

Abstract
Spaced learning has been shown consistently to benefit memory compared with massed learning, yet the neural representations and processes underlying the spacing effect are still poorly understood. In particular, two influential models (i.e., the encoding variability hypothesis and the study-phase retrieval hypothesis) could both model behavioral performance very well, but they make opposite hypotheses regarding the spacing effect's neural mechanisms. The present study attempted to provide empirical neural evidence to adjudicate these competing hypotheses. Using spatiotemporal pattern similarity (STPS) analysis of EEG data, this study investigated whether and how repetition lags (massed/short-spaced/long-spaced) modulated the STPS's contribution to episodic memory encoding in male and female human participants. The results revealed that greater item-specific STPS in the right frontal electrodes at 543–727 ms after stimulus onset was associated with better memory performance. More importantly, this STPS was larger under the spaced-learning condition than the massed-learning condition and partially mediated the spacing effect on memory performance. In addition, we found that massed learning was associated with stronger repetition suppression in the N400 component that reflected momentary retrieval strength, but reduced activity in the late positive component that was associated with memory retrieval. These results suggest that spaced learning improves long-term memory by increasing retrieval effort and enhancing the pattern reinstatement of prior neural representations, which may be achieved by reducing the momentary retrieval strength as the extended repetition lags might help to eliminate the residual representation in working memory.
SIGNIFICANCE STATEMENT As one of the most ubiquitous and fundamental phenomena in the history of memory research, the spacing effect provides an important window into understanding how enduring memory is formed in the brain and how different practice strategies could modulate these mechanisms to affect memory performance. By leveraging the neural representational analysis on scalp EEG data, the current study provides the first empirical data to show that spaced learning enhances memory by improving the spatiotemporal similarity that occurs at a late time window. Our results support the study-phase retrieval hypothesis but not the encoding variability hypothesis and emphasize the role of neural pattern reinstatement in strengthening memory via repeated study.
Keywords: EEG, episodic memory, reinstatement, representational pattern similarity, spacing effect
Introduction
One of the most robust and fundamental phenomena in learning and memory is the spacing effect (Ebbinghaus, 1964; Toppino and Gerbier, 2014). Compared with restudying the material in immediate succession (i.e., massed learning), interleaving repetitions with time or other materials (i.e., spaced learning) benefits memory. The spacing effect has been observed across diverse learning tasks and various learning materials in human and nonhuman species (Cepeda et al., 2006; Gerbier and Toppino, 2015; Smolen et al., 2016). Although many cognitive theories and computational models have been proposed to account for the spacing effect, whether and how the neural representations contribute to it remain unknown.
Among these models, the encoding variability hypothesis makes specific predictions regarding the representational mechanisms of the spacing effect. In particular, it assumes that greater variability across learning repetitions provides more routes to effective retrieval (Estes, 1959). Due to contextual drift over time (Glenberg, 1979), longer inter-repetition intervals (IRIs) would lead to greater contextual change and thus more variable encoding, resulting in better memory performance. Consistent with this hypothesis, introducing variations across massed repetitions has been found to improve memory performance (Paivio et al., 1988; Appleton-Knapp et al., 2005). Nevertheless, variations under the spaced-learning condition are not beneficial and sometimes are even detrimental to memory performance (Verkoeijen et al., 2004; Toppino and Gerbier, 2014).
To account for such results, the study-phase retrieval hypothesis proposes that each repetition serves as a retrieval cue to reactivate and then strengthen the representation of the prior experience (Thios and D'Agostino, 1976). Spacing could reduce the momentary retrieval strength, thus creating greater difficulty in memory retrieval and benefiting later memory (Bjork, 1988). Consistently, items that were not recognized at the second presentation were recalled poorly (Madigan, 1969). Still, the introduction of variations in the spaced condition could lead to retrieval failure and impair memory (Johnston and Uhl, 1976; Verkoeijen et al., 2004).
Studies examining the neural representational mechanisms of memory encoding might help to test these competing hypotheses. The encoding variability hypothesis predicts that better memory is achieved when the representations were more dissimilar across repetitions, and that spacing the repetitions enhances memory by further increasing the dissimilarity. Contrary to these predictions, however, studies using fMRI and representational similarity analysis have found that greater similarity rather than dissimilarity in neural representations across repetitions was associated with better subsequent memory (Xue et al., 2010; Ward et al., 2013; Hasinski and Sederberg, 2016; Zheng et al., 2018). Other studies have further demonstrated that reactivations of prior representations during subsequent learning contribute to successful memory encoding (Kuhl et al., 2010; Lu et al., 2015; Koen and Rugg, 2016).
Two questions remain to be addressed. First, because no study has compared the neural representations under the spaced- and massed-learning conditions, it is still unclear whether spacing could enhance neural pattern reinstatement to improve subsequent memory. Second, although massed learning was associated with larger neural repetition suppression (Wagner et al., 2000; Callan and Schweighofer, 2010; Xue et al., 2011), stronger momentary retrieval strength, and less retrieval processing (Appleton-Knapp et al., 2005; Zhao et al., 2015), the relationships among momentary retrieval strength, retrieval processing, and neural pattern similarity have yet to be established.
The present study used EEG and spatiotemporal pattern similarity analysis (Lu et al., 2015) to address these questions. We chose EEG instead of fMRI for two main reasons. First, due to the temporal autocorrelation of the BOLD response, the neural pattern similarity between two trials is affected by their temporal distance (Mumford et al., 2014), which seriously confounds the spacing effect. Second, EEG provides higher temporal resolution to differentiate 350–450 ms component reflecting retrieval strength (Rugg and Curran, 2007; Zhao et al., 2015), and the 500–700 ms component containing the neural pattern similarity that supports later memory (Lu et al., 2015). By comparing the neural processes and neural pattern similarity for trials under the massed, short-spaced, and long-spaced conditions and linking them to memory performance, our results could help to achieve a deeper mechanistic understanding of the spacing effect in learning.
Materials and Methods
Participants.
Thirty-three healthy Chinese college students (19 females; mean age = 19.8 ± 1.8 years, range 17–25) completed the experiment. All participants were right-handed, had a normal or corrected-to-normal vision, and no history of neurological or psychiatric diseases. Six additional subjects were recruited but excluded from final analysis due to their high rate of “no response” trials (one subject) or noisy EEG data (five subjects). The study was approved by the Institutional Review Board of the State Key Laboratory of Cognitive Neuroscience and Learning at Beijing Normal University.
Materials.
We used 288 pictures of Chinese faces in the learning phase and another 288 faces as foils in the recognition memory test. To minimize the primacy and recency effects, 12 additional faces (four for each run) were added at the beginning and the end of the study lists, but were not tested in the recognition task. All faces were unfamiliar to the subjects and were drawn from the CAS-PEAL face database (Gao et al., 2008). They were presented in the same size (180 × 240 pixels) in the center of the computer screen on a gray background.
Procedure and design.
During the encoding phase, participants were asked to make an age judgment (older or younger than 30 years old) on each presented face by pressing one of two buttons. They were not told about the subsequent memory task. All faces were presented twice within a run, with one-third under the massed (MA) learning condition (i.e., the IRI was 0–1 trials), one-third under the short-spaced (SS) learning condition (i.e., the IRI was 4–8 trials), and the rest under the long-spaced (LS) learning condition (i.e., the IRI was 94–96 trials) (Fig. 1A). The materials used in the three conditions were fully counterbalanced across participants. Each trial started with a fixation cross lasting 500–800 ms (randomly jittered), followed by the presentation of the stimulus for 1000 ms and a blank screen for 2500 ms. Participants were allowed to respond within 2000 ms after stimulus onset. The encoding phase consisted of three runs, each lasting 10 min.
Figure 1.

Experimental paradigm and behavioral results. A, Each novel Chinese face was presented twice under one of the three IRI conditions: MA (0–1 trials), SS (4–8 trials), and LS (94–96 trials). Participants were asked to perform an age judgment task during learning. B, A surprising recognition task was conducted 30 min after the encoding task. Participants were asked to decide whether they recognized each face on a 6-point scale, with 1 indicating “definitely new” and 6 indicating “definitely old.” Spaced learning enhanced memory performance as measured by both high confidence (scored 5 or above) hit rate and d′ (C), reduced repetition priming effect as measured by RT (D), and lower response consistency between the two presentations (E). F, RT was not associated with subsequent memory effect. Error bars indicate within-subject SE. *p < 0.05; ***p < 0.001.
A recognition memory test was conducted after a 30 min visual change detection task that was used as a distractor. During the recognition phase, participants were asked to judge whether they had studied each face earlier on a 6-point scale, with 1 indicating “definitely new” and 6 indicating “definitely old.” In total, 576 faces (288 old faces and 288 new faces) were pseudorandomly mixed and presented one by one over three runs, with the constraint that the numbers of old and new faces were matched within each run. Each trial started with a fixation cross lasting 500–800 ms (randomly jittered), followed by the presentation of the face for up to 3000 ms unless a response was made. A blank screen was then presented until the next trial (Fig. 1B). We used a fixed intertrial interval (ITI) of 4000 ms in the recognition phase so that the total duration of this phase was not affected by participants' response time, which could help to discourage the participants from making hasty responses.
Behavioral data analysis.
For memory performance, the old faces recognized with high confidence (scored 5 or above) were defined as remembered items and those scored 3 or below were defined as forgotten items. These cutoffs were chosen so that there were approximately equal numbers of remembered (42.59%) and forgotten items (40.82%). Both the hit rate and discriminability (d′) were analyzed by condition. Response time (RT) and response consistency (i.e., the rate of same response between two repetitions) in the age judgment task during the encoding phase were analyzed by spacing condition (MA, SS, and LS), subsequent memory (remembered vs forgotten), and, for RT only, by repetition (first vs second presentation). Repeated-measures ANOVA and paired-sample t test were conducted to examine the spacing effect in the encoding task and memory performance.
EEG recording and preprocessing.
Participants were seated ∼100 cm away from the computer screen in a soundproof, light-adjustable room. Continuous EEG data were recorded with a sampling rate of 1024 Hz using the 64-channel ActiveTwo EEG system (Biosemi). Ag-AgCl electrodes were mounted according to the 10–20 system.
EEG data preprocessing was implemented using MATLAB-based toolbox Fieldtrip (RRID:SCR_004849) (Oostenveld et al., 2011) and in-house MATLAB (RRID:SCR_001622) scripts. EEG data were re-referenced to the average of all electrodes, downsampled to the rate of 256 Hz, and filtered with a band-pass filter of 0.5–40 Hz. Eye movements, blinks, and muscle artifacts were identified and corrected using the independent components analysis algorithm. The continuous data were then segmented into epochs from −200 to 1000 ms with regard to stimulus onset. The prestimulus interval (−200 to 0 ms) was used as the baseline for baseline removal procedure. Trials contaminated by any remaining eye movement, blink, or muscle activity were rejected by visual inspection.
Spatiotemporal pattern similarity analysis (STPS).
We constructed spatiotemporal feature vectors from the single-trial epoch data and conducted the spatiotemporal pattern similarity analysis (Lu et al., 2015). The spatial features were scalp voltages from one of the six regions for better spatial specificity (15 channels per region) (Fig. 2A), and the temporal features were selected using a 100 ms sliding window (26 time points) from the epoch data, with a step size of one time point. The similarity between trials was calculated using Pearson correlation. The correlation coefficients were then converted to Fisher's Z scores for subsequent statistical analysis.
Figure 2.

Spaced learning was associated with greater item-specific STPS, which predicted better subsequent memory performance. A, The 64 electrodes were grouped into six regions for better spatial specificity. B, Statistics (F value) of subsequent memory by item specificity (WI vs BI) interaction obtained under the framework of memory by item specificity by spacing three-way ANOVA. The x-axis represents time and the y-axis represents the spatial regions. C, Plots of STPS differences between remembered and forgotten items as a function of WI and BI. The gray-shaded area indicates the temporal cluster showing a significant subsequent memory by item specificity interaction. The cluster remained significant after correcting for multiple comparisons using the cluster-based permutation test. D, Bar graph of the mean STPS in the corresponding temporal cluster in region 2 as a function of subsequent memory separately for WI and BI STPS. E, Plot of item-specific representation (WI − BI) across three spacing conditions, within the cluster showing subsequent memory effect. The orange-shaded area marks the temporal cluster showing a significant spacing (MA vs SS) by item specificity interaction after correcting for multiple comparisons using a cluster-based permutation test. F, Plot of the mean STPS in the shaded temporal cluster in E as a function of item specificity separately for spacing conditions, with SS being associated with better item-specific representations than MA. Error bars indicate within-subject SE. *p < 0.05; **p < 0.01; ***p < 0.001.
The within-item (WI) STPS was obtained by calculating the similarity between two repetitions of the same item. To determine whether the WI similarity reflected item-specific representations or common cognitive processes, we calculated the similarity for between-item (BI) pairs that matched the WI pairs in terms of their memory performance, spacing condition, number of repetitions (one or two), and IRI. Specifically, for a WI pair, we selected a BI pair in which the two trials were from the same spacing condition and showed the same subsequent memory performance as those in the WI pair. In addition, one trial of the pair was the first repetition of an item and the other trial was the second repetition of another item. Finally, the ITI between BI pairs and WI pairs were matched as closely as possible, although we did not find a significant effect of ITI on the pattern similarity of BI pairs (χ(1)2 = 0.21, p = 0.65). A greater WI similarity than BI similarity should reflect item-specific encoding.
In the current study, we aimed to examine the neural representational mechanisms underlying the spacing effect in enhancing episodic memory. As a result, we were particularly interested in the neural differences between spaced and massed condition that were associated with subsequent memory effect. In other words, although there could be many neural differences between the spaced and massed conditions, they might reflect different cognitive functions such as sensory processing or motor control, but such similarities are not theoretically (directly) related to the condition differences in memory performance. Instead, only the condition differences in neural pattern similarity that are linked to successful memory encoding would explain the spacing effect in memory. As a result, we needed first to localize the memory related neural patterns and then test whether spacing modulated them.
To achieve this goal, we did the following hypothesis-driven statistical analyses. First, we located the spatiotemporal windows where the item-specific representation was associated with subsequent memory performance. The WI STPS and BI STPS were separately grouped and averaged across pairs according to the status of subsequent memory performance and spacing condition, separately for each individual participant. Whole-brain three-way ANOVAs were conducted, with spacing condition (MA, SS, and LS), subsequent memory performance (remembered/forgotten), and item specificity (WIs/BIs) as within-subject factors. We particularly focused on the contrast of subsequent memory by item specificity interaction, which should be orthogonal to the spacing effect. In other words, although there were more remembered items in the spaced learning conditions than massed learning condition, the spacing effect should not bias the subsequent memory effect since the remembered and forgotten items were separately averaged within each learning condition before the three-way ANOVA. To make sure that the subsequent memory effect was consistent across spacing conditions, we also examined the subsequent memory by spacing interaction as well as the three-way interaction. Focusing on the time windows showing significant item-specific subsequent memory effects (i.e., significant item specificity by subsequent memory interaction, but no three-way interaction or memory by spacing interaction), we conducted post hoc comparisons to determine whether remembered items showed greater item-specific representations.
To determine the representational mechanisms of the spacing effect on subsequent memory, we then focused on those windows where the item-specific pattern similarity predicted subsequent memory performance to further investigate how spacing affected item-specific representations. In this analysis, the remembered items and forgotten items in each learning condition were pooled together and then averaged, which would allow for a better examination of the spacing effect than would separate averages of remembered and forgotten trials because there were different numbers of remembered items in different spacing conditions. We then conducted item specificity by spacing ANOVA. Focusing on the clusters showing item specificity by spacing interactions, we conducted post hoc comparisons to determine whether spacing could enhance item-specific pattern similarity.
Univariate event-related potential (ERP) analysis.
To detect the ERP components showing the subsequent memory effect, we averaged the EEG responses according to subsequent memory performance, spacing condition, and repetition separately for each individual participant. Similar to the STPS analysis, we first conducted a whole-brain three-way ANOVA, with spacing condition (MA, SS, and LS), subsequent memory performance (remembered/forgotten), and repetition (Rep1/Rep2) as within-subject factors. The ERP components associated with subsequent memory performance were defined as those showing a significant main effect of subsequent memory and no significant interaction with other two factors.
Focusing on these components, we then investigated whether spacing would modulate these components. Again, the remembered items and forgotten items in each learning condition were pooled together and then averaged. Since spacing could only modulate the EEG response evoked by the second repetition, we separately compared MA versus SS, MA versus LS, and SS versus LS on the ERP evoked by the second repetition. In an exploratory analysis, we also investigated whether spacing could modulate the repetition priming effect by a whole-brain spacing by repetition ANOVA.
Nonparametric cluster-based permutation test.
Corrections for multiple comparisons were performed using a nonparametric statistical method based on cluster-level permutation tests implemented in Fieldtrip toolbox. Statistical testing was performed for every time window, and the time windows whose statistical value was larger than a threshold (p = 0.05) were selected and clustered into connected sets on the basis of temporal adjacency. The observed cluster-level statistics were calculated by taking the sum of the statistical values within a cluster. Then, condition labels were permuted 10,000 times based on their exchangeability, and the maximum cluster statistic over all six regions in each permutation was chosen to construct a distribution of the cluster-level statistics under the null hypothesis. The nonparametric statistical test was obtained by calculating the proportion of randomized test statistics that exceeded the observed cluster-level statistics. For the spacing effect, the permutation test was conducted within the pre-defined window showing subsequent memory effect. For univariate analyses on ERPs, the procedures were the same except that the cluster was set on the basis of temporal and spatial adjacency. When pairwise comparisons between the three spacing conditions were conducted, the cluster-level tests were further corrected for multiple comparisons using Bonferroni correction.
Mixed-effects model.
The mixed-effects model is useful for modeling the influence of predictors at multiple levels of variables simultaneously and for jointly modeling both discrete and continuous variables (Gelman and Hill, 2006). It has been used in sophisticated fMRI designs (Ward et al., 2013; Xiao et al., 2017) and single-trial ERP analyses (Valente et al., 2014; Payne et al., 2015). In this study, the mixed-effects model was implemented with lme4 (Bates et al., 2015) in R (RRID:SCR_001905). Participants were included as a random effect. We used the likelihood ratio test to compare the models (with vs without the predictor) to determine the effect of the predictor.
Controlling the effect of univariate ERP amplitude on STPS.
To determine whether our key findings of pattern similarity were due to differences in univariate amplitude, we constructed linear mixed-effects models to examine the subsequent memory effect and spacing effect, by including the amplitude as covariate. In the model for the subsequent memory effect, memory strength (1–6) was used as the dependent variable; the WI pattern similarity in the spatiotemporal clusters showing subsequent memory effect was used as the predictor, and the corresponding mean EEG amplitude was included as the confounding factor. In the model for the spacing effect, the mean WI pattern similarity was used as the dependent variable, the spacing condition was used as the predictor, and the mean EEG amplitude was included as the confounding factor. Participants were included as a random effect.
Mediation analysis.
We performed the mediation effect test to further investigate whether the spacing effect on memory was mediated by the WI pattern similarity, which was related to both spacing condition and subsequent memory performance. Mixed-effects models as implemented by lme4 (Bates et al., 2015) in R (RRID:SCR_001905) were used to test the relationship between spacing and memory strength (1–6 confidence responses were used to index memory strength) (Y = a1 + b1X + ε1; (2) spacing and WI similarity (M = a2 + b2X + ε2); (3) spacing and memory strength with a mediator (Y = a3 + b3X + bM + ε3). In those equations, Y is the dependent variable, X is the predictor, and M is the mediator. The indirect effect was estimated as b2 × b. We used distribution-of-the-product method to compute the confidence interval (CI).
Results
Spaced learning was associated with better memory
Consistent with previous observations, the current study revealed a significant spacing effect on subsequent memory performance (Fig. 1C) (hit rate: F(2,64) = 6, p = 0.004; d′: F(2,64) = 8.63, p = 0.0005). The mean hit rate with high confidence (5 or above) for MA, SS, and LS were 40.08%, 42.94%, and 44.75%, respectively. Compared with MA, the hit rate was significantly higher under SS (t(32) = 2.5, p = 0.02) and LS (t(32) = 3.39, p = 0.002). Similarly, the d′ was also higher under SS (0.53, t(32) = 3.96, p = 0.0004) or LS (0.55, t(32) = 3.46, p = 0.002) than MA (0.41). No significant difference between SS and LS was found for either hit rate (t(32) = −1.18, p = 0.25) or d′ (t(32) = −0.63, p = 0.53).
Spaced learning reduced repetition priming and response consistency
Three-way (spacing condition, subsequent memory performance, and repetition) repeated-measures ANOVA revealed no significant main effect of subsequent memory (remembered vs forgotten) on reaction time (F(1,32) = 0.21, p = 0.65), nor its interactions with other two factors (all p > 0.066) (Fig. 1F). The interaction between repetition (P1 vs P2) and spacing condition (MA, SS, and LS) was significant (F(2,64) = 31.69, p < 0.0001). Further analyses showed that the RTs at the first presentation were comparable across the three spacing conditions (MA: 787 ms, SS: 790 ms, LS: 793 ms; F(2,64) = 0.48, p = 0.62), but the RTs at the second presentation were much shorter under the MA condition (694 ms) than under the SS (766 ms, t(32) = −9.83, p < 0.0001) and LS conditions (781 ms, t(32) = −10.16, p < 0.0001) and slightly shorter under the SS condition than the LS condition (t(32) = −2.19, p = 0.04) (Fig. 1D).
Two-way (spacing condition by subsequent memory performance) repeated-measures ANOVA were then used to analyze the response consistency between the two presentations. Results revealed a significant main effect of spacing condition (F(2,64) = 25, p < 0.0001), but no effect of subsequent memory performance (F(1,32) = 1.64, p = 0.21) or the interaction between spacing condition and subsequent memory performance (F(2,64) = 1.4, p = 0.25). Further paired sample t test revealed significantly higher consistency for MA (82.65%) than SS (76.45%) (t(32) = 4.4, p = 0.0001) and LS (72.41%) (t(32) = 6.85, p < 0.0001) and for SS than LS (t(32) = 3.35, p = 0.0021) (Fig. 1E). These results suggest that spaced learning was associated with weaker repetition priming and lower response consistency during learning, but these factors were not directly associated with subsequent memory performance.
Subsequently remembered items showed greater item-specific STPS
The above analyses revealed a significant spacing effect on subsequent memory performance. To determine the underlying neural mechanism, we tested the hypothesis that spaced learning would enhance memory by increasing item-specific neural pattern similarity that was associated with subsequent memory performance. As the first step, we located the spatiotemporal windows in which the item-specific STPS was associated with subsequent memory performance, using the contrast of subsequent memory by item–specificity interaction in the spacing by subsequent memory by item specificity three-way ANOVA. This analysis revealed a 543–727 ms cluster in right frontal region (region 2) showing a significant subsequent memory by item–specificity interaction (F(1,32)max = 12.84, Fclustersum = 391.75, pcluster = 0.03; Fig. 2B,C). No cluster in the whole brain showed significant spacing by subsequent memory interaction or spacing by subsequent memory by item specificity interaction. Post hoc tests showed significantly greater WI STPS for remembered items than forgotten items (F(1,32) = 10.98, p = 0.002), but no difference in BI STPS (F(1,32) = 0.34, p = 0.56). In addition, significantly greater WI than BI STPS was only found for subsequently remembered items (F(1,32) = 10.8, p = 0.003), but not for forgotten items (F(1,32) = 1.81, p = 0.19) (Fig. 2D). The subsequent memory effect in WI STPS remained significant after controlling the EEG amplitude (χ(1)2 = 7.99, p = 0.005). This finding replicated previous results from a study that used foreign characters as learning stimuli (Lu et al., 2015).
Spaced learning was associated with greater item-specific STPS
Having shown that remembered items were associated with greater item-specific STPS in the late time window, we further tested our core hypothesis that spaced learning improved memory performance by enhancing the item-specific STPS associated with memory performance. Focusing on the right frontal cluster that showed greater item-specific STPS for remembered items than forgotten items, we found a significant item specificity by spacing (MA vs SS) interaction in the 606–652 ms time window (F(1,32)max = 13.52, Fclustersum = 123.89, pcluster = 0.016) (Fig. 2E,F). Post hoc tests revealed that, compared with MA, SS showed significantly greater WI STPS (t(32) = 2.43, p = 0.02), but comparable BI STPS (t(32) = 0.46, p = 0.66) (Fig. 2F). The difference in WI STPS remained significant after controlling the EEG amplitude (χ(1)2 = 4.83, p = 0.028). We also found a 676–710 ms time window that showed a spacing (SS vs LS) by item specificity interaction (F(1,32)max = 9.54, Fclustersum = 70.16, pcluster = 0.036), but it did not survive Bonferroni correction. There was no time window showing significant difference in item specificity between MA and LS.
Within-item STPS partially mediated the spacing effect on memory
The above results suggest that spaced learning could enhance the item-specific STPS's contribution to memory performance by increasing the WI STPS. We further investigated whether the WI STPS indeed mediated the spacing effect on memory. We focused on the spacing effect between MA and SS because no significant difference between MA and LS in item-specific STPS was found. The WI STPS in the right frontal region (606–652 ms), which was associated with both subsequent memory performance and spacing effect, was averaged and used as the mediator. Mediation analysis showed that the WI STPS partially mediated the spacing effect on memory (indirect effect = 0.0047, 95% CI = 0.001–0.0093) (Fig. 3).
Figure 3.
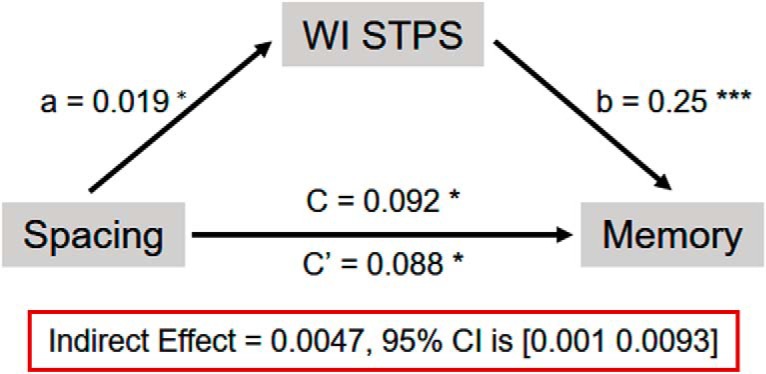
Within-item STPS partially mediated the spacing effect on memory. *p < 0.05; ***p < 0.001.
LPC was associated with the spacing effect on memory
The above analysis tested and supported our hypothesis that spaced learning could enhance memory by increasing item-specific STPS. In addition to STPS, previous studies have found several ERP components during encoding that were associated with subsequent memory performance and were modulated by repetition lags. To determine whether these ERP components could also support the spacing effect in our study, we first compared remembered items and forgotten items to identify memory-related ERPs and further investigated whether those ERPs were modulated by spacing condition. We found that a 500–844 ms response over the occipitoparietal electrodes (i.e., LPC) (Fig. 4A) showed a significant subsequent memory effect (Fclustersum = 6097.4, pcluster = 0.01), with remembered items evoking more positive-going waveforms than forgotten items (Fig. 4D,E). We did not find any cluster showing significant spacing by subsequent memory interaction or spacing by subsequent memory by repetition interaction. Focusing on the LPC, we found that on the second presentation, items under the MA condition evoked less positive-going LPC than those under the SS (672–813 ms, tclustersum = −520.49, pcluster = 0.008) and LS condition (613–766 ms, tclustersum = −703.71, pcluster = 0.002) (Fig. 4F,G). No cluster showed significant difference between SS and LS (all pcluster > 0.13). These results showed that spacing could modulate the LPC amplitudes that were associated with subsequent memory performance.
Figure 4.

Spacing-enhanced LPC responses. A, Topographic map of mean ERP differences between subsequently remembered items and forgotten items during 500–844 ms. B, C, Topographic maps of mean ERP differences between SS and MA (B) and LS and MA (C) on the second presentation during 672–766 ms. The asterisks (*) mark the channels that were used in the STPS analyses. D, Plot of ERP at Pz electrode as a function of memory and repetition. The orange-shaded area marks the temporal cluster showing the subsequent memory effect, with subsequently remembered items evoking more positive going LPC than forgotten items. E, Mean amplitudes of ERP response in the cluster showing the subsequent memory effect. F, Plot of ERP at PZ electrode as a function of repetition and spacing condition. The red line and blue line represent the temporal clusters showing larger amplitude for SS than MA and LS than MA, respectively. G, Bar graph of the mean ERP amplitudes in the cluster showing the spacing effect. H, Bar graph of the LPC STPS as a function of item specificity and subsequent memory. The LPC did not carry item-specific representation. I, Plot of the LPC STPS as a function of item specificity and spacing condition. Error bars indicate within-subject SE. *p < 0.05; **p < 0.01.
LPC reflected memory-related cognitive processes
To further probe the nature of neural representation encoded by the LPC, which was associated with the spacing effect, we applied the spatiotemporal pattern similarity analysis on this component; that is, 672–766 ms time window over the occipitoparietal electrodes (P1, CPz, Pz, POz, and P2). We found that, within this component, there was a significant main effect of subsequent memory; that is, remembered items showed greater pattern similarity than forgotten items (F(1,32) = 6.26, p = 0.018), but no significant effect of item specificity (F(1,32) = 1.97, p = 0.17) or subsequent memory by item–specificity interaction (F(1,32) = 0.002, p = 0.96) (Fig. 4H). In addition, there was no significant main effect of spacing (F(2,64) = 2.23, p = 0.12) or spacing by item–specificity interaction (F(2,64) = 0.37, p = 0.69) (Fig. 4I). These results suggest that the LPC might not carry item-specific representations, but rather may reflect general memory-related cognitive processes.
The above results suggest that both the LPC that reflects general cognitive processing and the item-specific STPS in the right frontal region were associated with the spacing effect on memory. Interestingly, they occurred at a similar time window but with different topographic distributions. Does LPC response contribute to the neural pattern reinstatement? We conducted a mixed-effects model analysis to determine the association between LPC amplitude and the degree of STPS. The result revealed a strong positive association between the LPC response of the second presentation and the WI STPS (χ(1)2 = 128.16, p < 0.0001). The correlation remained significant (χ(1)2 = 74.74, p < 0.0001) after controlling for the spacing condition, memory strength, and the mean amplitude of right frontal region. These results suggest that the LPC may contribute to the neural pattern reinstatement in the right frontal region. Due to the exploratory nature of the analysis on the relationship between ERP response and neural representation, more investigations are required to replicate these results and to further examine the nature of this correlation.
Spaced learning reduced the repetition effect on N400
In addition to more positive LPC, spaced learning also reduced the repetition effect on N400 in the central region as revealed by a significant interaction between spacing condition and repetition (300–484 ms, Fclustersum = 20845, pcluster = 0.0002) (Fig. 5A), though this cluster did not show a significant subsequent memory effect. Post hoc t tests revealed a significant repetition effect under the MA condition (t(32) = 6.79, p < 0.0001), but not under the SS condition (t(32) = 0.08, p = 0.94) or LS condition (t(32) = 0.14, p = 0.89) (Fig. 5D,E). Moreover, mixed-effects analysis suggested that the strength of N400 repetition suppression could predict the behavioral repetition priming effect under the MA condition (χ(1)2 = 14.54, p < 0.0001, β = 0.0047), but could not predict subsequent memory performance (χ(1)2 = 0.2, p = 0.66).
Figure 5.

Spacing reduced the repetition effect on N400. A, Topographic map of mean ERP differences between P2 and P1 under MA during 300–484 ms. B, C, Topographic maps of mean ERP differences between SS and MA (B) and LS and MA (C) on the second presentation. The asterisks (*) mark the channels that were used in the STPS analyses. D, Plot of ERP at Cz electrode as a function of spacing condition and repetition. The orange shaded area marks the temporal cluster showing a significant interaction between spacing condition and repetition. E, Mean amplitudes of N400 response as a function of repetition and spacing condition. F, Plot of WI and BI STPS in N400. G, STPS for remembered (Rem) and forgotten (Forg) items in the N400, which did not show any subsequent memory effect. Error bars indicate within-subject SE. ***p < 0.001.
We also conducted the STPS analyses for this component; that is, a 300–484 ms time window over the central electrodes (FC1, FCz, FC2, C1, Cz, C2, CP1, CPz, and CP2). We did not find any significant main effects or interactions (all p > 0.17) (Fig. 5F,G).
Discussion
The present study used EEG and representational analysis to determine the representational mechanisms underlying the spacing effect. Using novel faces as stimuli, the current study revealed that the item-specific STPS that occurred at the 543–727 ms time window over the right frontal scalp was greater for subsequently remembered items than forgotten items. This result replicated previous findings using novel foreign characters (Lu et al., 2015), suggesting that the role of STPS in memory is not affected by the choice of learning stimulus. Together with fMRI studies (Xue et al., 2010; Visser et al., 2013; Ward et al., 2013), existing evidence across various stimulus types, experimental tasks, repetition lags, and imaging techniques converges to emphasize the critical role of neural pattern similarity in supporting durable memory (Xue, 2018).
More importantly, this memory-related STPS was larger under the spaced learning condition than the massed learning condition and it partially mediated the spacing effect on memory. This contradicts the encoding variability hypothesis that greater dissimilarity benefits memory. Given the temporal contextual drift (Glenberg, 1979), how could spaced learning generate stronger pattern similarity than massed learning? According to the context maintenance and retrieval model (Polyn et al., 2009; Siegel and Kahana, 2014), when the first presentation was retrieved, its context could be reinstated (Manning et al., 2011). This context reinstatement could partially counteract the temporal context drift. Moreover, under the massed condition, the neural representation of the first learning experience might be still activated in the short-term memory system when the IRI is short (Raaijmakers, 2003; Van Strien et al., 2007). According to the new theory of disuse (Bjork and Bjork, 1992; Bjork, 1999), an item in memory can be characterized by two “strengths”: Storage strength reflects how well an item is learned and retrieval strength represents how accessible an item can be via recall or recognition. Under the massed learning condition, items are readily accessible and the strong momentary retrieval strength would reduce the memory retrieval processes, resulting in less gain in storage strength (Zhao et al., 2015).
By combining ERP and spatiotemporal pattern analysis, the current study revealed three lines of neural evidence to support the above hypotheses. First, consistent with many previous studies (Van Strien et al., 2007; Zhao et al., 2015; Manuel and Schnider, 2016), we found greater behavioral repetition priming effect and significant N400 repetition suppression under the massed condition than under the short- or long-spaced condition. These results are also consistent with the fMRI evidence of stronger neural repetition suppression in the sensory and frontoparietal regions under the MA learning condition (Callan and Schweighofer, 2010; Xue et al., 2010, 2011). Consistent with a previous fMRI study (Ward et al., 2013), we also found that the N400 repetition effect could predict the behavioral repetition priming, but not episodic memory performance.
Second, our results are consistent with previous studies showing that the N400 increased with repetitions (Henson et al., 2003; Schweinberger and Neumann, 2016) and reflected the momentary retrieval strength (Zhao et al., 2015). In addition, previous studies also suggested that the N400 supported the familiarity judgment (Curran and Cleary, 2003; Rugg and Curran, 2007; Kutas and Federmeier, 2011). The enhanced N400 response under the massed condition suggested that massed learning was associated with stronger momentary retrieval strength (Van Strien et al., 2007).
Third, the LPC representation contained no item-specific information but was associated with memory performance. Moreover, the LPC showed reduced response with repetitions and its amplitude was associated with the strength of memory-related STPS. Previous studies have shown that LPC is related to recollection (Rugg and Yonelinas, 2003; Rugg and Curran, 2007; Kappenman and Luck, 2012), in particular, the retrieval of complex information about prior events. These results suggest that the LPC is associated with general memory-retrieval processes that contribute to pattern reinstatement. Due to the stronger momentary retrieval strength under the massed condition, the requirement for memory retrieval of prior learning and the reactivation of prior memory trace are impaired.
These results also suggest that the representations in early and late time windows might carry distinct information and have a differential effect on long-term memory formation. A recent study combining MEG and fMRI suggests that, whereas the representation in the early time window corresponded to the representations in the early visual cortex, the representations in the later time window corresponded to the representations in the higher-order visual cortex (Cichy et al., 2014). Item-specific representation in the visual cortex was only found during perception, whereas item-specific representation during memory retrieval was found in the inferior parietal lobule (Xiao et al., 2017; Favila et al., 2018) and its representation was more abstract (Jeong and Xu, 2016; Ye et al., 2016) and more aligned to current goals (Favila et al., 2018). Because perceptual details might decay more rapidly than more abstract information such as semantics (Craik and Tulving, 1975), the early pattern similarity might reflect more sensory information that did not predict later memory, whereas the later item-specific pattern similarity might reflect more abstract information and contribute to long-term memory. Future studies should further investigate the nature of representation and their anatomical origins of the early and late components, which would help to deepen our understanding of the role of cortical representation in memory formation.
These findings could also well account for the differential effect of encoding variability on memory performance. According to the differentiation model of memory (Kılıç et al., 2017), repeated exposure to an item results in the storage of additional information in the single memory trace established during the first exposure. As the memory trace is updated, its similarity to other items decreases and becomes more distinguishable during retrieval. Introducing variance in the massed repetitions could reduce the momentary retrieval strength, enhance the retrieval of later abstract representations, and improve memory. However, introducing variance under the spaced condition could increase the chance of retrieval failure (Verkoeijen et al., 2005; Cepeda et al., 2006), in which case a new trace might be formed. When the study-phase retrieval failed, the probability of retrieving the old trace at the final test would be even lower. Therefore, the probability of recalling the item mostly depended on the new trace and the performance is thus impaired (Raaijmakers, 2003). This mechanism could also account for the inverted U-shaped relationship between lag and memory (Verkoeijen et al., 2005; Cepeda et al., 2006), as too long a lag could also result in retrieval failure. Finally, it could also account for the superadditivity effect, i.e., observed benefit of repetition on memory strength is higher than that predicted by the encoding variability hypothesis, since the representations for the two study events should not be considered as independent (Benjamin and Tullis, 2010).
The present study failed to replicate the detrimental effect of long IRI. Some behavioral studies found that the spacing effect could be effective in terms of days or months (Cepeda et al., 2008). In addition, the optimal interval also varied in different learning tasks, different repetition intervals, and different retention lags (Cepeda et al., 2006; Pashler et al., 2009; Toppino and Gerbier, 2014). In the current study, we tried the longest possible IRI with the constraint that trials from each condition were all within one session (to avoid any cross-session differences). It seems that this long-spaced interval might not be long enough to bend the curve. In addition, this design unavoidably results in an imbalance in the temporal distributions of repetitions under different spacing conditions, which might have some effect on memory performance. Indeed, we found a significant spacing (MA vs SS) by serial position interaction (F(3,96) = 2.74, p = 0.048). There were significant primacy and recency effects under the MA learning condition; that is, trials studied in the first and last quartiles of a run were remembered better than those studied in the middle (t(32) = 2.33, p = 0.026). However, no such effect was found in the SS condition (t(32) = 0.86, p = 0.4). This suggests that the serial position and spacing might interact to affect memory performance, which could potentially confound the result. Future studies could use a multiday design to determine whether pattern similarity plays a role in the inverted U-shaped relationship of spacing and memory.
Although our results suggest that repetition priming might be a result of residual working memory representation, which might hinder later pattern reinstatement and impair memory, either behavioral repetition priming or N400 repetition suppression could not quantitatively predict memory performance. In fact, many existing studies investigating this issue have found mixed results (Zhao et al., 2015). Due to the fact that the repetition priming is affected by the initial processing strength and the repetition lag, and in turn, would affect the retrieval of prior representation and further encoding, it would be difficult to establish a direct association between repetition priming and memory performance (Xue et al., 2011). Future studies should further investigate the relationship among repetition priming, neural repetition suppression, pattern reinstatement, and episodic memory performance.
In conclusion, our study provides several lines of novel neural evidence to advance our understanding of the mechanisms of repeated studies in enhancing memory. Contrary to the encoding variability hypothesis, better memory was associated with greater STPS that occurred at a late time window and spacing enhanced memory by increasing the STPS. This pattern similarity in the late window might reflect the reinstatement of more abstract representation, which was reduced under the massed condition due to the strong momentary retrieval strength. These results highlight the complex interactions of multiple memory processes and representations in determining memory performance.
Footnotes
This work was sponsored by the National Science Foundation of China (NSFC Grants 31730038 and 31861143040), the 111 Project (Grant B07008), the NSFC and the German Research Foundation (DFG) joint project NSFC 61621136008/DFG TRR-169, and the Guangdong Pearl River Talents Plan Innovative and Entrepreneurial Team (Grant 2016ZT06S220).
The authors declare no competing financial interests.
References
- Appleton-Knapp SL, Bjork RA, Wickens TD (2005) Examining the spacing effect in advertising: encoding variability, retrieval processes, and their interaction. J Consum Res 32:266–276. 10.1086/432236 [DOI] [Google Scholar]
- Bates D, Mächler M, Bolker B, Walker S (2015) Fitting Linear Mixed-Effects Models Usinglme4. Journal of Statistical Software. Available at https://www.jstatsoft.org/article/view/v067i01.
- Benjamin AS, Tullis J (2010) What makes distributed practice effective? Cognitive psychology 61:228–247. 10.1016/j.cogpsych.2010.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjork RA. (1988) Retrieval practice and the maintenance of knowledge. Practical aspects of memory: Current Research and Issues 1:396–401. [Google Scholar]
- Bjork RA. (1999) Assessing our own competence: Heuristics and illusions. In: Attention and performance XVII: Cognitive regulation of performance: Interaction of theory and application (Gopher D, Koriat A, eds), pp 435–459. Cambridge, MA: The MIT Press. [Google Scholar]
- Bjork RA, Bjork EL (1992) A new theory of disuse and an old theory of stimulus fluctuation. In: Essays in honor of William K. Estes, Vol 1: From learning theory to connectionist theory; Vol 2: From learning processes to cognitive processes. (Healy AF, Kosslyn SM, Shiffrin RM, eds), pp 35–67. Hillsdale, NJ: Lawrence Erlbaum Associates, Inc. [Google Scholar]
- Callan DE, Schweighofer N (2010) Neural correlates of the spacing effect in explicit verbal semantic encoding support the deficient-processing theory. Hum Brain Mapp 31:645–659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cepeda NJ, Pashler H, Vul E, Wixted JT, Rohrer D (2006) Distributed practice in verbal recall tasks: a review and quantitative synthesis. Psychol Bull 132:354–380. 10.1037/0033-2909.132.3.354 [DOI] [PubMed] [Google Scholar]
- Cepeda NJ, Vul E, Rohrer D, Wixted JT, Pashler H (2008) Spacing effects in learning: a temporal ridgeline of optimal retention. Psychol Sci 19:1095–1102. 10.1111/j.1467-9280.2008.02209.x [DOI] [PubMed] [Google Scholar]
- Cichy RM, Pantazis D, Oliva A (2014) Resolving human object recognition in space and time. Nat Neurosci 17:455–462. 10.1038/nn.3635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craik FI, Tulving E (1975) Depth of processing and the retention of words in episodic memory. J Exp Psychol Gen 104:268–294. 10.1037/0096-3445.104.3.268 [DOI] [Google Scholar]
- Curran T, Cleary AM (2003) Using ERPs to dissociate recollection from familiarity in picture recognition. Cogn Brain Res 15:191–205. 10.1016/S0926-6410(02)00192-1 [DOI] [PubMed] [Google Scholar]
- Ebbinghaus H. (1964) Memory: A contribution to experimental psychology. New York: Dover Publications. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Estes WK. (1959) The statistical approach to learning theory. Psychology: A study of a science 2:380–491. [Google Scholar]
- Favila SE, Samide R, Sweigart SC, Kuhl BA (2018) Parietal representations of stimulus features are amplified during memory retrieval and flexibly aligned with top-down goals. J Neurosci 38:7809–7821. 10.1523/JNEUROSCI.0564-18.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao W, Cao B, Shan S, Chen X, Zhou D, Zhang X, Zhao D (2008) The CAS-PEAL large-scale chinese face database and baseline evaluations. IEEE Transactions on Systems, Man, and Cybernetics-Part A: Systems and Humans 38:149–161. 10.1109/TSMCA.2007.909557 [DOI] [Google Scholar]
- Gelman A, Hill J (2006) Data analysis using regression and multilevel/hierarchical models. Cambridge: Cambridge University. [Google Scholar]
- Gerbier E, Toppino TC (2015) The effect of distributed practice: neuroscience, cognition, and education. Trends in Neuroscience and Education 4:49–59. 10.1016/j.tine.2015.01.001 [DOI] [Google Scholar]
- Glenberg AM. (1979) Component-levels theory of the effects of spacing of repetitions on recall and recognition. Mem Cognit 7:95–112. 10.3758/BF03197590 [DOI] [PubMed] [Google Scholar]
- Hasinski AE, Sederberg PB (2016) Trial-level information for individual faces in the fusiform face area depends on subsequent memory. Neuroimage 124:526–535. 10.1016/j.neuroimage.2015.08.065 [DOI] [PubMed] [Google Scholar]
- Henson RN, Goshen-Gottstein Y, Ganel T, Otten LJ, Quayle A, Rugg MD (2003) Electrophysiological and haemodynamic correlates of face perception, recognition and priming. Cereb Cortex 13:793–805. 10.1093/cercor/13.7.793 [DOI] [PubMed] [Google Scholar]
- Jeong SK, Xu Y (2016) Behaviorally relevant abstract object identity representation in the human parietal cortex. J Neurosci 36:1607–1619. 10.1523/JNEUROSCI.1016-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnston WA, Uhl CN (1976) The contributions of encoding effort and variability to the spacing effect on free recall. Journal of Experimental Psychology: Human Learning and Memory 2:153 10.1037/0278-7393.2.2.153 [DOI] [Google Scholar]
- Kappenman ES, Luck SJ (2012) The Oxford handbook of event-related potential components. New York, US: Oxford University Press. [Google Scholar]
- Kılıç A, Criss AH, Malmberg KJ, Shiffrin RM (2017) Models that allow us to perceive the world more accurately also allow us to remember past events more accurately via differentiation. Cogn Psychol 92:65–86. 10.1016/j.cogpsych.2016.11.005 [DOI] [PubMed] [Google Scholar]
- Koen JD, Rugg MD (2016) Memory reactivation predicts resistance to retroactive interference: evidence from multivariate classification and pattern similarity analyses. J Neurosci 36:4389–4399. 10.1523/JNEUROSCI.4099-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhl BA, Shah AT, DuBrow S, Wagner AD (2010) Resistance to forgetting associated with hippocampus-mediated reactivation during new learning. Nat Neurosci 13:501–506. 10.1038/nn.2498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutas M, Federmeier KD (2011) Thirty years and counting: finding meaning in the N400 component of the event-related brain potential (ERP). Annu Rev Psychol 62:621–647. 10.1146/annurev.psych.093008.131123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu Y, Wang C, Chen C, Xue G (2015) Spatiotemporal neural pattern similarity supports episodic memory. Curr Biol 25:780–785. 10.1016/j.cub.2015.01.055 [DOI] [PubMed] [Google Scholar]
- Madigan SA. (1969) Intraserial repetition and coding processes in free recall. J Verbal Learning Verbal Behav 8:828–835. 10.1016/S0022-5371(69)80050-2 [DOI] [Google Scholar]
- Manning JR, Polyn SM, Baltuch GH, Litt B, Kahana MJ (2011) Oscillatory patterns in temporal lobe reveal context reinstatement during memory search. Proc Natl Acad Sci U S A 108:12893–12897. 10.1073/pnas.1015174108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manuel AL, Schnider A (2016) Differential processing of immediately repeated verbal and non-verbal stimuli: an evoked-potential study. Eur J Neurosci 43:89–97. 10.1111/ejn.13114 [DOI] [PubMed] [Google Scholar]
- Mumford JA, Davis T, Poldrack RA (2014) The impact of study design on pattern estimation for single-trial multivariate pattern analysis. Neuroimage 103:130–138. 10.1016/j.neuroimage.2014.09.026 [DOI] [PubMed] [Google Scholar]
- Oostenveld R, Fries P, Maris E, Schoffelen JM (2011) FieldTrip: open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput Intell Neurosci 2011:156869. 10.1155/2011/156869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paivio A, Clark JM, Lambert WE (1988) Bilingual dual-coding theory and semantic repetition effects on recall. J Exp Psychol Learn Mem Cogn 14:163 10.1037/0278-7393.14.1.163 [DOI] [Google Scholar]
- Pashler H, Cepeda N, Lindsey RV, Vul E, Mozer MC (2009) Predicting the optimal spacing of study: A multiscale context model of memory. In: Advances in neural information processing systems 22 (Bengio Y, Schuurmans D, Lafferty J, Williams C, Culotta A, eds), pp 1321–1329. New York: Curran Associates Inc. [Google Scholar]
- Payne BR, Lee CL, Federmeier KD (2015) Revisiting the incremental effects of context on word processing: evidence from single-word event-related brain potentials. Psychophysiology 52:1456–1469. 10.1111/psyp.12515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polyn SM, Norman KA, Kahana MJ (2009) A context maintenance and retrieval model of organizational processes in free recall. Psychol Rev 116:129–156. 10.1037/a0014420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raaijmakers JGW. (2003) Spacing and repetition effects in human memory: application of the SAM model. Cogn Sci 27:431–452. 10.1207/s15516709cog2703_5 [DOI] [Google Scholar]
- Rugg MD, Curran T (2007) Event-related potentials and recognition memory. Trends Cogn Sci 11:251–257. 10.1016/j.tics.2007.04.004 [DOI] [PubMed] [Google Scholar]
- Rugg MD, Yonelinas AP (2003) Human recognition memory: a cognitive neuroscience perspective. Trends Cogn Sci 7:313–319. 10.1016/S1364-6613(03)00131-1 [DOI] [PubMed] [Google Scholar]
- Schweinberger SR, Neumann MF (2016) Repetition effects in human ERPs to faces. Cortex 80:141–153. 10.1016/j.cortex.2015.11.001 [DOI] [PubMed] [Google Scholar]
- Siegel LL, Kahana MJ (2014) A retrieved context account of spacing and repetition effects in free recall. J Exp Psychol Learn Mem Cogn 40:755–764. 10.1037/a0035585 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smolen P, Zhang Y, Byrne JH (2016) The right time to learn: mechanisms and optimization of spaced learning. Nat Rev Neurosci 17:77–88. 10.1038/nrn.2015.18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thios SJ, D'Agostino PR (1976) Effects of repetition as a function of study phase retrieval. J Verbal Learning Verbal Behav 15:529–536. 10.1016/0022-5371(76)90047-5 [DOI] [Google Scholar]
- Toppino TC, Gerbier E (2014) About practice: Repetition, spacing, and abstraction. In: The psychology of learning and motivation (Ross BH, ed), Vol 60, pp 113–189. San Diego, CA: Elsevier Academic Press. [Google Scholar]
- Valente A, Bürki A, Laganaro M (2014) ERP correlates of word production predictors in picture naming: a trial by trial multiple regression analysis from stimulus onset to response. Front Neurosci 8:390. 10.3389/fnins.2014.00390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Strien JW, Verkoeijen PP, Van der Meer N, Franken IH (2007) Electrophysiological correlates of word repetition spacing: ERP and induced band power old/new effects with massed and spaced repetitions. Int J Psychophysiol 66:205–214. 10.1016/j.ijpsycho.2007.07.003 [DOI] [PubMed] [Google Scholar]
- Verkoeijen PPJL, Rikers RMJP, Schmidt HG (2004) Detrimental influence of contextual change on spacing effects in free recall. J Exp Psychol Learn Mem Cogn 30:796–800. 10.1037/0278-7393.30.4.796 [DOI] [PubMed] [Google Scholar]
- Verkoeijen PP, Rikers RM, Schmidt HG (2005) Limitations to the spacing effect: demonstration of an inverted u-shaped relationship between inter-repetition spacing and free recall. Exp Psychol 52:257–263. 10.1027/1618-3169.52.4.257 [DOI] [PubMed] [Google Scholar]
- Visser RM, Scholte HS, Beemsterboer T, Kindt M (2013) Neural pattern similarity predicts long-term fear memory. Nat Neurosci 16:388–390. 10.1038/nn.3345 [DOI] [PubMed] [Google Scholar]
- Wagner AD, Maril A, Schacter DL (2000) Interactions between forms of memory: when priming hinders new episodic learning. J Cogn Neurosci 12:52–60. 10.1162/089892900564064 [DOI] [PubMed] [Google Scholar]
- Ward EJ, Chun MM, Kuhl BA (2013) Repetition suppression and multi-voxel pattern similarity differentially track implicit and explicit visual memory. J Neurosci 33:14749–14757. 10.1523/JNEUROSCI.4889-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao X, Dong Q, Gao J, Men W, Poldrack RA, Xue G (2017) Transformed neural pattern reinstatement during episodic memory retrieval. J Neurosci 37:2986–2998. 10.1523/JNEUROSCI.2324-16.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue G. (2018) The neural representations underlying human episodic memory. Trends Cogn Sci 22:544–561. 10.1016/j.tics.2018.03.004 [DOI] [PubMed] [Google Scholar]
- Xue G, Dong Q, Chen C, Lu Z, Mumford JA, Poldrack RA (2010) Greater neural pattern similarity across repetitions is associated with better memory. Science 330:97–101. 10.1126/science.1193125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue G, Mei L, Chen C, Lu ZL, Poldrack R, Dong Q (2011) Spaced learning enhances subsequent recognition memory by reducing neural repetition suppression. J Cogn Neurosci 23:1624–1633. 10.1162/jocn.2010.21532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye Z, Zhu B, Zhuang L, Lu Z, Chen C, Xue G (2016) Neural global pattern similarity underlies true and false memories. J Neurosci 36:6792–6802. 10.1523/JNEUROSCI.0425-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao X, Wang C, Liu Q, Xiao X, Jiang T, Chen C, Xue G (2015) Neural mechanisms of the spacing effect in episodic memory: a parallel EEG and fMRI study. Cortex 69:76–92. 10.1016/j.cortex.2015.04.002 [DOI] [PubMed] [Google Scholar]
- Zheng L, Gao Z, Xiao X, Ye Z, Chen C, Xue G (2018) Reduced fidelity of neural representation underlies episodic memory decline in normal aging. Cereb Cortex 28:2283–2296. 10.1093/cercor/bhx130 [DOI] [PubMed] [Google Scholar]