
Figure 1.
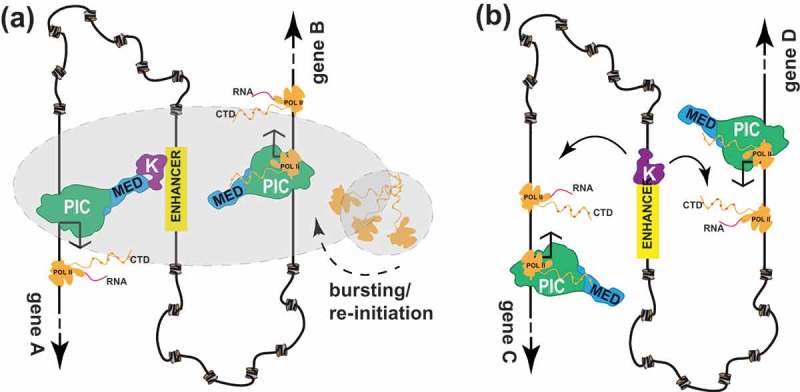
Speculative models for CDK8 or CDK19 module function at mammalian enhancers. a) CDK8 or CDK19 module (“K”) association with an enhancer (e.g. via TF binding) enables its interaction with promoters that are juxtaposed via chromatin loops. This co-localization may be facilitated by liquid-liquid phase separation (LLPS)[137], which is represented by grey shading. At gene A (left), CDK8/19 module–Mediator binding occurs after pol II clears the promoter. Whereas this interaction prevents rapid re-initiation by another pol II complex, promoter-bound CDK8-Mediator can regulate pol II pausing and/or elongation, through physical or functional interactions with the Super-Elongation Complex (SEC) or other factors [17,62]. At gene B (right), transcriptional bursting is depicted[137], in which multiple pol II complexes initiate in succession[70]. This process appears to be Mediator-dependent[148]. The CDK8/19 module does not associate with Mediator at this promoter, as CDK8 module–Mediator binding is mutually exclusive with Mediator–pol II binding [146,147]. Note that at a different point in time, the situation could be switched, with CDK8/19 module association with gene B and gene A undergoing transcriptional bursting. b) An alternative model enables the CDK8/19 module to function independently of Mediator[117]; the enhancer-bound kinase module may regulate transcription elongation via its juxtaposition with elongating pol II complexes at co-localized genes. In support of this model, enhancers have been observed to track with elongating pol II[149], and the CDK8 module appears to positively regulate transcription elongation[17].