and immunofluorescence analyses. qPCR showed a nearly complete deletion of PR mRNA expression in the brain of PRNesCre males and females (96% in hypothalamus and 99% in cortex and subcortical regions; Fig. 5A). In contrast to the brain, PR mRNA expression was similar in wild-type PR+/+, PRloxP/loxP, and PRNesCre mice for aorta, kidney, and thymus in males and for uterus, ovaries, and mammary glands in females (Fig. 5B). Immunofluorescence detection revealed that neural PR protein was expressed in the hypothalamus of PRloxP/loxP mice, but was almost completely absent in the hypothalamus of PRNesCre mice (Fig. 5C). In contrast to the hypothalamus, a similar distribution and density of PR immunoreactivity was observed in the uterus of PRloxP/loxP and PRNesCre female mice (Fig. 5C). Altogether, these results demonstrate that the PR gene was invalidated efficiently and selectively in the brains of both male and female PRNesCre mice. Because the Cre recombinase was not expressed in microglia, present results also show that, in the mouse brain, PRs are not present in this cell type.
Figure 5.
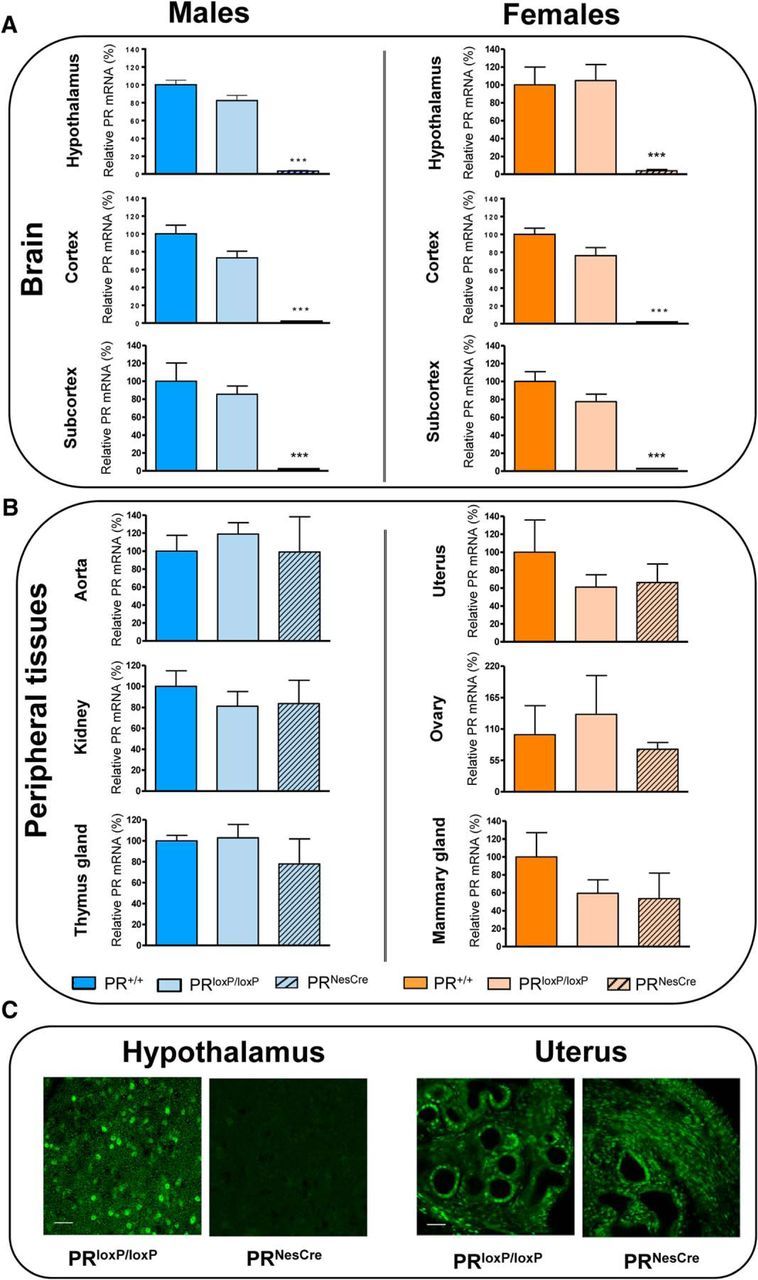
Verification of the selective deletion of the progesterone receptors in the brain of PRNesCre mice. A, B, PR mRNA expression was analyzed by q-PCR (n = 4 per group). A, In hypothalamus, cerebral cortex, and subcortical regions, PR mRNA expression was similar in wild-type (PR+/+) and male and female PRloxP/loxP mice, but was almost completely abolished in PRNesCre mice. B, In contrast to the brain, PR mRNA expression was not affected in peripheral tissues of PRNesCre mice, including aorta, kidney, and thymus for males and uterus, ovaries, and mammary glands for females. Results are presented as means ± SEM. Group differences were analyzed by one-way ANOVA followed by Newman–Keuls multiple-comparisons tests. ***p < 0.001 compared with PRloxP/loxP mice. C, Successful deletion of PR was also verified in the hypothalamus at the protein level by immunofluorescence. No specific immunostaining was detected in PRNesCre mice. In contrast to the hypothalamus, PR immunostaining was not decreased in the uterus of PRNesCre mice compared with PRloxP/loxP mice. Scale bar, 50 μm.
At the age of 1 month, PRNesCre mice had the same weight as sex-matched PRloxP/loxP mice. At 4 months, about the time when experiments of ischemic injury in young animals were performed, PRNesCre mice were 6–7% lighter than PRloxP/loxP mice of the same sex (males: PRNesCre: 26.4 ± 0.2 g; PRloxP/loxP: 28.1 ± 0.2 g; females: PRNesCre: 20.9 ± 0.2 g; PRloxP/loxP: 22.4 ± 0.2 g, means ± SEM; effect of sex: F(1,29) = 1208.1, p < 0.001; effect of genotype: F(1,29) = 100.3, p < 0.001). At 12 months, the weight differences between PRloxP/loxP and PRNesCre mice became even more pronounced, but did not exceed 20% (males: PRNesCre: 28.3 ± 0.3 g; PRloxP/loxP: 35.1 ± 1.1 g; females: PRNesCre: 27.3 ± 0.8 g; PRloxP/loxP: 31.3 ± 1.5 g, means ± SEM; effect of sex: F(1,31) = 5.55, p < 0.05; effect of genotype: F(1,31) = 28.5, p < 0.001). Therefore, during aging, PRNesCre mice of both sexes gain less weight.
Male PRNesCre mice showed normal fertility when mated with female PRloxP/loxP mice. Of eight pairs, all produced a number of offspring comparable to wild-types. In contrast, PRNesCre females exhibited significantly reduced fertility when mated with PRloxP/loxP males. Over a 3-month observation period, only 4 females of 24 produced offspring and the size of their litters did not exceed 2 pups. One reason for this low fertility of PRNesCre females was their irregular estrus cycles as determined by vaginal cytology and a markedly reduced number of corpus lutea within the ovaries of PRNesCre females. This reproductive phenotype is consistent with the previously reported inability of total PR knock-out mice to ovulate (Lydon et al., 1995).
Steroid profiling in brain and plasma of young adult PRloxP/loxP and PRNesCre mice of both sexes
To assess whether conditional inactivation of PR in neural cells of the CNS had an impact on steroid levels, extended brain and plasma profiles were established by GC-MS/MS for young (3 months old) male and female PRloxP/loxP and PRNesCre mice (Table 5). Both PRloxP/loxP and PRNesCre females were again sampled according to vaginal cytology at the diestrus stage.
Table 5.
Steroid profiling in brain (left hemisphere) and plasma of young adult PRloxP/loxP and PRNesCre mice of both sexes
| Steroid | PRloxP/loxP |
PRNesCre |
Two-way ANOVA | ||
|---|---|---|---|---|---|
| Males | Females | Males | Females | ||
| Left hemisphere (ng/g) | |||||
| Pregnenolone | 9.94 ± 0.61 | 6.84 ± 0.08* | 10.3 ± 0.78 | 6.14 ± 0.17* | Sex: F(1,17) = 20.4; p = 0.0003 |
| Genotype: F(1,17) = 0.05; p = 0.825 | |||||
| Interaction: F(1,17) = 0.43; p = 0.521 | |||||
| Progesterone | 0.73 ± 0.08 | 0.44 ± 0.06 | 0.38 ± 0.14 | 0.46 ± 0.11 | Sex: F(1,17) = 1.09; p = 0.311 |
| Genotype: F(1,17) = 2.59; p = 0.126 | |||||
| Interaction: F(1,17) = 3.10; p = 0.096 | |||||
| 5α-DHP | 24.0 ± 5.36 | 5.69 ± 0.95** | 28.3 ± 2.52 | 4.81 ± 0.32** | Sex: F(1,18) = 29.2; p < 0.0001 |
| Genotype: F(1,18) = 0.19; p = 0.669 | |||||
| Interaction: F(1,18) = 0.44; p = 0.515 | |||||
| 3α,5α-THP | 3.57 ± 0.79 | 0.53 ± 0.18*** | 1.46 ± 0.20$$ | 0.15 ± 0.03 | Sex: F(1,18) = 31.6; p < 0.0001 |
| Genotype: F(1,18) = 10.3; p = 0.005 | |||||
| Interaction: F(1,18) = 4.96; p = 0.039 | |||||
| 20α-DHP | 2.12 ± 0.16 | 22.6 ± 6.42** | 0.90 ± 0.28 | 18.5 ± 1.63* | Sex: F(1,16) = 21.9; p = 0.0003 |
| Genotype: F(1,16) = 0.43; p = 0.522 | |||||
| Interaction: F(1,16) = 0.13; p = 0.728 | |||||
| 5α,20α-THP | 1.30 ± 0.20 | 0.64 ± 0.16** | 0.50 ± 0.07$$ | 0.61 ± 0.05 | Sex: F(1,17) = 3.60; p = 0.075 |
| Genotype: F(1,17) = 8.38; p = 0.010 | |||||
| Interaction: F(1,17) = 7.28; p = 0.015 | |||||
| 5β-DHP | ND | 0.28 ± 0.04 | ND | 0.30 ± 0.06 | |
| 3β5α-THP | 0.44 ± 0.10 | 0.01 ± 0.004*** | 0.12 ± 0.02$$$ | 0.01 ± 0.002 | Sex: F(1,17) = 27.4; p < 0.0001 |
| Genotype: F(1,17) = 9.49; p = 0.007 | |||||
| Interaction: F(1,17) = 9.27; p = 0.007 | |||||
| 3α5β-THP | 0.03 ± 0.01 | 0.24 ± 0.05** | 0.10 ± 0.02 | 0.34 ± 0.06*** | Sex: F(1,17) = 32.3; p < 0.0001 |
| Genotype: F(1,17) = 4.55; p = 0.048 | |||||
| Interaction: F(1,17) = 0.30; p = 0.592 | |||||
| 3α5α20α-HHP | 1.29 ± 0.37 | 0.53 ± 0.10* | 0.35 ± 0.05$ | 0.58 ± 0.04 | Sex: F(1,15) = 1.69; p = 0.213 |
| Genotype: F(1,15) = 4.61; p = 0.049 | |||||
| Interaction: F(1,15) = 5.82; p = 0.029 | |||||
| 3β5α20α-HHP | 0.18 ± 0.05 | 0.04 ± 0.01** | 0.04 ± 0.01$$ | 0.03 ± 0.001 | Sex: F(1,15) = 8.63; p = 0.010 |
| Genotype: F(1,15) = 7.17; p = 0.017 | |||||
| Interaction: F(1,15) = 5.07; p = 0.040 | |||||
| 3α5β20α-HHP | 0.01 ± 0.003 | 0.07 ± 0.03* | ND | 0.27 ± 0.01***$$$ | |
| 20α-DHPREG | 0.41 ± 0.02 | 0.30 ± 0.02* | 0.35 ± 0.04 | 0.34 ± 0.01 | Sex: F(1,17) = 5.47; p = 0.032 |
| Genotype: F(1,17) = 0.25; p = 0.625 | |||||
| Interaction: F(1,17) = 4.16; p = 0.057 | |||||
| DOC | 1.78 ± 0.31 | 7.25 ± 1.32*** | 1.11 ± 0.45 | 6.02 ± 0.31** | Sex: F(1,17) = 38.1; p < 0.0001 |
| Genotype: F(1,17) = 1.28; p = 0.273 | |||||
| Interaction: F(1,17) = 0.11; p = 0.743 | |||||
| 5α-DHDOC | 0.87 ± 0.24 | 0.67 ± 0.08 | 0.71 ± 0.12 | 0.83 ± 0.11 | Sex: F(1,17) = 0.08; p = 0.786 |
| Genotype: F(1,17) = 0.0001; p = 0.992 | |||||
| Interaction: F(1,17) = 1.38; p = 0.257 | |||||
| 5β-DHDOC | ND | 4.11 ± 0.77 | ND | 5.21 ± 0.82 | |
| 3α5α-THDOC | 0.75 ± 0.05 | 0.06 ± 0.01*** | 0.33 ± 0.06$$$ | 0.12 ± 0.04** | Sex: F(1,18) = 102.2; p < 0.0001 |
| Genotype: F(1,18) = 16.2; p = 0.0008 | |||||
| Interaction: F(1,18) = 27.5; p < 0.0001 | |||||
| 3α5β-THDOC | ND | 1.00 ± 0.27 | ND | 1.41 ± 0.29 | |
| Corticosterone | 47.6 ± 4.73 | 32.5 ± 4.66 | 19.4 ± 3.05$$$ | 34.2 ± 4.29 | Sex: F(1,18) = 0.001; p = 0.979 |
| Genotype: F(1,18) = 9.86; p = 0.006 | |||||
| Interaction: F(1,18) = 12.6; p = 0.002 | |||||
| Testosterone | 0.09 ± 0.002 | 0.01 ± 0.004* | 0.11 ± 0.03 | 0.01 ± 0.004** | Sex: F(1,14) = 23.7; p = 0.0003 |
| Genotype: F(1,14) = 0.41; p = 0.533 | |||||
| Interaction: F(1,14) = 0.52; p = 0.482 | |||||
| 5α-DHT | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.02 ± 0.01 | Sex: F(1,16) = 2.41; p = 0.141 |
| Genotype: F(1,16) = 0.02; p = 0.901 | |||||
| Interaction: F(1,16) = 0.15; p = 0.704 | |||||
| 3α5α-THT | 0.01 ± 0.003 | 0.002 ± 0.00002** | 0.007 ± 0.004 | 0.002 ± 0.00004 | Sex: F(1,16) = 13.3; p = 0.002 |
| Genotype: F(1,16) = 1.85; p = 0.193 | |||||
| Interaction: F(1,16) = 1.83; p = 0.195 | |||||
| Estradiol | 0.02 ± 0.0003 | 0.01 ± 0.002 | 0.01 ± 0.002 | 0.01 ± 0.001 | Sex: F(1,16) = 0.89; p = 0.361 |
| Genotype: F(1,16) = 2.24; p = 0.154 | |||||
| Interaction: F(1,16) = 3.89; p = 0.066 | |||||
| Plasma (ng/ml) | |||||
| Pregnenolone | 0.35 ± 0.07 | 0.39 ± 0.02 | 0.23 ± 0.03 | 0.36 ± 0.12 | Sex: F(1,17) = 1.54; p = 0.232 |
| Genotype: F(1,17) = 1.33; p = 0.265 | |||||
| Interaction: F(1,17) = 0.51; p = 0.486 | |||||
| Progesterone | 0.52 ± 0.08 | 0.51 ± 0.09 | 0.43 ± 0.17 | 0.33 ± 0.02 | Sex: F(1,18) = 0.34; p = 0.565 |
| Genotype: F(1,18) = 2.46; p = 0.134 | |||||
| Interaction: F(1,18) = 0.29; p = 0.598 | |||||
| 5α-DHP | 9.66 ± 0.61 | 0.57 ± 0.10*** | 8.85 ± 0.66 | 0.67 ± 0.10*** | Sex: F(1,17) = 318.2; p < 0.0001 |
| Genotype: F(1,17) = 0.54; p = 0.471 | |||||
| Interaction: F(1,17) = 0.90; p = 0.357 | |||||
| 3α,5α-THP | 2.29 ± 0.47 | 0.18 ± 0.07*** | 0.89 ± 0.06$$$ | 0.19 ± 0.06 | Sex: F(1,16) = 33.3; p < 0.0001 |
| Genotype: F(1,16) = 8.05; p = 0.012 | |||||
| Interaction: F(1,16) = 8.30; p = 0.011 | |||||
| 20α-DHP | 1.51 ± 0.19 | 19.3 ± 4.35*** | 0.64 ± 0.11 | 22.3 ± 2.01*** | Sex: F(1,16) = 71.3; p < 0.0001 |
| Genotype: F(1,16) = 0.22; p = 0.645 | |||||
| Interaction: F(1,16) = 0.71; p = 0.412 | |||||
| 5α,20α-THP | 0.33 ± 0.10 | 0.17 ± 0.06 | 0.10 ± 0.02 | 0.18 ± 0.01 | Sex: F(1,16) = 0.33; p = 0.575 |
| Genotype: F(1,16) = 3.28; p = 0.089 | |||||
| Interaction: F(1,16) = 3.96; p = 0.064 | |||||
| 5β-DHP | ND | 0.11 ± 0.01 | 0.01 ± 0.002 | 0.10 ± 0.02 | |
| 3β5α-THP | 0.03 ± 0.01 | 0.002 ± 0.001** | 0.005 ± 0.001$$ | 0.004 ± 0.002 | Sex: F(1,17) = 9.21; p = 0.008 |
| Genotype: F(1,17) = 6.24; p = 0.023 | |||||
| Interaction: F(1,17) = 7.49; p = 0.014 | |||||
| 3α5β-THP | 0.01 ± 0.002 | 0.24 ± 0.02*** | 0.01 ± 0.002 | 0.31 ± 0.02***$$ | Sex: F(1,13) = 338.9; p < 0.0001 |
| Genotype: F(1,13) = 5.62; p = 0.034 | |||||
| Interaction: F(1,13) = 5.71; p = 0.033 | |||||
| 3α5α20α-HHP | 1.02 ± 0.28 | 0.71 ± 0.20 | 0.27 ± 0.02 | 0.50 ± 0.03 | Sex: F(1,16) = 0.04; p = 0.839 |
| Genotype: F(1,16) = 6.31; p = 0.023 | |||||
| Interaction: F(1,16) = 2.10; p = 0.167 | |||||
| 3β5α20α-HHP | 0.12 ± 0.04 | 0.03 ± 0.003** | 0.02 ± 0.002$ | 0.02 ± 0.005 | Sex: F(1,15) = 4.28; p = 0.056 |
| Genotype: F(1,15) = 6.74; p = 0.020 | |||||
| Interaction: F(1,15) = 4.61; p = 0.049 | |||||
| 3α5β20α-HHP | ND | 0.14 ± 0.06* | ND | 0.32 ± 0.02***$$ | |
| 20α -DHPREG | 0.02 ± 0.003 | 0.02 ± 0.005 | 0.02 ± 0.004 | 0.02 ± 0.003 | Sex: F(1,16) = 0.39; p = 0.540 |
| Genotype: F(1,16) = 2.18; p = 0.160 | |||||
| Interaction: F(1,16) = 0.14; p = 0.715 | |||||
| DOC | 0.15 ± 0.03 | 0.63 ± 0.22 | 0.14 ± 0.05 | 0.83 ± 0.25* | Sex: F(1,18) = 12.1; p = 0.003 |
| Genotype: F(1,17) = 0.30; p = 0.593 | |||||
| Interaction: F(1,17) = 0.43; p = 0.518 | |||||
| 5α-DHDOC | 5.09 ± 1.31 | 1.26 ± 0.12* | 3.34 ± 0.83 | 1.13 ± 0.22 | Sex: F(1,16) = 12.7; p = 0.003 |
| Genotype: F(1,16) = 1.24; p = 0.282 | |||||
| Interaction: F(1,16) = 0.92; p = 0.352 | |||||
| 5β-DHDOC | ND | 1.87 ± 0.33 | ND | 3.99 ± 1.60 | |
| 3α5α-THDOC | 0.97 ± 0.24 | 0.02 ± 0.01** | 0.57 ± 0.14 | 0.06 ± 0.02 | Sex: F(1,15) = 20.4; p = 0.0004 |
| Genotype: F(1,15) = 1.24; p = 0.283 | |||||
| Interaction: F(1,15) = 1.87; p = 0.192 | |||||
| 3α5β-THDOC | <0.005 | 0.14 ± 0.04 | <0.005 | 0.58 ± 0.27 | |
| Corticosterone | 82.1 ± 4.78 | 65.1 ± 11.2 | 46.8 ± 6.20$ | 76.2 ± 6.72 | Sex: F(1,17) = 0.55; p = 0.469 |
| Genotype: F(1,17) = 2.09; p = 0.167 | |||||
| Interaction: F(1,17) = 7.64; p = 0.013 | |||||
| Testosterone | 0.10 ± 0.02 | 0.002 ± 0.00003*** | 0.11 ± 0.03 | 0.01 ± 0.005*** | Sex: F(1,17) = 46.1; p < 0.0001 |
| Genotype: F(1,17) = 0.42; p = 0.523 | |||||
| Interaction: F(1,17) = 0.01; p = 0.913 | |||||
| 5α-DHT | 0.01 ± 0.001 | 0.002 ± 0.00002*** | 0.01 ± 0.003 | 0.002 ± 0.00003** | Sex: F(1,17) = 36.5; p < 0.0001 |
| Genotype: F(1,17) = 0.84; p = 0.373 | |||||
| Interaction: F(1,17) = 0.78; p = 0.389 | |||||
| 3α5α-THT | 0.02 ± 0.002 | 0.003 ± 0.001* | 0.03 ± 0.004 | 0.01 ± 0.01* | Sex: F(1,17) = 17.7; p = 0.0006 |
| Genotype: F(1,17) = 2.72; p = 0.117 | |||||
| Interaction: F(1,17) = 0.002; p = 0.969 | |||||
| Estradiol | 0.01 ± 0.001 | 0.01 ± 0.001 | 0.01 ± 0.001 | 0.01 ± 0.003 | Sex: F(1,17) = 0.11; p = 0.741 |
| Genotype: F(1,17) = 0.31; p = 0.587 | |||||
| Interaction: F(1,17) = 2.30; p = 0.148 | |||||
Results are presented as means ± SEM (n = 5–6 per group) and were analyzed by two-way ANOVA (sex × genotype) for brain and plasma followed by Newman–Keuls multiple-comparisons tests.
***p < 0.001,
**p < 0.01,
*p < 0.05 versus males; $$$p < 0.001, $$p < 0.01, $$p < 0.05 versus PRloxP/loxP.
Abbreviations of steroids are as for Table 1. ND, Not detected.
Levels of the major steroids measured in the PRloxP/loxP and PRNesCre mice were comparable to those reported in Figure 2 and Table 3 for wild-type C57BL/6 mice. Levels of pregnenolone were much higher in brain than in plasma, whereas similar low levels of progesterone were measured. Important for our purpose was the confirmation of much higher levels of PR active 5α-DHP in the male brain compared with females and plasma (Table 5). Furthermore, levels of 20α-DHP in brain and plasma were ∼10–20 times higher in females compared with males. However, absolute levels of 20α-DHP in this experiment were much higher than those shown in Figure 2. This difference may not necessarily reflect strain variations, but it may have been caused by higher levels of stress and adrenal activity. Indeed, levels of the stress response hormone corticosterone were also elevated in the present experiment. Results further confirm higher levels of 5β-reduced metabolites of progesterone and DOC in females compared with males.
Importantly, steroid levels were comparable between PRloxP/loxP and PRNesCre mice for PR-active progesterone and 5α-DHP (Table 5). However, plasma and brain levels of corticosterone were lower in PRNesCre males compared with PRloxP/loxP males. This may also have contributed to the slightly reduced levels of some 5α-reduced metabolites of progesterone and DOC in PRNesCre mice.
Changes in brain levels of progesterone and its metabolites after MCAO in male and female PRloxP/loxP and PRNesCre mice
PRNesCre mice were generated to uncover the cerebroprotective effects of endogenous brain progesterone and 5α-DHP, both active PR ligands, in males. It was thus important to examine whether levels of progesterone and its active metabolites were increased rapidly after MCAO in the transgenic male PRloxP/loxP and PRNesCre mice, as they are in wild-type males (Fig. 3). Brain levels of progesterone and its major reduced metabolites were analyzed by GC-MS/MS in the left infarcted hemisphere of PRloxP/loxP and PRNesCre male and female mice at 6 h after sham surgery or MCAO.
Analysis by GC-MS/MS revealed similar steroid profiles for PRloxP/loxP and PRNesCre mice at 6 h after sham surgery or MCAO (Fig. 6). However, brain levels of both progesterone and 20α-DHP were significantly higher in sham-operated PRNesCre males compared with sham-operated PRloxP/loxP males, suggesting a greater sensitivity of the former to surgical stress (Fig. 6B,E). After MCAO, brain levels of progesterone were elevated in the male brain and they were three times higher in males than in females (Fig. 6B). Brain levels of 5α-DHP were nearly 10 times higher in males compared with females and were upregulated markedly at 6 h after MCAO in males compared with sham operation (Fig. 6C). No increases in the levels of progesterone and 5α-DHP were found after MCAO in females, consistent with results presented in Figure 3 for C57BL/6 mice. Levels of 3α,5α-THP were also higher in males than in females, but were not upregulated in response to MCAO (Fig. 6D). Levels of 20α-DHP and 5α,20α-THP were low in both sham- and MCAO-operated males and females (Fig. 6E,F).
Figure 6.
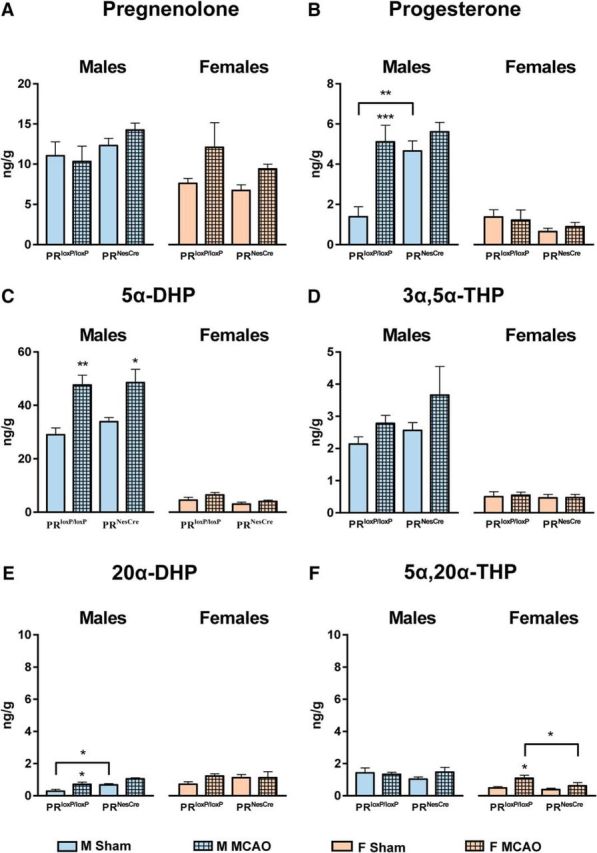
Levels of pregnenolone and progesterone and its major metabolites were analyzed by GC-MS/MS in the left hemisphere of males and females PRloxP/loxP and PRNesCre mice at 6 h after sham operation or MCAO. A, Levels of pregnenolone did not differ significantly between sexes and genotypes and were not affected by MCAO. B, Levels of progesterone were upregulated significantly in response to MCAO in PRloxP/loxP males. In contrast to PRloxP/loxP males, brain levels of progesterone were elevated after sham surgery and MCAO in PRNesCre males (effect of MCAO: F(1,19) = 15.1, p = 0.001; effect of genotype: F(1,19) = 9.85, p = 0.005; interaction between MCAO and genotype: F(1,19) = 5.29, p = 0.033). In females, levels of progesterone were lower compared with males and similar in sham and MCAO mice of both genotypes. C, Levels of 5α-DHP were much higher in males than in females and were only increased in response to MCAO in males (effect of MCAO in males: F(1,21) = 23.1, p < 0.0001). There were no differences for 5α-DHP levels between PRloxP/loxP and PRNesCre mice. D, As for 5α-DHP, brain levels of 3α,5α-THP were much higher in males than in females. Neither in males nor in females was there an effect of MCAO or genotype. E, Similar low levels of 20α-DHP (< 2 ng/g) were measured in the brain of PRloxP/loxP and PRNesCre males and females after sham surgery or MCAO. However, despite the low levels of 20α-DHP, they were upregulated significantly after MCAO in PRloxP/loxP males and were higher in sham-operated PRNesCre males compared with sham-operated PRloxP/loxP males (effect of MCAO in males: F(1,21) = 13.9, p = 0.001; effect of genotype in males: F(1,21) = 11.8, p = 0.003). F, As in the normal brain, levels of 5α,20α-THP were low after sham surgery or MCAO in both sexes. In females, 5α,20α-THP levels were slightly upregulated after MCAO in PRloxP/loxP mice (effect of MCAO in females: F(1,19) = 8.29, p = 0.010; effect of genotype in females: F(1,19) = 3.94, p = 0.062). Results are presented as means ± SEM (n = 5–7 per group) and were analyzed by 2-way ANOVA (MCAO × genotype) for males and females separately followed by Newman–Keuls multiple-comparisons tests. ***p < 0.001, **p < 0.01, *p < 0.05 compared with sham or as indicated.
Therefore, the response of brain steroids to MCAO surgery differs between sexes. Whereas males upregulate the production of PR active progesterone and 5α-DHP in response to ischemic injury, females do not show such changes.
Resistance of the brain to ischemic damage is reduced in young male and female PRNesCre mice
For all of the outcomes measured after MCAO, we first verified that they were similar for C57BL/6 and PRloxP/loxP mice. Neurological scores, time spent on the rotarod, and infarct volumes did not differ significantly between the two strains of mice and we used PRloxP/loxP mice as controls for the effect of targeted PR inactivation in the CNS.
If the endogenous PR ligands progesterone and 5α-DHP play a role in rapid endogenous cerebroprotective mechanisms, then deletion of PRs in the brain could be expected to reduce its resistance to ischemic injury. We therefore compared the total infarct volume, neurological outcomes, neuron densities, and microglial activation between young male and female PRloxP/loxP and PRNesCre mice at 6 h after MCAO (Figs. 7, 8, 9, left). Although sham surgery seemed to have an influence on the brain levels of some progesterone metabolites, it did not cause ischemic damage. Indeed, sham surgery did not affect cerebral blood flow, neurological deficit scores, or infarct volume.
Figure 7.
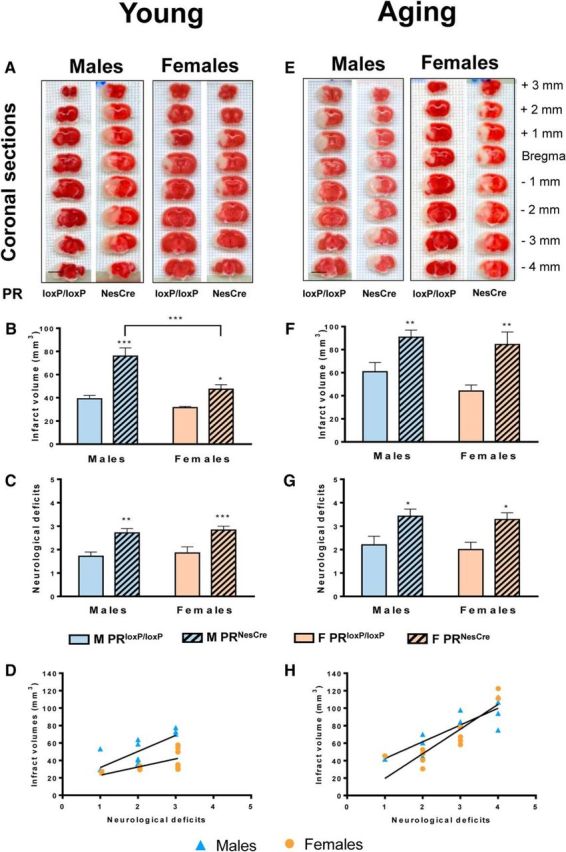
Infarct volumes and neurological deficits at 6 h after MCAO were increased in young and aging mice of both sexes after deletion of PR in the brain. Total infarct volumes and neurological deficit scores (higher scores reflect higher disability) were analyzed in young (A–D) and aging (E–H) PRloxP/loxP and PRNesCre mice. A, E, Representative 1-mm-thick coronal brain sections of young and aging male and female PRloxP/loxP and PRNesCre mice stained with 2,3,5-triphenyltetrazolium chloride. Infarcted tissue appears pale and normal tissue is colored dark red. Scale bar, 5 mm. B, In young males and females, total infarct volumes were significantly larger in PRNesCre mice compared with PRloxP/loxP mice. However, for PRNesCre mice, the infarct volume was significantly larger in males than in females. Data analysis by two-way ANOVA revealed significant effects of genotype (F(1,23) = 36.7, p < 0.0001) and sex (F(1,23) = 17.4, p = 0.0004), as well as a significant interactions between both factors (F(1,23) = 5.89, p = 0.024). C, Neurological deficits were analyzed by Kruskal–Wallis ANOVA (p = 0.003). In both sexes, neurological deficits were greater in PRNesCre mice compared with PRloxP/loxP mice. D, Spearman's test showed highly positive correlations between neurological deficit scores and infarct volumes for males (r = 0.86, p = 0.0003) and females (r = 0.70, p = 0.009). F, In aging males and females, total infarct volumes were significantly larger in PRNesCre mice compared with PRloxP/loxP mice. In contrast to young mice, infarct volumes were comparable between males and females. Indeed, two-way ANOVA only showed a significant effect of genotype (F(1,20) = 18.7, p = 0.0003). However, infarct volumes were larger in aging mice compared with young mice, as revealed by a two-way ANOVA with age and sex as factors (effect of age: PRloxP/loxP mice: F(1,20) = 15.2, p = 0.0009; PRNesCre mice: F(1,23) = 12.6, p = 0.002). G, As for young mice, neurological deficits were greater in PRNesCre mice compared with PRloxP/loxP mice (p = 0.017 by Kruskal–Wallis analysis). Interestingly, neurological deficits were higher in aging than in young PRNesCre mice (p = 0.018 by Mann–Whitney test), but there was no such effect of age on functional outcomes in PRloxP/loxP mice. H, Spearman's test showed strong positive correlations between neurological deficit scores and infarct volumes for both aging males (r = 0.81, p = 0.002) and females (r = 0.92, p < 0.001). Results are presented as means ± SEM (n = 5–7 per group). Two-way ANOVA (sex × genotype) followed by Newman–Keuls multiple-comparisons tests were used for infarct volume analysis. ***p < 0.001, **p < 0.01, *p < 0.05 compared with PRloxP/loxP mice or as indicated.
Figure 8.
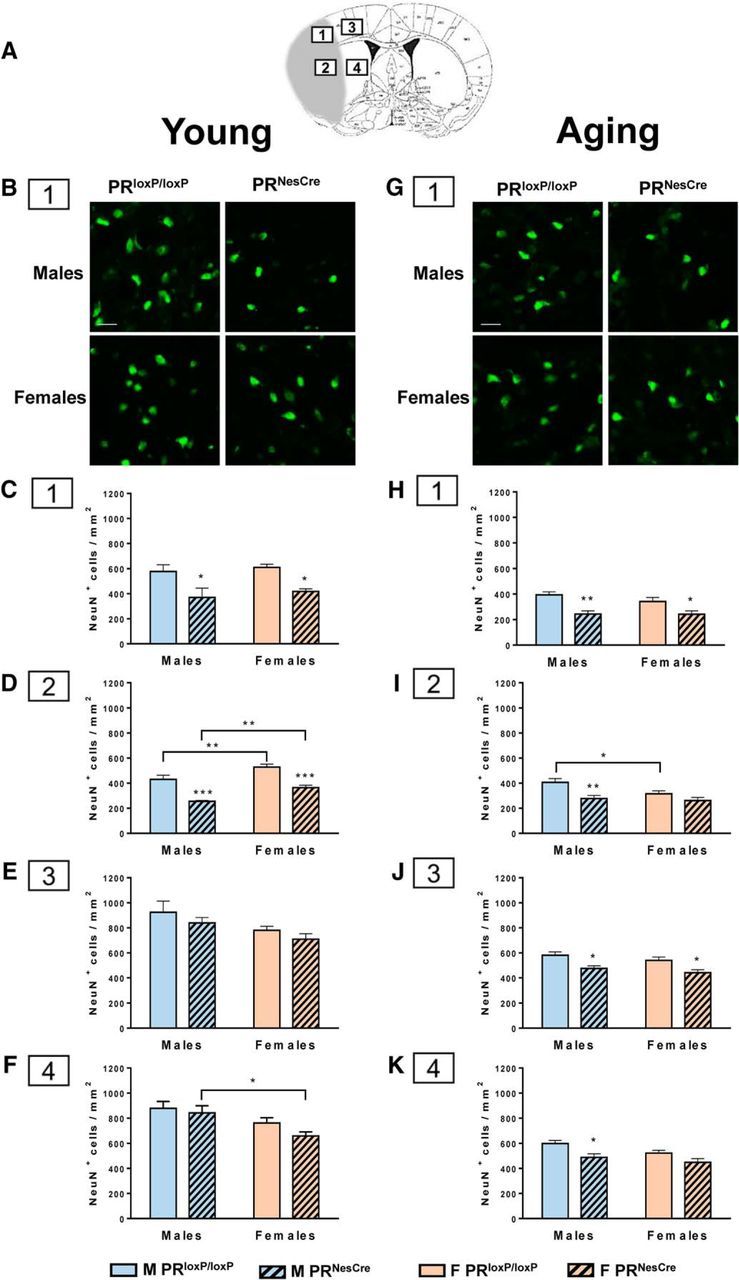
The density of NeuN+ neurons at 6 h after MCAO was decreased in the ipsilateral ischemic hemisphere in young and aging mice after deletion of PR in the brain. A, Localization of the four regions where neurons were counted at the level of the cerebral cortex and striatum. The regions were located on the infarct and peri-infarct, which were delimited by DAPI staining of cell nuclei. The regions are drawn on a coronal brain section taken from the atlas of Franklin and Paxinos (2008) at the level of bregma +0.14. The infarct area is schematically presented in gray. B, G, Representative immunofluorescence staining of NeuN+ neurons in region 1 (infarct in the cerebral cortex) of young and aging PRloxP/loxP and PRNesCre mice of both sexes. Scale bar, 100 μm. B–F, Neuronal density in young mice. C, In region 1 (infarct in the cerebral cortex), there were no differences between males and females, but the density of NeuN+ neurons was significantly lower in PRNesCre mice compared with PRloxP/loxP mice (two-way ANOVA with sex and genotype as factors; effect of genotype: F(1,20) = 20.1, p = 0.0002). D, In region 2 (infarct in the striatum), the density of NeuN+ neurons was also significantly lower in PRNesCre mice compared with PRloxP/loxP mice (effect of genotype: F(1,20) = 48.8, p < 0.0001) for both sexes. However, neuron density was slightly but significantly higher in females than in males for both PRloxP/loxP and PRNesCre mice (effect of sex: F(1,20) = 18.3, p = 0.0004). E, F, in the peri-infarct in the cerebral cortex (E) and striatum (F), neuronal density was about twice as high as at the infarct. There were no differences between PRloxP/loxP and PRNesCre mice, but the density of neurons tended to be higher in males (effect of sex in region 3: F(1,20) = 5.84, p = 0.025 and in region 4: F(1,20) = 10.2, p = 0.005). G–K, Neuronal density in aging mice. H, As in region 1 of young mice, the density of NeuN+ neurons was significantly lower in PRNesCre males and females compared with PRloxP/loxP mice (effect of genotype: F(1,20) = 20.7, p = 0.0002). However, the density of neurons was lower in aging mice compared with young mice as revealed by a two-way ANOVA with age and sex as factors (effect of age: PRloxP/loxP mice: F(1,19) = 37.8, p < 0.0001; PRNesCre mice: F(1,21) = 16.7, p = 0.0005). I, In region 2, the density of NeuN+ neurons was significantly lower in PRNesCre males compared with PRloxP/loxP males (effect of genotype: F(1,20) = 11.7, p = 0.003). Again, a two-way ANOVA with age and sex as factors showed that the density of neurons was lower in aging mice compared with young mice (effect of age: PRloxP/loxP mice: F(1,19) = 17.2, p = 0.0006; PRNesCre mice: F(1,21) = 3.20, p = 0.088). J, K, In contrast to the young mice, neuronal density was lower in the peri-infarct in cerebral cortex (J) and striatum (K) of PRNesCre mice compared with PRloxP/loxP (effect of genotype in region 3: F(1,20) = 16.7, p = 0.0006 and in region 4: F(1,20) = 10.6, p = 0.004). Again, the density of neurons was lower in the aging mice compared with the young mice (p < 0.001 for regions 3 and 4 of PRNesCre and PRloxP/loxP mice). Results are presented as means ± SEM (n = 5–7 per group) and were analyzed by Newman–Keuls multiple-comparisons tests after two-way ANOVA (sex × genotype). ***p < 0.001, **p < 0.01, *p < 0.05 compared with PRloxP/loxP mice or as indicated.
Figure 9.
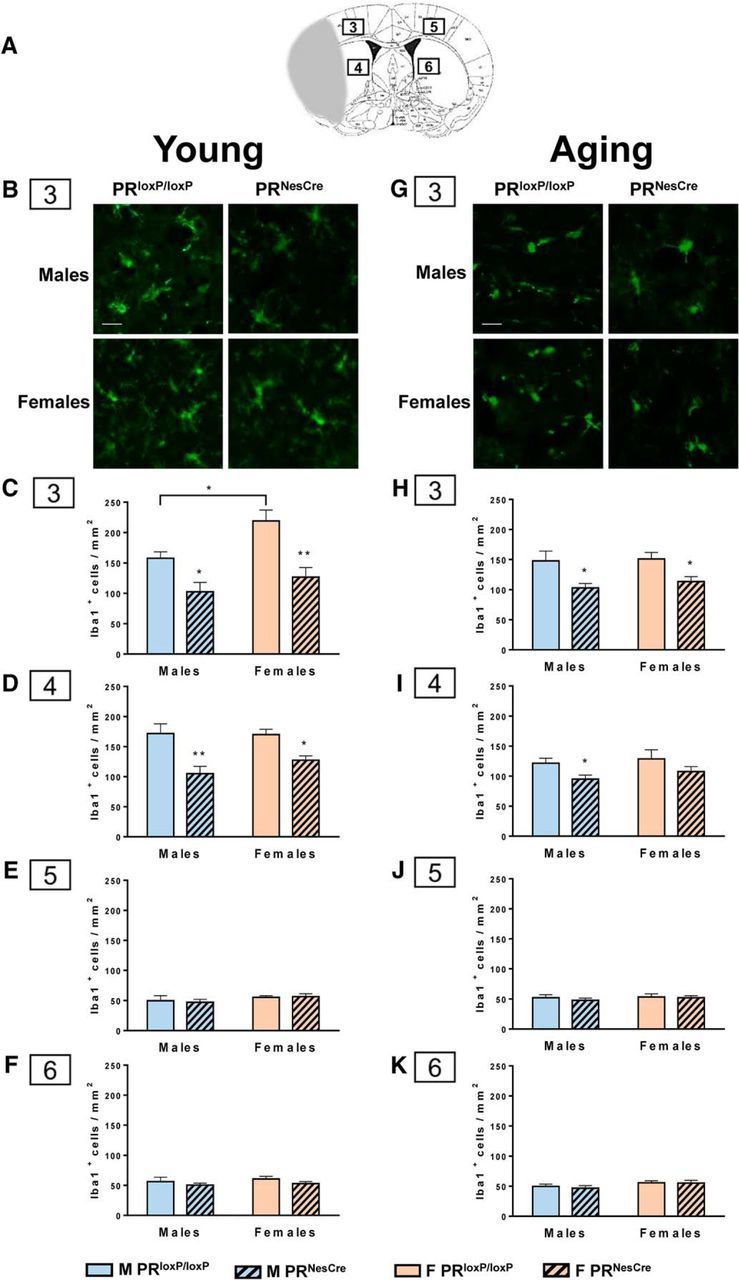
The density of activated Iba1+ microglial cells at 6 h after MCAO was decreased in the peri-infarct in young and aging mice after deletion of PR in the brain. A, Localization of the four regions where microglial cells were counted at the level of the cerebral cortex and striatum. The regions are drawn on a coronal brain section taken from the atlas of Franklin and Paxinos (2008) at the level of bregma +0.14. The infarct area is presented schematically in gray. B, G, Representative immunofluorescence staining of Iba1+ microglial cells in region 3 (peri-infarct in the cerebral cortex) of young and aging PRloxP/loxP and PRNesCre mice of both sexes. Scale bar, 100 μm. B–F, Microglial density in young mice. C, In region 3 (peri-infarct in the cerebral cortex), the density of Iba1+ cells was lower in PRNesCre mice compared with PRloxP/loxP mice (effect of genotype: F(1,18) = 22.8, p = 0.0002). In PRloxP/loxP mice, the density of microglial cells was slightly higher in females (effect of sex: F(1,18) = 7.68, p = 0.013). D, In region 4 (peri-infarct in the striatum), as in cerebral cortex, the density of Iba1+ cells was lower in PRNesCre mice compared with PRloxP/loxP mice (effect of genotype: F(1,18) = 22.8, p = 0.0002). E, F, In contrast, the density of microglial cells was low and not affected by the inactivation of PR or the sex of the mice in cerebral cortex (E) and striatum (F) of the contralateral nonischemic hemisphere (in regions 5 and 6). G–K, Microglial density in aging mice. H, I, in the peri-infarct in cerebral cortex (H) and striatum (I), the density of Iba1+ microglial cells was lower in PRNesCre mice compared with PRloxP/loxP mice (effect of genotype: cerebral cortex: F(1,20) = 15.6, p = 0.0008; striatum: F(1,20) = 6.02, p = 0.024). J, K, As for the young mice, the density of microglial cells was low and not affected by the inactivation of PR in cerebral cortex (J) and striatum (K) of the contralateral nonischemic hemisphere (in regions 5 and 6) of both sexes. Results are presented as means ± SEM (n = 5–7 per group) and were analyzed by Newman–Keuls multiple-comparisons tests after two-way ANOVA (sex × genotype). ***p < 0.001, **p < 0.01, *p < 0.05 compared with PRloxP/loxP mice or as indicated.
Infarct volumes were increased significantly in PRNesCre mice of both sexes compared with PRloxP/loxP mice (Fig. 7A,B). In males, the size of the infarct was nearly doubled after deletion of PRs, whereas in PRNesCre females, the infarct was increased by 30%. Infarct volumes did not differ between sexes in PRloxP/loxP mice, but were significantly larger in male than in female PRNesCre mice (Fig. 7B), pointing to the existence of additional PR-independent protective mechanisms in females.
Neurological deficits were assessed on a six-point scale, with elevated scores reflecting higher functional disability (Gibson et al., 2011). In contrast to the infarct size, they were similarly increased in male and female PRNesCre males compared with PRloxP/loxP mice (Fig. 7C). Therefore, endogenous PR-dependent signaling contributed to the preservation of neural functions in both sexes. Highly positive correlations were observed between infarct volumes and neurological deficits in both sexes (Fig. 7D).
To go further in the evaluation of the tissue damage at 6 h after MCAO, the density of neurons labeled with an antibody against the neuronal nuclear antigen NeuN was measured by immunofluorescence analysis. Neurons were counted in the infarct and in the peri-infarct at the levels of the cerebral cortex and striatum (Fig. 8A,B). In both brain regions, MCAO is known to produce widespread ischemic cell death, with the most rapid damage occurring in the striatum (Carmichael, 2010). DAPI counterstaining of cell nuclei allowed us to define the demarcation between the infarct and peri-infarct areas.
At the infarct core in both cerebral cortex and striatum, neuron density was significantly lower in PRNesCre mice compared with PRloxP/loxP mice for both sexes (Fig. 8C,D). At the peri-infarct area, the density of neurons was almost twice that counted in the infarct core and no differences were observed between PRNesCre mice and PRloxP/loxP mice (Fig. 8E,F). Similar neuronal densities with no differences between PRNesCre mice and PRloxP/loxP mice were also observed in the contralateral hemisphere. Sex differences in the density of NeuN+ neurons were observed after MCAO in the striatum (Fig. 8D,F), but not in the cortex (Fig. 8C,E).
The density of activated microglia was assessed by immunofluorescence analysis of Iba1+ cells in cerebral cortex and striatum. As early as 6 h after MCAO, microglial activation was only observed in the peri-infarct (Fig. 9B–D). In both cortex and striatum, the density of activated microglial cells was significantly lower in PRNesCre mice compared with PRloxP/loxP mice for both sexes (Fig. 9C,D). The density of Iba1+ microglial cells was low in the contralateral nonischemic side and was unaffected by the PR deletion (Fig. 9E,F).
Resistance of the brain to ischemic damage is also reduced in aging male and female mice after deletion of PRs
To determine whether the endogenous PR-dependent cerebroprotective mechanisms remain functional during aging, infarct volume, neurological outcomes, neuron densities, and microglial activation were examined at 6 h after MCAO in aging (12–14 months old) male and female PRloxP/loxP and PRNesCre mice (Figs. 7, 8, 9, right) and were compared with the results obtained for young (3 months old) mice (Figs. 7, 8, 9, left).
In aging mice, the invalidation of PRs also resulted in larger infarct volumes and there were no differences between sexes (Fig. 7E,F). As expected, infarct volumes were larger in aging compared with young mice (Fig. 7B,F). Neurological deficit scores were increased to the same extent in both sexes, reflecting higher functional disability (Fig. 7G). Interestingly, neurological deficits tended to be higher in aging than in young mice, but only for the PRNesCre mice. As for the young mice, positive correlations were observed between infarct volumes and neurological deficits (Fig. 7H). Therefore, endogenous PR signaling continues to play an important role in cerebroprotection in aging males and females.
Densities of neurons were lower in the aging males and females compared with young adults in infarct and peri-infarct areas of cortex and striatum (Fig. 8). Sex differences in the density of NeuN+ neurons were only observed in the infarct at the level of the striatum in aging PRloxP/loxP mice (Fig. 8I). Neuron density was significantly lower in PRNesCre mice compared with PRloxP/loxP mice, not only in the infarct region, but also in the peri-infarct area (Fig. 8H–K), suggesting a more rapid spreading of the ischemic lesion in aging compared with young mice. The reduced number of neurons after inactivation of PR in the aging brain uncovered a role for endogenous PR signaling in neuroprotection in cerebral cortex and dorsal striatum in both infarct and peri-infarct.
Activated Iba1+ microglial cells were observed in the peri-infarct of aging mice at 6 h after MCAO (Fig. 9G–I). The density of Iba1+ cells was lower in aging mice compared with young mice, at the level of the striatum. In both cerebral cortex and striatum, the density of activated microglial cells in the peri-infarct was lower in PRNesCre mice compared with PRloxP/loxP mice, thus confirming a role of endogenous PR signaling in the activation of microglial responses (Fig. 9H,I). In the contralateral nonischemic side of the cerebral cortex and striatum, the density of Iba1+ cells was low with no effect of PR deletion (Fig. 9J,K).
Discussion
In this study, we demonstrate the importance of endogenous progesterone, its metabolites, and neural PRs, in cerebroprotection in young and aging mice of both sexes during the early acute phase after stroke. We chose females in diestrus, a stage lasting >2 d of the 4 d reproductive cycle (Byers et al., 2012) because it was considered to be advisable to avoid short peak levels of ovarian progesterone and estradiol (Smith et al., 1975; Nilsson et al., 2015) in order to uncover a role of endogenous brain progesterone in female cerebroprotection. Only limited information is available concerning the influence of the estrus cycle on responses to cerebral ischemia, but one study reported that hypertensive female rats develop smaller cerebral infarcts during the short proestrus stage (Carswell et al., 2000).
The unexpected results of brain steroid profiling by GC-MS/MS lent further support to the notion that progesterone and its metabolites are important brain steroids in males. Whereas levels of neuroactive 5α-reduced metabolites of progesterone were particularly elevated in the male brain, females instead produced larger amounts of 20α-DHP. This metabolite of progesterone has been reported to have little affinity for PR and no progestogenic activity (Nadeem et al., 2016). Instead, because the formation of the 20α-hydroxy-pregnanes is reversible, they may regulate the availability of biologically active 20-keto-pregnanes (Pelletier, 2010). Even more striking was the rapid and marked increase in brain levels of the PR ligands progesterone and 5α-DHP after MCAO in males but not in females.
Because of its elevated endogenous brain levels of progesterone and 5α-DHP, one may have predicted the male brain to be more resistant to ischemic damage than the female brain. However, this was not the case under the present experimental conditions: infarct volume and neurological deficit scores did not differ between male and female PRloxP/loxP mice at 6 h after MCAO. Moreover, in both sexes, infarct volumes and neurological deficits were increased and neuronal densities decreased in PRNesCre mice, demonstrating an important role of PR signaling in the resistance of brain tissue to ischemic damage for both sexes. Somehow, the low brain levels of progesterone and 5α-DHP in females were sufficient for efficient cerebroprotection. However, males may rely more than females on cerebroprotection by endogenous progesterone and 5α-DHP, as suggested by the larger infarct volumes in PRNesCre males compared with PRNesCre females. These observations point to additional protective mechanisms against ischemic injury in the female brain.
An auxiliary cerebroprotective agent may be estradiol despite the fact that similar low brain levels of the hormone were measured in both sexes and they were not increased in response to MCAO. Females may indeed be more sensitive to the cerebroprotective effects of estradiol than males because estrogen receptor α (ERα) is upregulated rapidly in the ischemic brain of females but not males (Dubal et al., 2006; Westberry et al., 2008). Estrogens may potentiate the cerebroprotective effects of progesterone in females by upregulating PR expression (Waters et al., 2008).
Steroid profiling revealed other potential cerebroprotective mechanisms in females. Therefore, whereas progesterone is mainly converted to 5α-reduced metabolites in males, females produce higher amounts of GABAA receptor active 5β-reduced metabolites of progesterone and DOC, respectively, 3α,5β-THP, and 3α,5β-THDOC (Gunn et al., 2015). The positive modulation of GABAA receptors by 3α-reduced metabolites of progesterone and DOC and the resulting decrease in neuronal excitability is indeed a well recognized mechanism involved in neuroprotective responses (Sayeed et al., 2006; Brinton, 2013; Melcangi and Panzica, 2014).
Another sex difference deserving attention was the exposure of the male brain to higher amounts of injury-induced corticosterone compared with females. Endogenous corticosterone and stress have indeed been shown to exacerbate brain injury resulting from cerebral ischemia and the suppression of glucocorticoid production or actions has been proposed as a therapeutic option for stroke patients (Smith-Swintosky et al., 1996). Progesterone has been shown recently to counter the deleterious effects of stress after brain ischemia (Espinosa-Garcia et al., 2017). In males, higher levels of progesterone and 5α-DHP may protect the brain against the ischemia-induced rise in corticosterone, whereas lower levels of glucocorticoids may improve the resistance of the female brain to ischemic damage.
Previous studies have shown that CNS levels of progesterone are increased in response to traumatic brain and spinal cord injury and MCAO (Labombarda et al., 2006; Meffre et al., 2007; Liu et al., 2012; Lopez-Rodriguez et al., 2015, 2016). Because treatment with progesterone exerts protective effects in these different injury models, it has been presumed that increased production of progesterone may be part of protective responses of neural cells to injury (De Nicola et al., 2009). Here, we provide evidence for this concept by showing that deletion of PRs in the brain results in increased ischemic damage and aggravation of neurological deficits after MCAO in the absence of treatment with exogenous progesterone.
We have shown previously that the resistance of the brain to ischemic damage is decreased in total PR knock-out mice (Liu et al., 2012). However, this study did not allow us to distinguish between central and peripheral actions of progesterone. The present results demonstrate a key role of neural PR in the cerebroprotective effects of the hormone. This finding has two major implications. First, synthetic progestins, which have been designed to target PRs selectively for contraceptive purposes or for the treatment of endocrine disorders, may be used as cerebroprotective agents (Sitruk-Ware and El-Etr, 2013; Schumacher et al., 2014). However, synthetic progestins cannot mimic the complex and regulated metabolism of natural progesterone in the brain (Kumar et al., 2017). Second, a route of progesterone administration with direct access to the brain such as intranasal delivery may be particularly advantageous for acute stroke therapy (Fréchou et al., 2015).
In men and women, age is the most important risk factor for stroke and a prognostic marker for poor outcomes (Wong et al., 2013). Concordantly, studies involving aging animals report worse stroke outcomes (Liu et al., 2009). The present study confirms that infarct volumes are larger in aging than in young mice. Moreover, neurological deficits were more severe in aging PRNesCre mice. Treatment with progesterone continues to reduce infarct volumes in aging rodents of both sexes (Wang et al., 2010; Gibson et al., 2011; Wali et al., 2016). Here, we show that endogenous PR-dependent cerebroprotective mechanisms remain operational in aging males and females: infarct volumes and neurological deficits were increased and the density of neurons decreased in PRNesCre mice compared with PRloxP/loxP mice. The preservation of PR-dependent cerebroprotective mechanisms at the early acute phase after stroke could explain why the 6 h time window for treatment by progesterone is preserved in aging males and females (Yousuf et al., 2014).
In the PRNesCre mice, PR expression was ablated in neurons, astrocytes, and oligodendrocytes, but not in microglial cells. The latter do not express PR even after lipopolysaccharide challenge (Sierra et al., 2008). Cell-specific inactivation of PR will allow us identify the neural cells on which progesterone and 5α-DHP act.
Using GC-MS/MS technology for steroid profiling and conditional CNS-specific PR knock-out mice, the present study demonstrates an important role of endogenous progesterone and its metabolites in the resistance of brain tissue to ischemic damage. Importantly, endogenous PR-dependent cerebroprotective mechanisms are operational in young and aging males and females. Remarkably, summed brain levels of PR-active pregnanes are much higher in males than in females and they are specifically upregulated in males after MCAO. Therefore, progesterone and its neuroactive metabolites may play a particularly important role in the male brain.
Footnotes
X.Z. was funded by a Scholarship from the China Scholarship Council. We thank Pr. Pierre Chambon and the Mouse Clinical Institute (MCI, Illkirch, France) for the generous gift of the PR+/loxP mice and Brigitte Delespierre, Fanny Aprahamian, and Annie Cambourg for excellent technical assistance.
The authors declare no competing financial interests.
References
- Baulieu EE. (1997) Neurosteroids: of the nervous system, by the nervous system, for the nervous system. Recent Prog Horm Res 52:1–32. [PubMed] [Google Scholar]
- Belelli D, Lambert JJ, Peters JA, Gee KW, Lan NC (1996) Modulation of human recombinant GABAA receptors by pregnanediols. Neuropharmacology 35:1223–1231. 10.1016/S0028-3908(96)00066-4 [DOI] [PubMed] [Google Scholar]
- Brinton RD. (2013) Neurosteroids as regenerative agents in the brain: therapeutic implications. Nat Rev Endocrinol 9:241–250. 10.1038/nrendo.2013.31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown TJ, Clark AS, MacLusky NJ (1987) Regional sex differences in progestin receptor induction in the rat hypothalamus: effects of various doses of estradiol benzoate. J Neurosci 7:2529–2536. [PMC free article] [PubMed] [Google Scholar]
- Byers SL, Wiles MV, Dunn SL, Taft RA (2012) Mouse estrous cycle identification tool and images. PLoS One 7:e35538. 10.1371/journal.pone.0035538 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell BCV, Donnan GA, Lees KR, Hacke W, Khatri P, Hill MD, Goyal M, Mitchell PJ, Saver JL, Diener HC, Davis SM (2015) Endovascular stent thrombectomy: the new standard of care for large vessel ischaemic stroke. Lancet Neurol 14:846–854. 10.1016/S1474-4422(15)00140-4 [DOI] [PubMed] [Google Scholar]
- Carmichael ST. (2010) Targets for neural repair therapies after stroke. Stroke 41:S124–S126. 10.1161/STROKEAHA.110.597146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carswell HV, Dominiczak F, Macrae IM (2000) Estrogen status affects sensitivity to focal cerebral ischemia in stroke-prone spontaneously hypertensive rats. Am J Physiol Hear Circ Physiol 278:H290–H294. [DOI] [PubMed] [Google Scholar]
- Celotti F, Melcangi RC, Martini L (1992) The 5 alpha-reductase in the brain: molecular aspects and relation to brain function. Front Neuroendocrinol 13:163–215. [PubMed] [Google Scholar]
- Chen M, Penning TM (2014) 5beta-Reduced steroids and human Delta(4)-3-ketosteroid 5beta-reductase (AKR1D1). Steroids 83:17–26. 10.1016/j.steroids.2014.01.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Nicola AF, Labombarda F, Gonzalez Deniselle MC, Gonzalez SL, Garay L, Meyer M, Gargiulo G, Guennoun R, Schumacher M (2009) Progesterone neuroprotection in traumatic CNS injury and motoneuron degeneration. Front Neuroendocrinol 30:173–187. 10.1016/j.yfrne.2009.03.001 [DOI] [PubMed] [Google Scholar]
- Donnan GA, Davis SM, Parsons MW, Ma H, Dewey HM, Howells DW (2011) How to make better use of thrombolytic therapy in acute ischemic stroke. Nat Rev Neurol 7:400–409. 10.1038/nrneurol.2011.89 [DOI] [PubMed] [Google Scholar]
- Dubal DB, Rau SW, Shughrue PJ, Zhu H, Yu J, Cashion AB, Suzuki S, Gerhold LM, Bottner MB, Dubal SB, Merchanthaler I, Kindy MS, Wise PM (2006) Differential modulation of estrogen receptors (ERs) in ischemic brain injury: A role for ERα in estradiol-mediated protection against delayed cell death. Endocrinology 147:3076–3084. 10.1210/en.2005-1177 [DOI] [PubMed] [Google Scholar]
- Espinosa-Garcia C, Sayeed I, Yousuf S, Atif F, Sergeeva EG, Neigh GN, Stein DG (2017) Stress primes microglial polarization after global ischemia: therapeutic potential of progesterone. Brain Behav Immun pii: S0889-1591(17)30201–5. 10.1016/j.bbi.2017.06.012 [DOI] [PubMed] [Google Scholar]
- Fisher M. (2011) New approaches to neuroprotective drug development. Stroke 42:S24–S27. 10.1161/STROKEAHA.110.592394 [DOI] [PubMed] [Google Scholar]
- Franklin K, Paxinos G (2008) The mouse brain in stereotaxic coordinates, 3rd ed Amsterdam: Elsevier. [Google Scholar]
- Fréchou M, Zhang S, Liere P, Delespierre B, Soyed N, Pianos A, Schumacher M, Mattern C, Guennoun R (2015) Intranasal delivery of progesterone after transient ischemic stroke decreases mortality and provides neuroprotection. Neuropharmacology 97:394–403. 10.1016/j.neuropharm.2015.06.002 [DOI] [PubMed] [Google Scholar]
- Gibson CL, Murphy SP (2004) Progesterone enhances functional recovery after middle cerebral artery occlusion in male mice. J Cereb Blood Flow Metab 24:805–813. 10.1097/01.WCB.0000125365.83980.00 [DOI] [PubMed] [Google Scholar]
- Gibson CL, Coomber B, Murphy SP (2011) Progesterone is neuroprotective following cerebral ischaemia in reproductively ageing female mice. Brain 134:2125–2133. 10.1093/brain/awr132 [DOI] [PubMed] [Google Scholar]
- Golanov EV, Reis DJ (1995) Contribution of cerebral edema to the neuronal salvage elicited by stimulation of cerebellar fastigial nucleus after occlusion of the middle cerebral artery in rat. J Cereb Blood Flow Metab 15:172–174. 10.1038/jcbfm.1995.19 [DOI] [PubMed] [Google Scholar]
- Grimm SL, Hartig SM, Edwards DP (2016) Progesterone receptor signaling mechanisms. J Mol Biol 428:3831–3849. 10.1016/j.jmb.2016.06.020 [DOI] [PubMed] [Google Scholar]
- Gunn BG, Cunningham L, Mitchell SG, Swinny JD, Lambert JJ, Belelli D (2015) GABA receptor-acting neurosteroids: A role in the development and regulation of the stress response. Front Neuroendocrinol 36:28–48. 10.1016/j.yfrne.2014.06.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill KK, Roemer SC, Churchill ME, Edwards DP (2012) Structural and functional analysis of domains of the progesterone receptor. Mol Cell Endocrinol 348:418–429. 10.1016/j.mce.2011.07.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillisch A, von Langen J, Menzenbach B, Droescher P, Kaufmann G, Schneider B, Elger W (2003) The significance of the 20-carbonyl group of progesterone in steroid receptor binding: A molecular dynamics and structure-based ligand design study. Steroids 68:869–878. 10.1016/j.steroids.2003.08.009 [DOI] [PubMed] [Google Scholar]
- Hosie AM, Wilkins ME, da Silva HM, Smart TG (2006) Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites. Nature 444:486–489. 10.1038/nature05324 [DOI] [PubMed] [Google Scholar]
- Iadecola C, Anrather J (2011) Stroke research at a crossroad: asking the brain for directions. Nat Neurosci 14:1363–1368. 10.1038/nn.2953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar N, Fagart J, Liere P, Mitchell SJ, Knibb AR, Petit-Topin I, Rame M, El-Etr M, Schumacher M, Lambert JJ, Rafestin-Oblin ME, Sitruk-Ware R (2017) Nestorone® (NES) a novel progestin for non-oral contraception: structure-activity relationships and brain metabolism studies. Endocrinology 158:170–182. 10.1210/en.2016-1426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Labombarda F, Pianos A, Liere P, Eychenne B, Gonzalez S, Cambourg A, De Nicola AF, Schumacher M, Guennoun R (2006) Injury elicited increase in spinal cord neurosteroid content analyzed by gas chromatography mass spectrometry. Endocrinology 147:1847–1859. 10.1210/en.2005-0955 [DOI] [PubMed] [Google Scholar]
- Lambert JJ, Cooper MA, Simmons RDJ, Weir CJ, Belelli D (2009) Neurosteroids: endogenous allosteric modulators of GABAA receptors. Psychoneuroendocrinology 34:48–58. [DOI] [PubMed] [Google Scholar]
- Lange CA, Gioeli D, Hammes SR, Marker PC (2007) Integration of rapid signaling events with steroid hormone receptor action in breast and prostate cancer. Annu Rev Physiol 69:171–199. 10.1146/annurev.physiol.69.031905.160319 [DOI] [PubMed] [Google Scholar]
- Liere P, Pianos A, Eychenne B, Cambourg A, Liu S, Griffiths W, Schumacher M, Sjövall J, Baulieu EE (2004) Novel lipoidal derivatives of pregnenolone and dehydroepiandrosterone and absence of their sulfated counterparts in rodent brain. J Lipid Res 45:2287–2302. 10.1194/jlr.M400244-JLR200 [DOI] [PubMed] [Google Scholar]
- Liu A, Margaill I, Zhang S, Labombarda F, Coqueran B, Delespierre B, Liere P, Marchand-Leroux C, O'Malley BW, Lydon JP, De Nicola AF, Sitruk-Ware R, Mattern C, Plotkine M, Schumacher M, Guennoun R (2012) Progesterone receptors: a key for neuroprotection in experimental stroke. Endocrinology 153:3747–3757. 10.1210/en.2012-1138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu F, Yuan R, Benashski SE, McCullough LD (2009) Changes in experimental stroke outcome across the life span. J Cereb Blood Flow Metab 29:792–802. 10.1038/jcbfm.2009.5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25:402–408. 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- Lopez-Rodriguez AB, Acaz-Fonseca E, Giatti S, Caruso D, Viveros MP, Melcangi RC, Garcia-Segura LM (2015) Correlation of brain levels of progesterone and dehydroepiandrosterone with neurological recovery after traumatic brain injury in female mice. Psychoneuroendocrinology 56:1–11. 10.1016/j.psyneuen.2015.02.018 [DOI] [PubMed] [Google Scholar]
- Lopez-Rodriguez AB, Acaz-Fonseca E, Spezzano R, Giatti S, Caruso D, Viveros MP, Melcangi RC, Garcia-Segura LM (2016) Profiling neuroactive steroid levels after traumatic brain injury in male mice. Endocrinology 157:3983–3993. 10.1210/en.2016-1316 [DOI] [PubMed] [Google Scholar]
- Lydon JP, DeMayo FJ, Funk CR, Mani SK, Hughes AR, Montgomery CA Jr, Shyamala G, Conneely OM, O'Malley BW (1995) Mice lacking progesterone receptor exhibit pleiotropic reproductive abnormalities. Genes Dev 9:2266–2278. 10.1101/gad.9.18.2266 [DOI] [PubMed] [Google Scholar]
- Meffre D, Pianos A, Liere P, Eychenne B, Cambourg A, Schumacher M, Stein DG, Guennoun R (2007) Steroid profiling in brain and plasma of male and pseudopregnant female rats after traumatic brain injury: Analysis by gas chromatography/mass spectrometry. Endocrinology 148:2505–2517. 10.1210/en.2006-1678 [DOI] [PubMed] [Google Scholar]
- Melcangi RC, Panzica GC (2014) Allopregnanolone: state of the art. Prog Neurobiol 113:1–5. 10.1016/j.pneurobio.2013.09.005 [DOI] [PubMed] [Google Scholar]
- Mergenthaler P, Dirnagl U (2011) Protective conditioning of the brain: expressway or roadblock? J Physiol 589:4147–4155. 10.1113/jphysiol.2011.209718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murr SM, Stabenfeldt GH, Bradford GE, Geschwind II (1974) Plasma testosterone during pregnancy in the mouse. Endocrinology 94:1209–1211. 10.1210/endo-94-4-1209 [DOI] [PubMed] [Google Scholar]
- Nadeem L, Shynlova O, Matysiak-Zablocki E, Mesiano S, Dong X, Lye S (2016) Molecular evidence of functional progesterone withdrawal in human myometrium. Nat Commun 7:11565. 10.1038/ncomms11565 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nilsson ME, Vandenput L, Tivesten Å, Norlén AK, Lagerquist MK, Windahl SH, Börjesson AE, Farman HH, Poutanen M, Benrick A, Maliqueo M, Stener-Victorin E, Ryberg H, Ohlsson C (2015) Measurement of a comprehensive sex steroid profile in rodent serum by high-sensitive gas chromatography-tandem mass spectrometry. Endocrinology 156:2492–2502. 10.1210/en.2014-1890 [DOI] [PubMed] [Google Scholar]
- Pelletier G. (2010) Steroidogenic enzymes in the brain: morphological aspects, Ed 1New York: Elsevier. [DOI] [PubMed] [Google Scholar]
- Peinnequin A, Mouret C, Birot O, Alonso A, Mathieu J, Clarencon D, Agay D, Chancerelle Y, Multon E (2004) Rat pro-inflammatory cytokine and cytokine related mRNA quantification by real-time polymerase chain reaction using SYBR green. BMC Immunol 5:3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poletti A, Coscarella A, Negri-Cesi P, Colciago A, Celotti F, Martini L (1998) 5 alpha-reductase isozymes in the central nervous system. Steroids 63:246–251. 10.1016/S0039-128X(98)00018-X [DOI] [PubMed] [Google Scholar]
- Raskin K, de Gendt K, Duittoz A, Liere P, Verhoeven G, Tronche F, Mhaouty-Kodja S (2009) Conditional inactivation of androgen receptor gene in the nervous system: effects on male behavioral and neuroendocrine responses. J Neurosci 29:4461–4470. 10.1523/JNEUROSCI.0296-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupprecht R, Reul JM, Trapp T, van Steensel B, Wetzel C, Damm K, Zieglgänsberger W, Holsboer F (1993) Progesterone receptor-mediated effects of neuroactive steroids. Neuron 11:523–530. 10.1016/0896-6273(93)90156-L [DOI] [PubMed] [Google Scholar]
- Sarkar J, Wakefield S, MacKenzie G, Moss SJ, Maguire J (2011) Neurosteroidogenesis is required for the physiological response to stress: role of neurosteroid-sensitive GABAA receptors. J Neurosci 31:18198–18210. 10.1523/JNEUROSCI.2560-11.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saver JL. (2006) Time is brain–quantified. Stroke 37:263–266. 10.1161/01.STR.0000196957.55928.ab [DOI] [PubMed] [Google Scholar]
- Sayeed I, Stein DG (2009) Progesterone as a neuroprotective factor in traumatic and ischemic brain injury. New York: Elsevier. [DOI] [PubMed] [Google Scholar]
- Sayeed I, Guo Q, Hoffman SW, Stein DG (2006) Allopregnanolone, a progesterone metabolite, is more effective than progesterone in reducing cortical infarct volume after transient middle cerebral artery occlusion. Ann Emerg Med 47:381–389. 10.1016/j.annemergmed.2005.12.011 [DOI] [PubMed] [Google Scholar]
- Schumacher M, Mattern C, Ghoumari A, Oudinet JP, Liere P, Labombarda F, Sitruk-Ware R, De Nicola AF, Guennoun R (2014) Revisiting the roles of progesterone and allopregnanolone in the nervous system: resurgence of the progesterone receptors. Prog Neurobiol 113:6–39. 10.1016/j.pneurobio.2013.09.004 [DOI] [PubMed] [Google Scholar]
- Sierra A, Gottfried-Blackmore A, Milner TA, McEwen BS, Bulloch K (2008) Steroid hormone receptor expression and function in microglia. Glia 56:659–674. 10.1002/glia.20644 [DOI] [PubMed] [Google Scholar]
- Sitruk-Ware R, El-Etr M (2013) Progesterone and related progestins: potential new health benefits. Climacteric 16:69–78. 10.3109/13697137.2013.802556 [DOI] [PubMed] [Google Scholar]
- Smith MS, Freeman ME, Neill JD (1975) The control of progesterone secretion during the estrous cycle and early pseudopregnancy in the rat: prolactin, gonadotropin and steroid levels associated with rescue of the corpus luteum of pseudopregnancy. Endocrinology 96:219–226. 10.1210/endo-96-1-219 [DOI] [PubMed] [Google Scholar]
- Smith-Swintosky VL, Pettigrew LC, Sapolsky RM, Phares C, Craddock SD, Brooke SM, Mattson MP (1996) Metyrapone, an inhibitor of glucocorticoid production, reduces brain injury induced by focal and global ischemia and seizures. J Cereb Blood Flow Metab 16:585–598. 10.1097/00004647-199607000-00008 [DOI] [PubMed] [Google Scholar]
- Traish AM, Wotiz HH (1990) Monoclonal and polyclonal antibodies to human progesterone receptor peptide-(533–547) recognize a specific site in unactivated (8S) and activated (4S) progesterone receptor and distinguish between intact and proteolyzed receptors. Endocrinology 127:1167–1175. 10.1210/endo-127-3-1167 [DOI] [PubMed] [Google Scholar]
- Tronche F, Kellendonk C, Kretz O, Gass P, Anlag K, Orban PC, Bock R, Klein R, Schütz G (1999) Disruption of the glucocorticoid receptor gene in the nervous system results in reduced anxiety. Nat Genet 23:99–103. 10.1038/12703 [DOI] [PubMed] [Google Scholar]
- Türeyen K, Vemuganti R, Sailor KA, Dempsey RJ (2004) Infarct volume quantification in mouse focal cerebral ischemia: a comparison of triphenyltetrazolium chloride and cresyl violet staining techniques. J Neurosci Methods 139:203–207. 10.1016/j.jneumeth.2004.04.029 [DOI] [PubMed] [Google Scholar]
- Vivien D, Gauberti M, Montagne A, Defer G, Touzé E (2011) Impact of tissue plasminogen activator on the neurovascular unit: from clinical data to experimental evidence. J Cereb Blood Flow Metab 31:2119–2134. 10.1038/jcbfm.2011.127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wali B, Ishrat T, Won S, Stein DG, Sayeed I (2014) Progesterone in experimental permanent stroke: a dose-response and therapeutic time-window study. Brain 137:486–502. 10.1093/brain/awt319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wali B, Ishrat T, Stein DG, Sayeed I (2016) Progesterone improves long-term functional and histological outcomes after permanent stroke in older rats. Behav Brain Res 305:46–56. 10.1016/j.bbr.2016.02.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J, Jiang C, Liu C, Li X, Chen N, Hao Y (2010) Neuroprotective effects of progesterone following stroke in aged rats. Behav Brain Res 209:119–122. 10.1016/j.bbr.2010.01.026 [DOI] [PubMed] [Google Scholar]
- Waters EM, Torres-Reveron A, McEwen BS, Milner TA (2008) Ultrastructural localization of extranuclear progestin receptors in the rat hippocampal formation. J Comp Neurol 511:34–46. 10.1002/cne.21826 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westberry JM, Prewitt AK, Wilson ME (2008) Epigenetic regulation of the estrogen receptor alpha promoter in the cerebral cortex following ischemia in male and female rats. Neuroscience 152:982–989. 10.1016/j.neuroscience.2008.01.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams KC, Renthal NE, Condon JC, Gerard RD, Mendelson CR (2012) MicroRNA-200a serves a key role in the decline of progesterone receptor function leading to term and preterm labor. Proc Natl Acad Sci U S A 109:7529–7534. 10.1073/pnas.1200650109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong R, Bath PM, Kendall D, Gibson CL (2013) Progesterone and cerebral ischaemia: the relevance of ageing. J Neuroendocrinol 25:1088–1094. 10.1111/jne.12042 [DOI] [PubMed] [Google Scholar]
- Yousuf S, Sayeed I, Atif F, Tang H, Wang J, Stein DG (2014) Delayed progesterone treatment reduces brain infarction and improves functional outcomes after ischemic stroke: a time-window study in middle-aged rats. J Cereb Blood Flow Metab 34:297–306. 10.1038/jcbfm.2013.198 [DOI] [PMC free article] [PubMed] [Google Scholar]