

Summary
Bacteria of the genus Vibrio are common members of aquatic environments where they compete with other prokaryotes and defend themselves against grazing predators. A macromolecular protein complex called the type VI secretion system (T6SS) is used for both purposes. Previous research showed that the sole T6SS of the human pathogen V. cholerae is induced by extracellular (chitin) or intracellular (low c‐di‐GMP levels) cues and that these cues lead to distinctive signalling pathways for which the proteins TfoX and TfoY serve as master regulators. In this study, we tested whether the TfoX‐ and TfoY‐mediated regulation of T6SS, concomitantly with natural competence or motility, was conserved in non‐cholera Vibrio species, and if so, how these regulators affected the production of individual T6SSs in double‐armed vibrios. We show that, alongside representative competence genes, TfoX regulates at least one T6SS in all tested Vibrio species. TfoY, on the other hand, fostered motility in all vibrios but had a more versatile T6SS response in that it did not foster T6SS‐mediated killing in all tested vibrios. Collectively, our data provide evidence that the TfoX‐ and TfoY‐mediated signalling pathways are mostly conserved in diverse Vibrio species and important for signal‐specific T6SS induction.
Introduction
Secretion systems that enable the export of macromolecules across membranes are often associated with pathogenesis in Gram‐negative bacteria. However, these machineries and their substrates also play important roles in the interactions between bacteria and other microorganisms in their natural environment (Costa et al., 2015; Green and Mecsas, 2016). An important secretion system, the type VI secretion system (T6SS), was first described in Vibrio cholerae and is a syringe‐like killing device involved in virulence and interbacterial warfare (Pukatzki et al., 2006; Ho et al., 2014; Russell et al., 2014). T6SSs are present in ∼25% of sequenced Gram‐negative bacteria, including pathogenic and nonpathogenic species, and are defined by a set of 13 core components (Das and Chaudhuri, 2003; Bingle et al., 2008; Boyer et al., 2009). These components build a macromolecular complex that shares both structural and functional homology with contractile bacteriophage tails, such as in the phage T4 (Leiman et al., 2009; Basler, 2015; Taylor et al., 2018). Briefly, a tail‐like structure, which is composed of hemolysin coregulated protein (Hcp) hexamers and sharpened by spike‐like tip proteins (VgrG and PAAR), is surrounded by a contractile sheath structure and polymerizes onto a membrane‐spanning multiprotein complex. When the sheath contracts, the effector‐decorated tip complex is propelled outwards together with the inner tube to puncture the membrane of a neighbouring target cell and inject toxic effector proteins (reviewed by Zoued et al., 2014; Cianfanelli et al., 2016). Each T6SS encodes its own sets of variable effector proteins that determine the specificity and the biological function of the system, as they can impact both eukaryotic host cells and bacterial competitors. To prevent self‐intoxication and to kin‐discriminate siblings, bacteria produce effector‐specific neutralizing immunity proteins, which are frequently encoded adjacent to the effector gene in a bicistronic unit (Durand et al., 2014; Alcoforado Diniz et al., 2015). The structural components of the T6SS are mostly encoded within larger gene clusters, which are variable with respect to genetic content and organization. The genes coding for the secreted core components (Hcp, VgrG, PAAR) are often found in smaller auxiliary clusters together with the genes that encode the effectors‐immunity pairs (Boyer et al., 2009). Many bacterial genomes encode more than one T6SS, which likely serve different needs (Boyer et al., 2009). Accordingly, bacteria employ a wide variety of regulatory mechanisms, from common cues to specific regulatory cascades, to ensure appropriate expression of T6SS genes (Miyata et al., 2013), which likely vary within species when multiple T6SS systems are present. Studying the various T6SS systems and their regulation is, therefore, important as such knowledge sheds light onto the evolution of bacterial species in response to new niches and their interaction with environmental hosts.
V. cholerae O1 El Tor strains, which are responsible for the current seventh pandemic of cholera (further referred to as pandemic strains), harbour a single T6SS. In contrast, many non‐cholera Vibrio species encode several T6SSs, such as clinical isolates of V. parahaemolyticus and environmental isolates of V. vulnificus, both of which carry two T6SSs (Yu et al., 2012; Church et al., 2016). The genome of the squid symbiont V. fischeri, on the other hand, codes for one T6SS even though a recent study by Speare et al. identified a secondary T6SS in several V. fischeri isolates that fostered niche domination within the squid's light organ (Speare et al., 2018).
The T6SS of pandemic V. cholerae is silent under standard laboratory conditions (Pukatzki et al., 2006). In contrast to pandemic strains, the O37 serogroup strains V52 and ATCC25872 are toxigenic but nonpandemic and their T6SS is constitutively activated, as is the case for most environmental isolates (Pukatzki et al., 2006; Unterweger et al., 2012; Bernardy et al., 2016; Van der Henst et al., 2018). To explain the differences in T6SS activity and to provide insight into the T6SS's biological functions, it is, therefore, important to understand the underlying regulatory pathways in pandemic V. cholerae strains. Accordingly, several minor and major T6SS regulators have been identified in V. cholerae (Joshi et al., 2017) by changing their abundance through deletion or forced expression of the respective genes, which resulted in changes in T6SS gene expression or T6SS activity. Two of these major regulators, TfoX and TfoY (Borgeaud et al., 2015; Metzger et al., 2016), contain TfoX‐like N‐ and C‐terminal domains and proteins containing such domains are usually annotated as regulators of natural competence for transformation due to the presence of these domains in the competence regulator Sxy of Haemophilus influenzae (Redfield, 1991) and TfoX in V. cholerae (Meibom et al., 2005). We recently showed that TfoX induces the T6SS in parallel with the competence machinery at a high bacterial cell density (measured by quorum sensing [QS] and signalled through its master regulator, HapR) on chitinous surfaces, leading to T6SS‐mediated killing followed by the absorption of prey‐released DNA (Borgeaud et al., 2015). Coupling kin‐discriminating neighbour killing and competence fosters horizontal gene transfer and, ultimately, evolution (Veening and Blokesch, 2017).
Interestingly, V. cholerae and most other members of the genus Vibrio (vibrios) contain a TfoX homologue named TfoY (Pollack‐Berti et al., 2010; Metzger et al., 2016). However, despite the homology to TfoX and its common annotation as a competence regulator, TfoY is neither essential nor sufficient for competence induction in V. cholerae (Metzger et al., 2016). TfoY also induces the T6SS of V. cholerae, though the TfoX and TfoY regulons are nonoverlapping, with TfoX co‐inducing competence and TfoY orchestrating a defensive reaction. This defensive reaction includes enhanced motility and the co‐production of the T6SS with additional extracellular enzymes, some of which are known to intoxicate amoebal predators (Metzger et al., 2016; Van der Henst et al., 2018). Moreover, TfoX is naturally produced upon growth on chitinous surfaces (Meibom et al., 2005), while TfoY production is inhibited by c‐di‐GMP, an secondary messenger in bacteria, such that it is only present at low intracellular c‐di‐GMP levels (Inuzuka et al., 2016; Metzger et al., 2016).
Given the presence of TfoX and TfoY in many vibrios, we aimed to understand their role in non‐cholera species. We show that the T6SS activation is highly conserved across vibrios, with the majority of tested species concomitantly inducing natural competence and motility when TfoX and TfoY, respectively, are produced. Interestingly, we observed different scenarios when comparing those Vibrio species that encode more than one T6SS, with TfoX and TfoY not fulfilling the same inducing role for all T6SSs. We, therefore, conclude that some vibrios specifically induce one or the other T6SS under different environmental conditions driven by either TfoX or TfoY, while other species combine forces to simultaneously induce both T6SSs.
Results and discussion
Conservation of T6SS production by TfoX and TfoY in pandemic isolates of V. cholerae
Since its discovery in a nonpandemic strain of V. cholerae, it has been reported that the T6SS of pandemic strains is silent under standard laboratory conditions (Pukatzki et al., 2006). We recently showed that the two regulatory proteins TfoX and TfoY foster T6SS production and, accordingly, interbacterial competition in pandemic V. cholerae (Borgeaud et al., 2015; Metzger et al., 2016). The previous study that addressed TfoY, however, was based on a single strain of V. cholerae, namely strain A1552 (Yildiz and Schoolnik, 1998). This human isolate originated from food‐borne transmission on an airplane returning to the US from Peru/South America (Blokesch, 2012a) and belongs to the West‐African South American (WASA) lineage of the currently ongoing seventh cholera pandemic (Domman et al., 2017; Matthey et al., 2018). As strain‐specific differences are frequently reported, this study was designed to first confirm the generality of TfoX‐ and TfoY‐dependent T6SS activation in pandemic strains of V. cholerae. To do this, we tested five isolates from three different countries (Bangladesh, Peru and Bahrain) for T6SS‐dependent killing of Escherichia coli. Notably, we used a derivative of the first sequenced isolates of V. cholerae, pandemic strain N16961 (Heidelberg et al., 2000), in which the frameshift mutation in the quorum‐sensing regulator gene hapR was repaired (Kühn et al., 2014). As demonstrated in Supporting Information Fig. S1, all strains behaved similarly to A1552 upon TfoX or TfoY production. We, therefore, suggest that the dual control over T6SS by these two regulators is conserved among pandemic V. cholerae.
Functionality of TfoX and TfoY homologues from other Vibrio species in V. cholerae
Pollack‐Berti et al. showed that TfoX and TfoY existed in all fully sequenced Vibrionaceae (Pollack‐Berti et al., 2010), so we wondered whether these homologues would act in a comparable manner in non‐cholera vibrios despite the fact that these Vibrio species are often adapted to different environmental niches (Le Roux and Blokesch, 2018) and their homologous proteins might, therefore, serve different, adaptation‐specific functions. We, therefore, compared the protein sequences of TfoX and TfoY from four different Vibrio species, V. cholerae, V. parahaemolyticus, V. alginolyticus and V. fischeri, with the additional species chosen because previous studies had already described the genetic organization of their T6SS (Salomon et al., 2013; 2014; 2015; Speare et al., 2018). Moreover, these additional species have isolates that contain two T6SSs. We, therefore, considered the possibility that TfoX and TfoY, which are produced under vastly different conditions in V. cholerae (Metzger and Blokesch, 2016), might be specialized for the induction of a single T6SS in those organisms that harbour several systems. By aligning the protein sequences, we observed a high level of protein conservation for TfoX and TfoY, with a sequence identity of 68%/66%, 67%/66% and 59%/67% when the homologous proteins of V. parahaemolyticus (TfoXVp/TfoYVp), V. alginolyticus (TfoXVa/TfoYVa) and V. fischeri (TfoXVf/TfoYVf) were compared to the respective homologues of V. cholerae (Fig. 1A). Given these identity values, we concluded that the TfoX proteins of V. parahaemolyticus and V. alginolyticus were more closely related to the homologous protein of V. cholerae, whereas TfoX from V. fischeri diverged the most, which is in accordance with the phylogenetic distances between the species (Fig. 1A). Notably, TfoY remained equally conserved despite the phylogenetic divergence among the species.
Figure 1.
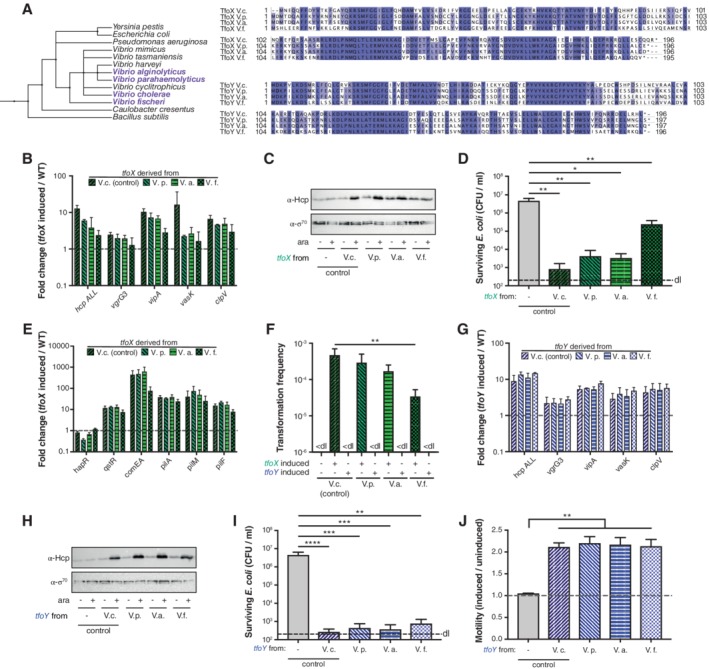
TfoX and TfoY proteins from diverse Vibrio species are functional in V. cholerae.
A. Phylogenetic tree and protein sequence conservation of TfoX and TfoY across Vibrio species. The phylogenetic tree was generated on phlyot.biobyte.de, a phylogenetic tree generator, based on NCBI taxonomy. The protein sequences of TfoX and TfoY were aligned using ClustalOmega, and subsequently modified with Jalview. Identical residues are highlighted in shades of blue (above threshold of 65%).
B–J. Chromosomally‐located tfoX and tfoY from cholera (V. cholerae) and non‐cholera Vibrio species (V. parahaemolyticus, V. alginolyticus and V. fischeri) were expressed under the control of the arabinose‐inducible PBAD promoter in V. cholerae. The colour code (TfoX‐induced, green; TfoY‐induced, blue) is used throughout the graphs. The WT strain lacking an inducible copy of tfoX or tfoY served as the negative control. The fold change (tfoX‐ or tfoY‐induced over parental WT strain) of relative gene expression of (B, G) T6SS genes or (E) representative competence genes is shown.
C, H. Detection of Hcp protein produced by TfoX‐ and TfoY‐expressing cells. Cells were grown in the absence or the presence of the inducer (arabinose) as indicated below the image. Detection of σ70 served as a loading control.
D, I. Interspecies killing assay with E. coli as prey. V. cholerae harbouring different inducible versions of (D) tfoX or (I) tfoY were co‐cultured with the prey on LB agar plates supplemented with arabinose. The survival of the prey was determined on selective LB agar plates and is depicted as CFU per ml.
F. Natural transformation using genomic DNA is maintained in a V. cholerae strain expressing tfoX from non‐cholera vibrios but is nonfunctional upon tfoY expression. The indicated strains were grown under inducible conditions, and the genomic DNA of A1552‐lacZ‐Kan served as the transforming material. Transformation frequencies reflect the number of transformants divided by the total number of CFUs. < dl, below detection limit.
J. Motility was scored on soft agar with and without arabinose as an inducer. The motility phenotype was quantified as the ratio between the induced and uninduced conditions, as shown on the Y‐axis. Abbreviations: V.c., Vibrio cholerae A1552, V.p., Vibrio parahaemolyticus RIMD2210633; V.a., Vibrio alginolyticus 12G01; V.f., Vibrio fischeri ES114. Bar plots represent the average of at least three independent biological replicates (± SD). Statistical significance is indicated (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
Given the high level of protein conservation, we next tested whether the protein homologues from the non‐cholera vibrios could function in V. cholerae. For this purpose, we cloned the respective genes preceded by an arabinose‐inducible promoter (PBAD) onto a site‐specific transposon (mini‐Tn7) together with the gene that encodes the arabinose‐responsive regulatory protein AraC (Supporting Information Table S1). Next, we grew the genetically engineered strains and the parental WT strain in the presence of arabinose (as inducer), isolated their RNA and then quantified the transcript levels of representative T6SS‐encoding genes. As shown in Fig. 1B, heterologous production of the diverse TfoX proteins resulted in increased T6SS transcript levels compared to the uninduced conditions, though the level of induction was slightly lower than the production of V. cholerae's own TfoX, especially for the most diverging protein variant from V. fischeri (TfoXVf). We confirmed these data at the protein level through the detection of the inner tube protein of the T6SS, Hcp (Fig. 1C). Consistent with the production of Hcp and the increase of the T6SS transcript level, these strains were able to kill E. coli in an interbacterial competition assay, with the least predatory behaviour observed for the most divergent TfoXVf ‐carrying variant (Fig. 1D).
TfoX of V. cholerae was first identified as the master regulator of natural competence for transformation (Meibom et al., 2005; Metzger and Blokesch, 2016). The TfoX‐induced competence regulon includes those genes that encode the structural components of the DNA‐uptake machinery, which primarily consists of a central pilus structure (encoded by pil genes) (Seitz and Blokesch, 2013) and the DNA binding protein ComEA that is essential for reeling DNA across the outer membrane and into the periplasmic space (Seitz and Blokesch, 2014; Seitz et al., 2014; Matthey and Blokesch, 2016). We, therefore, tested whether the TfoX variants from the other Vibrio species would likewise foster competence‐gene expression, which was indeed the case (Fig. 1E). Compared to the other variants, the more diverse TfoXVf did not induce the same high transcript levels of comEA, which, accordingly, also led to lower transformation frequencies (Fig. 1F). Notably, comEA expression in V. cholerae is regulated by an intermediate transcription factor, QstR, which is dependent on the production of both TfoX and HapR (Lo Scrudato and Blokesch, 2013; Jaskólska et al., 2018). HapR is the master regulator of the QS system, meaning that comEA expression is dependent on a functional QS system and on high cell density. As V. fischeri – but not the other tested Vibrio species – lacks a QstR homologue, we suggest that TfoXVf induces comEA at lower levels, as it is adapted to the more diverse QS circuit of V. fischeri and the absence of QstR (Milton, 2006; Verma and Miyashiro, 2013).
To assess the functionality of the TfoY homologues in V. cholerae we first tested the ability of the TfoY variants to foster natural transformation, given that these proteins are frequently annotated as ‘DNA transformation protein TfoX’ or ‘TfoX‐family DNA transformation protein’ due to their similarity to TfoX. However, no transformants were detected upon the production of any of these proteins, indicating their inability to induce natural competence in V. cholerae (Fig. 1F). Conversely and consistent with the high sequence similarity of the proteins from the four organisms, all TfoY variants functioned at the same level as the V. cholerae TfoY to increase T6SS transcript levels (Fig. 1G), to induce Hcp protein production (Fig. 1H), and, consequently, to foster interbacterial killing of E. coli (Fig. 1I). In addition, the four TfoY variants were able to induce motility as a T6SS‐independent phenotype, as visualized on swarming agar plates and quantified in Fig. 1J. Taken together, these data indicate that TfoX and TfoY are conserved proteins among these four Vibrio species and that the TfoY variants can fully replace the V. cholerae innate protein, while the TfoX variants are functionally replaceable at high levels for TfoXVp/TfoXVa and intermediate levels for TfoXVf.
TfoX but not TfoY induces V. fischeri's primary T6SS
While the complementation assay in V. cholerae described above provided a first insight into the conservation of TfoX and TfoY from diverse vibrios by indicating their similarity to those of V. cholerae, the experiments did not tell us whether these proteins would be involved in similar phenotypes as in V. cholerae in their native hosts. We, therefore, genetically engineered the different Vibrio species to place inducible copies of their own tfoX and tfoY genes onto a mini‐Tn7 transposon that was then integrated into their own genomes (Supporting Information Table S1). Next, we analysed the different phenotypes under uninduced and induced conditions. For V. fischeri strain ES114 (Supporting Information Table S1), we showed that TfoX production led to a high induction of T6SS transcripts, while TfoY only partially induced a subset of T6SS genes (Fig. 2A). Consistent with these expression data, we witnessed the T6SS‐mediated killing of E. coli only upon TfoX induction, while TfoY induction did not change the number of surviving co‐cultured prey bacteria (Fig. 2B).
Figure 2.
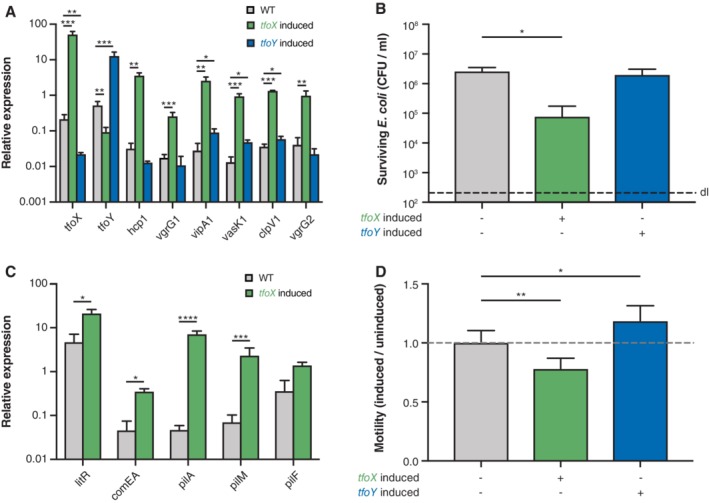
. TfoX but not TfoY induce V. fischeri's T6SS.
The effect of TfoX (green) and TfoY (blue) production in V. fischeri ES114 was scored by qRT‐PCR, and the relative expression values comparing WT versus TfoX‐ or TfoY‐induced conditions are indicated for representative (A) T6SS or (C) competence genes.
B. TfoX induction in V. fischeri leads to interbacterial killing of E. coli prey cells. V. fischeri strains were co‐cultured with E. coli for 4 h at 28°C on plain LBS agar plates supplemented with arabinose to induce the arabinose‐inducible copy of tfoX or tfoY (as indicated below the graph). The values for the recovered prey are indicated on the Y‐axis.
D. TfoY fosters motility in V. fischeri. Quantification of the motility phenotype of TfoX‐ or TfoY‐induced bacteria normalized by the uninduced conditions. The WT strain without any inducible gene served as a control.
A–D. Bar plots represent the average of at least three independent biological replicates (± SD). Statistical significance is indicated (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
Pollak‐Berti et al. showed that TfoX was required for natural competence for transformation in V. fischeri (Pollack‐Berti et al., 2010), as is the case for V. cholerae (Meibom et al., 2005). We, therefore, measured relative transcript levels of representative competence genes in this organism and confirmed their induction upon TfoX production (Fig. 2C). Surprisingly, while we only observed a mild induction of comEA relative to the levels of induction seen in V. cholerae (Fig. 1), we witnessed an almost 10‐fold upregulation of litR, which encodes the main QS regulator LitR (homologue of V. cholerae's HapR). This phenotype differs from what is known about competence regulation in V. cholerae in which case hapR transcript levels are left unchanged upon TfoX induction (Meibom et al., 2005; Metzger and Blokesch, 2016). We, therefore, speculate that an increase in LitR levels can compensate for the lack of the intermediate regulator QstR in V. fischeri. Indeed, the QstR protein of V. cholerae is required for competence‐mediated DNA uptake in two ways: (i) through direct and indirect induction of certain competence genes, such as comEA, as described above; and (ii) due to its ability to downregulate the dns gene (Lo Scrudato and Blokesch, 2013; Jaskólska et al., 2018) encoding for the extracellular nuclease Dns, which degrades transforming material outside the cell and within the periplasmic space (Blokesch and Schoolnik, 2008; Seitz and Blokesch, 2014). Notably, the HapR protein of V. cholerae also silences dns expression, but only partially in the absence of QstR (Lo Scrudato and Blokesch, 2013). It is, therefore, likely that the increased production of LitR upon TfoX production in V. fischeri compensates for the absence of QstR to ensure sufficient repression of dns, which, ultimately, would allow DNA uptake to occur. It is not clear, however, why the comEA transcripts were not induced to higher levels, though we recently demonstrated for V. cholerae that the comEA gene requires an additional but so far unidentified regulatory input apart from TfoX and QstR (Jaskólska et al., 2018). Indeed, artificial QstR production was sufficient to induce the T6SS genes in this organism at comparable levels as the upstream regulatory protein TfoX, even in the absence of HapR, while the comEA transcript levels were only partially increased compared to a noninduced WT strain. We, therefore, hypothesize that this input signal might be conserved in V. fischeri, though missing under the tested conditions. This would lead to a high overall competence‐gene induction upon TfoX production but lower comEA transcripts as compared to V. cholerae (Fig. 1E).
Lastly, we determined that the motility of V. fischeri changed upon TfoY production (Fig. 2D), even though the difference between the TfoY‐induced and TfoY‐uninduced state was not as pronounced as for V. cholerae (Fig. 1J). We argue that this might be caused by a basal production level of TfoY in this organism that does not rely on the artificial induction. A higher basal TfoY level would result in a higher basal motility, which would, therefore, appear as a less pronounced induction upon additional TfoY production. There was also a decrease in motility observable upon TfoX production (Fig. 1D), which is consistent with the idea of a basal TfoY level, as the expression data showed that TfoX significantly lowered the tfoY transcript levels (Fig. 2A). To see whether this is a conserved regulatory phenomenon, we tested whether TfoX production also lowered tfoY levels in V. cholerae, which was indeed the case with an average repression of 4.85 (± 0.5)‐fold (based on three independent biological experiments; p = 0.0001). For V. fischeri TfoY production lowered tfoX transcript levels (Fig. 2A), which was likewise the case for V. cholerae though to a lower extend (1.45 ± 0.1‐fold; p = 0.025), suggesting that both proteins work independently of each other and are mutually exclusive. This finding contradicts a previous study on V. fischeri in which the authors concluded that TfoY was required for efficient TfoX‐mediated transformation (Pollack‐Berti et al., 2010). Notably, both studies used different conditions (e.g. inducible TfoX/TfoY expression in this study versus a transposon‐inserted mutant of tfoY (Pollack‐Berti et al., 2010)) and further studies are, therefore, required to conclusively show if TfoY is indeed involved in natural transformation in V. fischeri or not, as we suggest based on the herein‐described data.
TfoX and TfoY have a different impact on T6SS induction in V. alginolyticus
While V. cholerae strains and the examined V. fischeri strain ES114 contain only a single T6SS, several other Vibrio species contain more than one. Hence, we asked whether, and if so how, TfoX and TfoY affect the production of different T6SSs within the same organism. To address this question, we first tested the contribution of these regulators to the two T6SSs (T6SS1 and T6SS2) of V. alginolyticus in an E. coli‐killing assay using wildtype (WT) V. alginolyticus or its hcp1 (referred to as ΔT6SS1 throughout the text), hcp2 (referred to as ΔT6SS2 throughout the text) or double mutant as a predator. Consistent with a previous report by Salomon et al. (2015), WT V. alginolyticus did not show interbacterial predation at 37°C on standard LB medium (e.g. without additional salt; Fig. 3A). Upon production of TfoX or TfoY, however, predation by the WT was significantly increased. This effect was less pronounced but reproducible in the strain that only carried the complete T6SS2 gene set (ΔT6SS1), while a strain that only carried a complete T6SS1 gene set (ΔT6SS2) was not able to kill the E. coli cells, indicating that T6SS1 is nonfunctional at this temperature. In contrast, both systems were inducible at 30°C, with T6SS1 being specifically induced by TfoY and not TfoX, while T6SS2 was responsive to both regulatory proteins (Fig. 3B). Basic killing activity was also observed in the WT and the mutant lacking T6SS1 but not in a mutant lacking T6SS2, suggesting that T6SS2 is induced under the tested conditions (in a TfoX‐ and TfoY‐independent manner; Fig. 3C), but can be further boosted upon TfoX and TfoY production (Fig. 3B). While this finding seems at first glance to contradict a previous study that reported basal activity for T6SS1 (Salomon et al., 2015), it should be noted that the experimental setup differed in that we used an assay in which the Vibrio are tested at high cell density, as was previously developed for V. cholerae (Borgeaud et al., 2015), while Salomon et al. first diluted the cells to low densities. The rationale behind our experimental setup was the knowledge that the TfoX‐mediated pathway requires co‐induction by HapR in V. cholerae (homologues are LuxR in V. alginolyticus and OpaR in V. parahaemolyticus) (Lo Scrudato and Blokesch, 2012; Metzger and Blokesch, 2016; Jaskólska et al., 2018), which we assumed to be a conserved feature in other Vibrio species (see data below).
Figure 3.
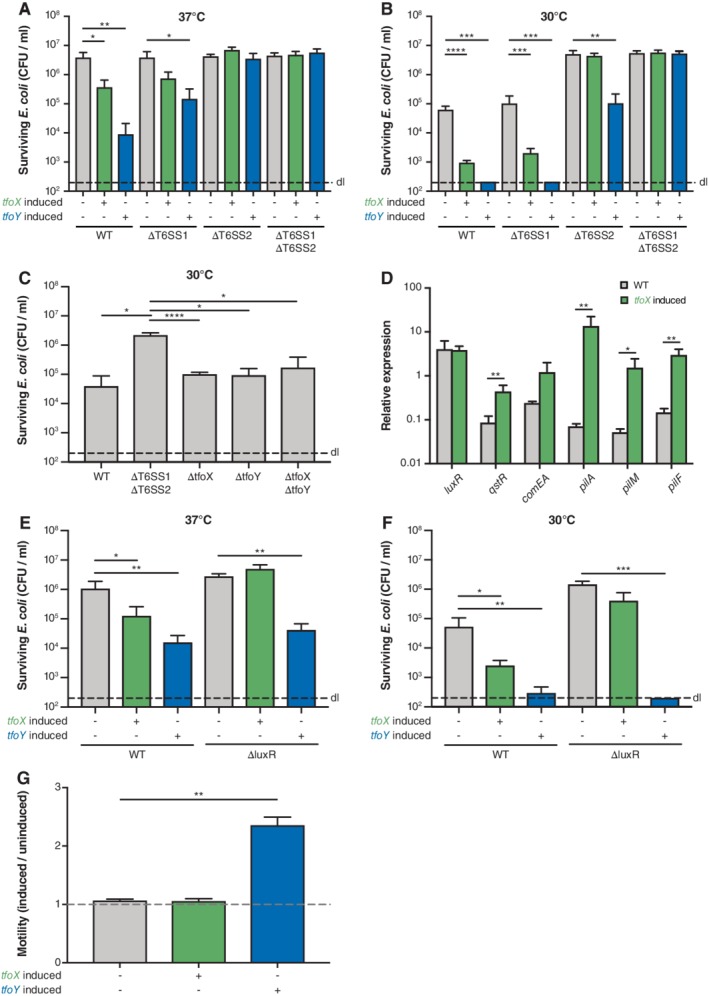
TfoY is a universal T6SS inducer in V. alginolyticus, while TfoX is specific for T6SS2.
A–C, E, F. Interspecies killing assay of V. alginolyticus with E. coli as prey. V. alginolyticus strains were co‐cultured with E. coli prey for 4 h at (A, E) 37°C or (B, C, F) 30°C on plain LB agar plates (C) or plates supplemented with arabinose (A, B, E, F; to induce tfoX or tfoY, as indicated below the graph). The recovery of the prey is indicated as CFU/ml on the Y‐axis.
D. TfoX induces competence genes in V. alginolyticus. The relative expression of representative competence genes was scored in WT and tfoX‐induced strains as indicated on the Y‐axis.
G. Motility ratio between uninduced and induced V. alginolyticus strains is indicated. Details as in Fig. 2D. Abbreviations: WT, V. alginolyticus strain 12G01; ΔT6SS1, 12G01Δhcp1; ΔT6SS2, 12G01Δhcp2; ΔT6SS1ΔT6SS2, 12G01Δhcp1Δhcp2; ΔtfoX, 12G01ΔtfoX; ΔtfoY, 12G01ΔtfoY; ΔtfoXΔtfoY, 12G01ΔtfoXΔtfoY; ΔluxR, 12G01ΔluxR. Bar plots represent the average of at least three independent biological replicates (± SD). Statistical significance is indicated (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
In addition to T6SS activation, we also tested the level of competence genes transcripts and bacterial motility upon TfoX and TfoY induction. As shown in Fig. 3D, a significant TfoX‐mediated induction of representative competence genes occurred, indicating the protein's conserved action. Notably, the comEA transcript induction levels again turned out to be unexpectedly low compared to the induction levels in V. cholerae (Fig. 1E). ComEA plays a primary role in ratcheting the DNA into the periplasmic space in competent bacteria (Seitz et al., 2014), and we, therefore, expect that these low transcript levels would lead to insufficient amounts of the ComEA proteins and defective DNA uptake. As comEA is co‐regulated by the TfoX‐ and QS pathways in V. cholerae (Suckow et al., 2011; Lo Scrudato and Blokesch, 2012), comparable to the T6SS genes (Borgeaud et al., 2015; Metzger et al., 2016), we wondered whether QS was impaired in the tested V. alginolyticus strain. As demonstrated in Fig. 3E and F, TfoX‐mediated T6SS induction worked efficiently in the WT but was impaired in a luxR‐minus strain (the hapR homologue in this organism; Supporting Information Table S1). These data indicate that QS is functional in the WT strain and support the notion that TfoX and LuxR co‐regulate the T6SS2, consistent with earlier data that showed a requirement of LuxR for basal T6SS2 activity in V. alginolyticus (Yang et al., 2018). TfoY‐mediated T6SS activation, however, occurred independently of LuxR (Fig. 3E and F) consistent with what is known for V. cholerae (Metzger et al., 2016). Based on these data, we conclude that the additional but so far unknown regulatory input required for comEA expression aforementioned (Jaskólska et al., 2018) might also be a prerequisite in V. alginolyticus and that this pathway and not the QS network overall might be nonfunctional in either the strain used in this study or under the tested conditions or both. The motility‐inducing phenotype of TfoY, on the other hand, was very pronounced in V. alginolyticus (Fig. 3G), strengthening the idea that TfoY protein levels and a high motility are linked throughout the genus Vibrio, even in organisms that contain lateral flagella in addition to the polar flagellum (McCarter, 2001).
TfoX and TfoY are both dedicated to one specific T6SS in V. parahaemolyticus
Lastly, we tested the contribution of TfoX and TfoY to T6SS regulation in V. parahaemolyticus. As with V. alginolyticus, this bacterium also contains two T6SSs, T6SS1 and T6SS2, which were previously shown to be differentially regulated from each other with respect to temperature, salinity, QS inputs and surface sensing (Salomon et al., 2013). Indeed, T6SS1 of V. parahaemolyticus was found to be most active under marine‐like conditions and at low cell densities while system 2 was more adapted to lower salt conditions (e.g. LB medium) and higher cell densities (Salomon et al., 2013). Notably, these data on T6SS2 regulation and a previous study that suggested T6SS2 was involved in adhesion to host cells were solely based on the Hcp2 protein levels (inside and outside the cells), while interbacterial killing activity was not directly observed for T6SS2 (Yu et al., 2012; Salomon et al., 2013). This QS‐dependent T6SS2 regulation of Hcp production confirmed previous results in V. cholerae (Ishikawa et al., 2009) in which the authors demonstrated that HapR was required for hcp expression. However, despite the fact that pandemic V. cholerae produce detectable levels of Hcp at high cell densities in vitro, the bacteria are unable to kill prey under such conditions (Pukatzki et al., 2006), which can be overcome by TfoX or TfoY production (Metzger et al., 2016). Indeed, we previously demonstrated that upon induction of these two regulators, the Hcp levels increased further, and T6SS‐mediated prey killing was observed. Notably, TfoY‐mediated T6SS activity in V. cholerae occurred independently of HapR, indicating that TfoY acts independently from the QS pathway, in contrast with TfoX‐mediated T6SS activation (Borgeaud et al., 2015; Metzger et al., 2016).
In V. parahaemolyticus, both TfoX and TfoY production led to significant E. coli killing (Fig. 4A). When testing V. parahaemolyticus strains that contained only a complete T6SS2 gene set (ΔT6SS1) or only all T6SS1 genes (ΔT6SS2), it was clear that TfoX specifically induced T6SS2, while TfoY solely led to T6SS1‐mediated prey killing (Fig. 4A). These data also suggested that the T6SS2 was slightly active in both the WT and the strain lacking T6SS1, even without artificial TfoX/TfoY production and that neither TfoX nor TfoY caused this basal T6SS2 activity (Fig. 4B). Interestingly, a recent study showed T6SS2‐dependent killing of a V. parahaemolyticus strain that lacked a newly identified effector immunity pair (RhsP and RhsPi) by its parental WT strain, even in the absence of TfoX or TfoY induction (Jiang et al., 2018). These data, therefore, suggest that the basal killing activity that we observed against E. coli as a prey might be more enhanced against nonimmune siblings or, alternatively, that the experimental conditions were different to the current work and that under such conditions the T6SS2 is highly active. Lastly, we cannot exclude the possibility that strain differences or the domestication of certain strains has caused the difference in basal T6SS activity. Notably, the focus of the current study was on the contribution of TfoX or TfoY to T6SS activity, which we unambiguously demonstrate to enhance T6SS activity in V. parahaemolyticus.
Figure 4.
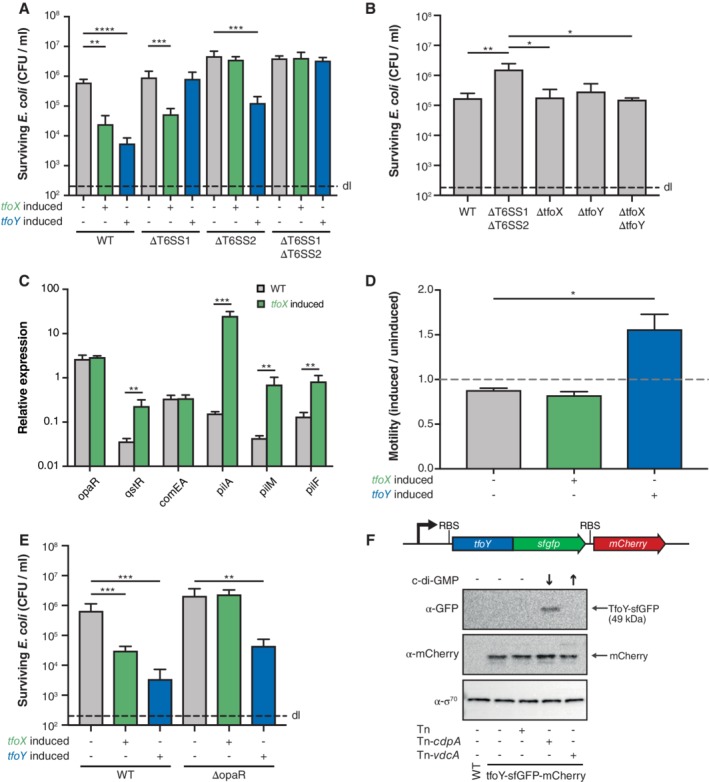
TfoX and TfoY each induce a dedicated T6SS in V. parahaemolyticus.
A, E. Interspecies killing of E. coli by V. parahaemolyticus. V. parahaemolyticus strains (WT, T6SS1‐, T6SS2‐, T6SS1/T6SS2‐minus or opaR‐minus) were co‐cultured with E. coli prey for 4 h at 30°C on LB agar plates supplemented with arabinose to induce tfoX or tfoY (as indicated below the graph). The recovery of the prey is indicated on the Y‐axis.
B. Basal T6SS activity is independent of TfoX and TfoY. WT V. parahaemolyticus or its tfoX‐, tfoY‐ or tfoX/tfoY‐minus variants were tested for their ability to reduce the number of E. coli prey cells. The T6SS double mutant (ΔT6SS1ΔT6SS2) served as a control.
C. Relative expression of representative competence genes in WT or tfoX‐induced bacteria.
D. Quantification of TfoX‐ or TfoY‐induced motility phenotypes. Details as in Fig. 2D.
F. TfoY is produced under low c‐di‐GMP conditions in V. parahaemolyticus. Detection of TfoY‐sfGFP and mCherry by western blotting in cdpA‐ or vdcA‐inducible reporter strains that contain increased or decreased intracellular c‐di‐GMP levels, as indicated by the arrows. The transposon‐less or empty transposon‐carrying reporter strain as well as the parental WT strain served as controls. Detection of σ70 served as a loading control. RBS, ribosome‐binding site.
Abbreviations: WT, V. parahaemolyticus POR1 (RIMD2210633 derivative); ΔT6SS1, POR1Δhcp1; ΔT6SS2, POR1Δhcp2; ΔT6SS1ΔT6SS2, POR1Δhcp1Δhcp2. Bar plots represent the average of at least three independent biological replicates (± SD). Statistical significance is indicated (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
Examining other TfoX‐ and TfoY‐mediated phenotypes confirmed the increase of competence gene transcripts and enhanced motility respectively (Fig. 4C and D). However, comEA transcript levels were again unexpectedly low even though the induction levels of qstR were comparable with what we observed for V. cholerae (Fig. 1E). While these data were similar to the above‐described observations for V. alginolyticus (Fig. 3D), which suggested a comEA‐specific expression defect, we nonetheless tested whether the V. parahaemolyticus strain used in this study was QS proficient or QS deficient. Indeed, previous studies in different Vibrio species described gain‐of‐function mutations in luxO, encoding a repressor of HapR/LitR/OpaR synthesis (Gode‐Potratz and McCarter, 2011; Kimbrough and Stabb, 2015; Stutzmann and Blokesch, 2016) and clinical V. parahaemolyticus isolates with such luxO mutations were previously described to be locked in a pathogenic state (Kernell Burke et al., 2015). To exclude that this has happened in the strain we were working with, we first sequenced luxO and opaR and confirmed that both genes were mutation free. Additionally, we deleted opaR from the WT strain and compared this strain to its TfoX‐ or TfoY‐inducible derivatives (Supporting Information Table S1) in an E. coli‐killing assay. As shown in Fig. 4E, the TfoY‐induced prey killing was maintained upon deletion of opaR, while the TfoX‐mediated T6SS activity was abrogated. Since OpaR is produced at high cell density, its deletion blocks QS‐dependent signalling. We, therefore, conclude that the WT strain has a functional OpaR and that TfoX‐mediated T6SS induction in V. parahaemolyticus requires co‐regulation by OpaR, similar to what we have shown for V. cholerae (Borgeaud et al., 2015; Metzger et al., 2016) and for V. alginolyticus (Fig. 3E and F). These data also support the idea that the low expression levels seem specific for comEA and not based on a general mutation within the QS cascade. The low level of the comEA transcripts also explains why we were unable to naturally transform V. parahaemolyticus even after TfoX induction. Interestingly, Chimalapti et al. recently reported rare transformants for competence‐induced V. parahaemolyticus with frequencies between 1 × 10−8 and 2.7 × 10−6 (Chimalapati et al., 2018), which corresponds to 0.01% to 1% of what is commonly observed for V. cholerae (see Fig. 1 and (Lo Scrudato and Blokesch, 2012) despite the ∼1000‐fold molar excess of the selective resistance marker that was used as transforming DNA in this study. It should be noted that these experiments were based on the heterologous expression of the V. cholerae tfoX homologue from a multicopy plasmid, which makes a direct comparison with the current study on the bacterium's indigenous TfoX protein impossible.
Our data suggest that TfoX induces the T6SS2 in V. parahaemolyticus. As previous studies showed that the chitin sensors ChiS, TfoS and the downstream‐regulated small RNA TfoR, which is required for tfoX mRNA translation, are highly conserved in diverse Vibrio species (Li and Roseman, 2004; Yamamoto et al., 2011; 2014), we suggest that TfoX is exclusively produced upon growth on chitinous surfaces, as first demonstrated for V. cholerae (Meibom et al., 2004; 2005). The production of TfoY is less understood. In a previous study on V. cholerae, we showed that lowered c‐di‐GMP levels led to the production of TfoY under standard laboratory conditions (e.g. in LB medium) and we and others suggested that this c‐di‐GMP‐dependent control occurred at the translational level (Inuzuka et al., 2016; Metzger et al., 2016). Interestingly, Gode‐Potratz et al. demonstrated that growth on surfaces lowered the levels of the secondary messenger c‐di‐GMP in V. parahaemolyticus (Gode‐Potratz et al., 2011). Consistently, Salomon et al. established that surface sensing correlated with Hcp1 production in this organism (Salomon et al., 2013). Based on these data, we speculated that surface sensing, TfoY production and T6SS1 induction might be linked. To address this hypothesis and especially the link between low c‐di‐GMP levels and TfoY production, we genetically engineered V. parahaemolyticus to carry a translational fusion between its indigenous tfoY gene and the gene that encodes super‐folder GFP (sfGFP). In addition, we included a transcriptional mCherry‐encoding reporter gene behind this construct, which, ultimately, allowed us to monitor the transcriptional and translational control of tfoY. Next, we incorporated either an empty mini‐Tn7 transposon (Tn) or transposons that carried arabinose‐inducible genes coding for a c‐di‐GMP‐producing diguanylate cyclase (Tn‐vdcA) or for a phosphodiesterase (Tn‐cdpA) into this strain (Supporting Information Table S1). These strains as well as the transposon‐deficient parental strain and the WT without any fluorescent protein‐encoding genes were then grown in the presence of the inducer arabinose followed by a western blot analysis to visualize the TfoY‐sfGFP fusion protein or the transcriptional reporter protein mCherry. Using this approach, we observed that TfoY was produced solely at low c‐di‐GMP levels (Fig. 4F). Importantly, based on the comparable levels of mCherry in all reporter strains, we concluded that the tfoY transcript levels did not significantly change in response to increased or decreased c‐di‐GMP levels (Fig. 4F). This finding contradicts a recent study in V. cholerae that concluded that TfoY was induced at both low and high intracellular concentrations of c‐di‐GMP and that this regulation occurred at the translational and transcriptional levels, the latter due to activation by the c‐di‐GMP‐dependent transcription factor VpsR (Pursley et al., 2018). We, therefore, constructed a similar dual translational‐transcriptional reporter strain in V. cholerae (Supporting Information Table S1) and tested this strain under normal, increased or decreased c‐di‐GMP levels. Using this approach, we were able to detect an increase in the TfoY protein under low c‐di‐GMP levels compared to normal conditions, while an increase in intracellular c‐di‐GMP did not result in higher TfoY protein levels (Supporting Information Fig. S2), consistent with our previous findings (Metzger et al., 2016). Importantly, we also did not observe a change in the transcriptional reporter under high c‐di‐GMP conditions (Supporting Information Fig. S2), despite normal VpsR function in the pandemic V. cholerae strain (A1552) used in this study (Yildiz et al., 2001), which confirmed the data presented above for V. parahaemolyticus. An explanation for this discrepancy might be that we engineered the construct at the native locus of tfoY within the V. parahaemolyticus or V. cholerae genomes, while Pursley et al. were unsuccessful in detecting tagged TfoY variants using a similar approach. They, therefore, decided to construct a TfoY‐GFP translational‐fusion‐encoding reporter gene on a plasmid, which could change the expression pattern compared to the gene's native locus. In addition, these authors performed their experiments in a Vibrio polysaccharide mutant as parental strain (ΔvpsL), which is deficient in biofilm formation (Pursley et al., 2018), while our strain maintained a normal ability for this process.
Collectively, we conclude that the lowered c‐di‐GMP levels observed in surface‐sensing V. parahaemolyticus (Gode‐Potratz et al., 2011) might trigger TfoY induction, which induces the T6SS1 and subsequently initiates motility, though the exact mechanism for the latter remains to be discovered. The data provided here for V. parahaemolyticus, therefore, support our previous suggestion that TfoY triggers a defensive escape reaction that might allow Vibrio species to defend themselves against bacterial competitors and eukaryotic predators. Interestingly, a recent study that profiled the gene expression of V. parahaemolyticus reported increased tfoY expression (VP1028; falsely annotated as tfoX and, therefore, discussed as competence regulator in this study) within infected infant rabbits when compared to in vitro conditions (Livny et al., 2014), suggesting that TfoY might also play a role in vivo and, therefore, in pathogenesis.
Conclusion
In this study, we investigated the ecological implications upon TfoX and TfoY production in diverse Vibrio species. We showed a conserved pattern of competence and motility induction by TfoX and TfoY, respectively, and demonstrated that the link between the TfoX and TfoY regulatory proteins and T6SSs is highly conserved among cholera and non‐cholera Vibrio species (Fig. 5), highlighting the general importance of these regulators. Notably, TfoX induced at least one T6SS in every tested species concomitantly with an interbacterial killing phenotype. Hence, we suggest that chitin‐mediated TfoX production (Meibom et al., 2005; Metzger and Blokesch, 2016) and the resulting coupling between T6SS‐facilitated neighbour killing and competence‐mediated DNA uptake (Borgeaud et al., 2015) might be an ancient phenotype maintained in most Vibrio species. Our study also suggests that TfoX and TfoY are mutually exclusive (Fig. 2A and data presented above). Indeed, TfoX‐mediated tfoY repression seems like a prerequisite for proper colonization of chitinous surfaces, as TfoY‐induced motility would otherwise counteract this process. Thus, even upon contact with the surface and the reported decrease in c‐di‐GMP levels (Gode‐Potratz et al., 2011), tfoY translation would not occur on chitin due to the absence/low levels of the template mRNA. In contrast, when vibrios sense non‐chitinous surfaces, resulting in the absence of the chemoattracting chitin‐degradation product chitobiose (Keyhani and Roseman, 1999), TfoY would be produced. Interestingly, the effects of TfoY turned out to be more versatile compared to TfoX (Fig. 5). While the c‐di‐GMP‐dependent inhibition of TfoY translation seemed conserved, at least in the two tested Vibrio species (Fig. 4E and Supporting Information Fig. S2), the regulons seem to have diverged in the different organisms. Indeed, TfoY induces both T6SSs in V. alginolyticus and one T6SS in V. cholerae and V. parahaemolyticus (out of two T6SSs present in the latter bacterium) but this regulator does not impact the activity of the T6SS of V. fischeri under the tested conditions. However, several isolates of V. fischeri, other than the squid isolate ES114, contain a secondary T6SS. This T6SS2 is located on a strain‐specific genomic island on the small chromosome and was recently shown to contribute to competitor elimination within the host's light organ (Speare et al., 2018). While we were unable to genetically engineer the T6SS2‐containing fish symbiont MJ11 strain for technical reasons, it is tempting to speculate that this secondary T6SS of V. fischeri might be controlled by the TfoY protein. In line with this idea is the fact that V. fischeri's secondary T6SS is broadly conserved among vibrios and is similar to the TfoY‐inducible T6SS1 of V. alginolyticus and V. parahaemolyticus (Speare et al., 2018), which we confirmed by BLAST analyses of V. fischeri's T6SS2 genes VFMJ11_A0804 to VFMJ11_A0809. Notably, Speare et al. proposed the possibility that this secondary T6SS2 was present in the common ancestor of V. fischeri and related Vibrio species but was eventually lost in niche‐adapted strains for which interbacterial killing was no longer advantageous. We extend this hypothesis and speculate that niche‐adapted vibrios might no longer require a TfoY‐mediated defensive response against either competing bacteria or grazing predators. Future studies are required to address these interesting evolutionary hypotheses.
Figure 5.
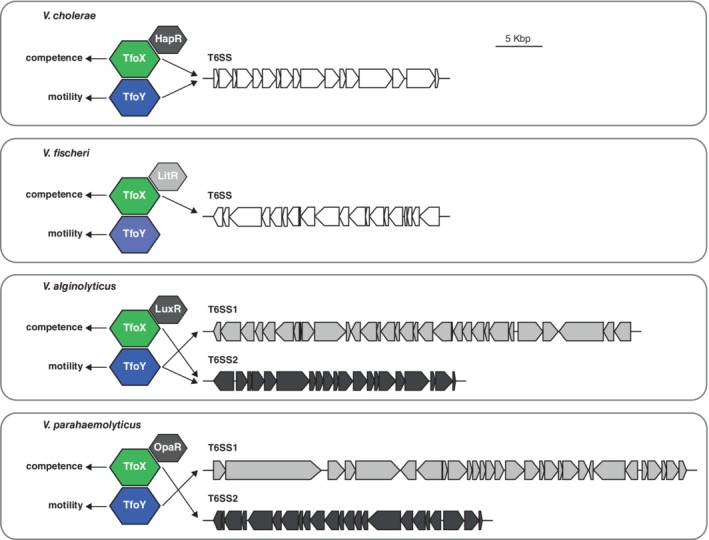
Summary scheme of TfoX‐ and TfoY‐induced phenotypes in V. cholerae and non‐cholera Vibrio species.
TfoX‐ and TfoY‐mediated regulation of natural competence, motility and type VI secretion is depicted. The T6SS clusters of V. cholerae (only the major cluster is shown), V. fischeri (only the conserved T6SS cluster is shown), V. alginolyticus and V. parahaemolyticus are indicated (scale corresponds to 5 kbp as indicated). Arrows indicate positive regulation by the respective regulator TfoX or TfoY. The scheme also shows the co‐regulating QS master regulators of TfoX‐mediated phenotypes (experimentally demonstrated for HapR, LuxR and OpaR and predicted for LitR).
Experimental procedures
Bacterial strains, plasmids and growth conditions
The V. cholerae, V. alginolyticus, V. parahaemolyticus and V. fischeri strains and plasmids used in this study are listed in Supporting Information Table S1. E. coli strains DH5α (Yanisch‐Perron et al., 1985), TOP10 (Invitrogen), SM10λpir (Simon et al., 1983), S17‐1λpir (Simon et al., 1983) and MFDpir (Ferrieres et al., 2010) were used for cloning purposes and/or served as donor in bacterial mating experiments.
Unless otherwise stated, the V. cholerae, V. alginolyticus, V. parahaemolyticus and E. coli strains were grown aerobically in Lysogeny broth (LB; 10 g/l of tryptone, 5 g/l of yeast extract, 10 g/l of sodium chloride; Carl Roth) or on LB agar plates at 30°C or 37°C. V. fischeri strains were cultured aerobically in LB salt (LBS) medium (10 g/l of tryptone, 5 g/l of yeast extract, 20 g/l of sodium chloride, 20 mM Tris–HCl [pH 7.5], 0.2% glycerol; adapted from (Dunlap, 1989; Graf et al., 1994)) or LBS agar plates at 28°C. LB/LBS motility plates had less agar (0.3%) than standard LB/LBS agar plates (1.5%). The following compounds were added if required at the given concentrations: arabinose (0.02% or 0.2%), diaminopimelic acid (DAP; 0.3 mM) or the antibiotics kanamycin (75 μg/ml), gentamicin (50 μg/ml), ampicillin (100 μg/ml) and chloramphenicol (2.5 μg/ml or 5 μg/ml). l‐arabinose‐supplemented medium was used for the expression of tfoX or tfoY under the control of the PBAD promoter. DAP was added as an essential growth supplement for E. coli strain MFDpir (Ferrieres et al., 2010). Medium without DAP was used to counter‐select MFDpir strains after tri‐parental mating with V. fischeri. For the E. coli counter‐selection after tri‐parental mating with V. cholerae, V. alginolyticus and V. parahaemolyticus, Thiosulfate citrate bile salts sucrose (TCBS) agar plates were used and prepared following the manufacturer's instructions (Sigma‐Aldrich/Fluka, Buchs, Switzerland). Marine broth 2216 (BD Difco™ 2216) was used to isolate single colonies of V. alginolyticus and V. parahaemolyticus after E. coli counter‐selection.
Genetic engineering of strains and plasmids
DNA manipulations were performed according to standard molecular biology‐based protocols (Sambrook et al., 1982). Enzymes were purchased from the listed companies and were used as recommended by the manufacturer: Pwo polymerase (Roche), Taq polymerase (Promega), restriction enzymes (New England Biolabs). Following initial screening by PCR (using bacterial cells as templates), genetically engineered strains and plasmids were verified by Sanger sequencing (Microsynth, Switzerland).
V. cholerae strains were genetically modified using both a gene‐disruption method based on the counter‐selectable plasmid pGP704‐Sac28 (Meibom et al., 2004) or the TransFLP gene disruption method previously described by our group (De Souza Silva and Blokesch, 2010; Blokesch, 2012b; Borgeaud and Blokesch, 2013). This transformation‐based genetic engineering technique was used to replace the tfoY gene by a translational and transcriptional tfoY‐mCherry::gfp fusion at the natural locus on the V. cholerae A1552 chromosome. Plasmid pGP704‐Sac‐Kan (see below), derived from pGP704‐Sac28, was used for genetic modifications of V. parahaemolyticus and V. alginolyticus.
To construct the counter‐selectable plasmid pGP704‐Sac‐Kan, the aph gene was amplified from a kanamycin‐resistance gene (aph)‐carrying transposon using primers with overhanging SspI and BsaI restriction sites. A restriction digestion was performed on both the PCR product and the vector pGP704‐Sac28, which were subsequently ligated, to replace the bla gene of pGP704‐Sac28 with the aph gene, resulting in plasmid pGP704‐Sac‐Kan (Supporting Information Table S1).
All pBAD‐derived plasmids harbouring tfoX‐strep or tfoY‐strep genes from different Vibrio species were constructed in the following way: The genes were amplified with Strep‐tagII‐encoding primers using gDNA of the respective parental Vibrio strain as a template. The restriction‐enzyme‐digested PCR product was subsequently cloned into plasmid pBAD/MycHisA (Supporting Information Table S1). The resulting plasmids served as templates for fragments containing araC, the arabinose‐inducible promoter PBAD and the respective tfoX or tfoY genes, which were subcloned into the mini‐Tn7‐containing delivery plasmids (Supporting Information Table S1). The plasmid pGP704‐Tn‐CmR is a derivative of pGP704‐mTn7‐minus‐SacI (Nielsen et al., 2006), where the aacC1 gene within the mini‐Tn7 was replaced by the cat gene. To this end, the PCR‐amplified cat gene (including its promoter) of pBR‐FRT‐Cat‐FRT2 (Metzger et al., 2016) was cloned into pGP704‐mTn7‐minus‐SacI, which was digested using restriction enzymes SbfI and EcoRV. For the insertion of this mini‐Tn7 transposon into the Vibrio spp. chromosomes, a tri‐parental mating strategy was employed (Bao et al., 1991). The donor plasmids are indicated in Supporting Information Table S1.
Natural transformation assay
Natural transformation assays in liquid were performed with minor modifications to the previous protocol (Lo Scrudato and Blokesch, 2012). This assay is chitin‐independent and uses strains carrying an arabinose‐inducible copy of tfoX‐strep or tfoY‐strep (both from V. cholerae, V. alginolyticus, V. parahaemolyticus or V. fischeri) on the chromosome. V. cholerae strains were pre‐grown overnight in LB medium and further grown in the presence of the arabinose inducer up to an optical density at 600 nm (OD600) of 1.0. At this point, 0.5 ml of the cultures were supplemented with 1 μg of genomic DNA. For all natural transformation assays performed in V. cholerae, the genomic DNA of strain A1552‐lacZ‐Kan (Marvig and Blokesch, 2010) served as transforming material. Cells were further incubated under shaking conditions at 30°C for 4 h. Serial dilutions were spotted on LB to count the total number of cells and on kanamycin‐containing LB agar plates to select the transformants. Transformation frequencies were calculated as the number of colony‐forming units (CFUs) of the transformants divided by the total number of CFUs. Averages of at least three biologically independent experiments are provided. For statistical analyses, the data were log‐transformed (Keene, 1995) and significant differences were determined by the two‐tailed Student's t‐test. When no transformants were recovered, the value was set to the detection limit to allow for statistical analysis.
Gene expression analysis by quantitative reverse transcription PCR
Quantitative reverse transcription PCR (qRT‐PCR)‐based transcript scoring in V. cholerae, V. alginolyticus, V. parahaemolyticus and V. fischeri was performed following a previously established protocol (Lo Scrudato and Blokesch, 2012). Strains with and without an arabinose‐inducible copy of specified genes (tfoX‐strep or tfoY‐strep) were grown for 6 h in 2.5 ml LB or LBS supplemented with 0.2% arabinose. Cultures (2 ml) were processed for RNA isolation and subsequent cDNA synthesis. Relative gene expression values were normalized against the gyrA transcript levels. Fold changes were determined using the relative expression values of the induced strains divided by the values of the parental WT strain (as specified for each inducible construct in the figures). Averages of at least three biologically independent experiments (± SD) are provided with statistical analyses (two‐tailed Student's t‐test) being done on the log‐transformed data (Keene, 1995).
Interbacterial killing assay using E. coli as prey
The E. coli killing assay was performed following a previously established protocol with minor adaptions (Borgeaud et al., 2015). The E. coli prey cells and the given predator cells were mixed at a ratio of 1:10 and spotted onto membrane filters on pre‐warmed LB and/or LBS agar plates (± 0.2% ara). After 4 h of incubation at 28°C (predator: V. fischeri) or 30°C and 37°C (as indicated for the predators V. cholerae, V. alginolyticus and V. parahaemolyticus), bacteria were resuspended and serial dilutions were spotted onto antibiotic‐containing LB agar plates to enumerate the CFUs (shown as CFU/ml). Arabinose‐uptake‐deficient E. coli TOP10 and its derivative TOP10‐TnKan served as prey. Significant differences were determined by a two‐tailed Student's t‐test on log‐transformed data of at least three biological replicates. If no prey cells were recovered, the value was set to the detection limit to allow calculation of the average of the three independent biological experiments and statistical analysis.
Motility assay
The motility of the Vibrio species was assessed by spotting 2 μl of the respective overnight culture onto freshly prepared LB and/or LBS motility agar plates (containing 0.3% agar) with or without 0.2% arabinose. Following the incubation at 30°C for 6 h for V. cholerae, V. parahaemolyticus and V. alginolyticus, or at room temperature for 7 h for V. fischeri, the diameters of the bacterial swarming were determined. The motility induction was calculated by dividing the swarming diameter of induced versus uninduced strains. The averages of at least three independent experiments (±SD) are provided. For statistical analyses, a two‐tailed Student's t‐test was performed.
SDS‐PAGE and western blotting
Cell lysates were prepared as described previously (Metzger et al., 2016). In brief, after cultivation with or without arabinose for 3 or 6 h, bacterial cell pellets were resuspended in Laemmli buffer, adjusting for the total number of bacteria according to the OD600 values. Proteins were separated by sodium dodecyl sulfate (SDS)‐polyacrylamide gel electrophoresis and western blotted as described (Lo Scrudato and Blokesch, 2012). Primary antibodies against Hcp (Eurogentec) (Metzger et al., 2016), GFP (Roche, Switzerland) and mCherry (BioVision, USA distributed via LubioScience, Switzerland) were used at 1:5000 dilutions, and E. coli Sigma70 (BioLegend, USA distributed via Brunschwig, Switzerland) was used at a 1:10 000 dilution. Goat anti‐rabbit horseradish peroxidase (HRP) and goat anti‐mouse HRP (both diluted 1:20 000; Sigma‐Aldrich, Switzerland) served as secondary antibodies. Lumi‐LightPLUS western blotting substrate (Roche, Switzerland) was used as an HRP substrate and the signals were detected using a ChemiDoc XRS+ station (BioRad).
Author contributions
Conception, design and analysis: L.C.M, N.M., and M.B.; performed research: L.C.M, N.M., C.S., E.J.C. and M.B; wrote the manuscript: L.C.M. and M.B. with input from N.M.
Supporting information
Table S1. Bacterial strains and plasmids used in this study.
Fig. S1. TfoX‐ and TfoY‐induced T6SS production is conserved in pandemic V. cholerae strains. Interspecies killing assay between diverse V. cholerae strains and E. coli as prey. The V. cholerae O1 El Tor strains tested are as follows: A1552, N16961rep (with repaired frameshift mutation in hapR), C6709, E7946 and P27459 as indicated below the graph. Co‐culturing with E. coli occurred on LB agar plates supplemented with arabinose to induce tfoX or tfoY where indicated. The parental strains without inducible copies of the regulatory genes served as a control. Prey recovery is indicated as CFU/ml on the Y‐axis. Bar plots represent the average of three independent biological replicates (± SD). Statistical significance is indicated (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
Fig. S2. TfoY production is translationally but not transcriptionally controlled by c‐di‐GMP in V. cholerae. V. cholerae reporter strains carrying a gene encoding for a translational fusion between TfoY and mCherry (tfoY‐mCherry) with or without a downstream transcriptional reporter gene (gfp) at the gene's native chromosomal locus were genetically manipulated to insert inducible copies vdcA or cdpA into their genome (inside a mini‐Tn7 transposon). Cells were then grown under inducible conditions to increase or decrease intracellular c‐di‐GMP concentrations, as shown by the arrows above the images. Detection of TfoY‐mCherry and GFP occurred through western blotting. The reporter strain lacking the transposon as well as the parental WT served as controls. Detection of σ70 served as a loading control.
Acknowledgements
We thank members of the Blokesch lab and Frédérique Le Roux for fruitful discussions, Nina Vesel for constructing plasmid pGP704‐Sac‐Kan and Sandrine Stutzmann, Julien Chambaud and Tiziana Scrignari for technical assistance. We acknowledge Dor Salomon for providing the V. parahaemolyticus and V. alginolyticus strains, for discussing unpublished data, and for arranging co‐submission. This work was supported by the Swiss National Science Foundation (31003A 162551) and a Consolidator Grant from the European Research Council (ERC; 724630‐CholeraIndex). M.B. is a Howard Hughes Medical Institute (HHMI) International Research Scholar (grant 55008726).
References
- Alcoforado Diniz, J. , Liu, Y.C. , and Coulthurst, S.J. (2015) Molecular weaponry: diverse effectors delivered by the type VI secretion system. Cell Microbiol 17: 1742–1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bao, Y. , Lies, D.P. , Fu, H. , and Roberts, G.P. (1991) An improved Tn7‐based system for the single‐copy insertion of cloned genes into chromosomes of Gram‐negative bacteria. Gene 109: 167–168. [DOI] [PubMed] [Google Scholar]
- Basler, M. (2015) Type VI secretion system: secretion by a contractile nanomachine. Philos Trans R Soc Lond B Biol Sci 370: 20150021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernardy, E.E. , Turnsek, M.A. , Wilson, S.K. , Tarr, C.L. , and Hammer, B.K. (2016) Diversity of clinical and environmental isolates of Vibrio cholerae in natural transformation and contact‐dependent bacterial killing indicative of type VI secretion system activity. Appl Environ Microbiol 18: 2833–2842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bingle, L.E. , Bailey, C.M. , and Pallen, M.J. (2008) Type VI secretion: a beginner's guide. Curr Opin Microbiol 11: 3–8. [DOI] [PubMed] [Google Scholar]
- Blokesch, M. (2012a) A quorum sensing‐mediated switch contributes to natural transformation of Vibrio cholerae . Mob Genet Elements 2: 224–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blokesch, M. (2012b) TransFLP—a method to genetically modify V. cholerae based on natural transformation and FLP‐recombination. J Vis Exp: e3761 10.3791/3761 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blokesch, M. , and Schoolnik, G.K. (2008) The extracellular nuclease Dns and its role in natural transformation of Vibrio cholerae . J Bacteriol 190: 7232–7240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borgeaud, S. , and Blokesch, M. (2013) Overexpression of the tcp gene cluster using the T7 RNA polymerase/promoter system and natural transformation‐mediated genetic engineering of Vibrio cholerae . PLoS One 8: e53952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borgeaud, S. , Metzger, L.C. , Scrignari, T. , and Blokesch, M. (2015) The type VI secretion system of Vibrio cholerae fosters horizontal gene transfer. Science 347: 63–67. [DOI] [PubMed] [Google Scholar]
- Boyer, F. , Fichant, G. , Berthod, J. , Vandenbrouck, Y. , and Attree, I. (2009) Dissecting the bacterial type VI secretion system by a genome wide in silico analysis: what can be learned from available microbial genomic resources? BMC Genomics 10: 104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chimalapati, S. , de Souza Santos, M. , Servage, K. , De Nisco, N.J. , Dalia, A.B. , and Orth, K. (2018) Natural transformation in Vibrio parahaemolyticus: a rapid method to create genetic deletions. J Bacteriol 200: e00032‐18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Church, S.R. , Lux, T. , Baker‐Austin, C. , Buddington, S.P. , and Michell, S.L. (2016) Vibrio vulnificus type 6 secretion system 1 contains anti‐bacterial properties. PLoS One 11: e0165500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cianfanelli, F.R. , Monlezun, L. , and Coulthurst, S.J. (2016) Aim, load, fire: the type VI secretion system, a bacterial Nanoweapon. Trends Microbiol 24: 51–62. [DOI] [PubMed] [Google Scholar]
- Costa, T.R.D. , Felisberto‐Rodrigues, C. , Meir, A. , Prevost, M.S. , Redzej, A. , Trokter, M. , and Waksman, G. (2015) Secretion systems in Gram‐negative bacteria: structural and mechanistic insights. Nat Rev Microbiol 13: 343–359. [DOI] [PubMed] [Google Scholar]
- Das, S. , and Chaudhuri, K. (2003) Identification of a unique IAHP (IcmF associated homologous proteins) cluster in Vibrio cholerae and other proteobacteria through in silico analysis. In Silico Biol 3: 287–300. [PubMed] [Google Scholar]
- De Souza Silva, O. , and Blokesch, M. (2010) Genetic manipulation of Vibrio cholerae by combining natural transformation with FLP recombination. Plasmid 64: 186–195. [DOI] [PubMed] [Google Scholar]
- Domman, D. , Quilici, M.L. , Dorman, M.J. , Njamkepo, E. , Mutreja, A. , Mather, A.E. , et al (2017) Integrated view of Vibrio cholerae in the Americas. Science 358: 789–793. [DOI] [PubMed] [Google Scholar]
- Dunlap, P.V. (1989) Regulation of luminescence by cyclic AMP in cya‐like and crp‐like mutants of Vibrio fischeri . J Bacteriol 171: 1199–1202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durand, E. , Cambillau, C. , Cascales, E. , and Journet, L. (2014) VgrG, Tae, Tle, and beyond: the versatile arsenal of type VI secretion effectors. Trends Microbiol 22: 498–507. [DOI] [PubMed] [Google Scholar]
- Ferrieres, L. , Hemery, G. , Nham, T. , Guerout, A.M. , Mazel, D. , Beloin, C. , and Ghigo, J.M. (2010) Silent mischief: bacteriophage Mu insertions contaminate products of Escherichia coli random mutagenesis performed using suicidal transposon delivery plasmids mobilized by broad‐host‐range RP4 conjugative machinery. J Bacteriol 192: 6418–6427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gode‐Potratz, C.J. , and McCarter, L.L. (2011) Quorum sensing and silencing in Vibrio parahaemolyticus . J Bacteriol 193: 4224–4237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gode‐Potratz, C.J. , Kustusch, R.J. , Breheny, P.J. , Weiss, D.S. , and McCarter, L.L. (2011) Surface sensing in Vibrio parahaemolyticus triggers a programme of gene expression that promotes colonization and virulence. Mol Microbiol 79: 240–263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graf, J. , Dunlap, P.V. , and Ruby, E.G. (1994) Effect of transposon‐induced motility mutations on colonization of the host light organ by Vibrio fischeri . J Bacteriol 176: 6986–6991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green, E.R. , and Mecsas, J. (2016) Bacterial secretion systems: an overview. Microbiol Spectr 4 10.1128/microbiolspec.VMBF-0012-2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heidelberg, J.F. , Eisen, J.A. , Nelson, W.C. , Clayton, R.A. , Gwinn, M.L. , Dodson, R.J. , et al (2000) DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae . Nature 406: 477–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho, B.T. , Dong, T.G. , and Mekalanos, J.J. (2014) A view to a kill: the bacterial type VI secretion system. Cell Host Microbe 15: 9–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inuzuka, S. , Nishimura, K. , Kakizawa, H. , Fujita, Y. , Furuta, H. , Matsumura, S. , and Ikawa, Y. (2016) Mutational analysis of structural elements in a class‐I cyclic di‐GMP riboswitch to elucidate its regulatory mechanism. J Biochem 160: 153–162. [DOI] [PubMed] [Google Scholar]
- Ishikawa, T. , Rompikuntal, P.K. , Lindmark, B. , Milton, D.L. , and Wai, S.N. (2009) Quorum sensing regulation of the two hcp alleles in Vibrio cholerae O1 strains. PLoS One 4: e6734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaskólska, M. , Stutzmann, S. , Stoudmann, C. , and Blokesch, M. (2018) QstR‐dependent regulation of natural competence and type VI secretion in Vibrio cholerae. Nucleic Acids Res 46: 10619–10634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang, N. , Tang, L. , Xie, R. , Li, Z. , Burkinshaw, B. , Liang, X. , et al (2018) Vibrio parahaemolyticus RhsP represents a widespread group of pro‐effectors for type VI secretion systems. Nat Commun 9: 3899. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Joshi, A. , Kostiuk, B. , Rogers, A. , Teschler, J. , Pukatzki, S. , and Yildiz, F.H. (2017) Rules of engagement: the type VI secretion system in Vibrio cholerae . Trends Microbiol 25: 267–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keene, O.N. (1995) The log transformation is special. Stat Med 14: 811–819. [DOI] [PubMed] [Google Scholar]
- Kernell Burke, A. , Guthrie, L.T. , Modise, T. , Cormier, G. , Jensen, R.V. , McCarter, L.L. , and Stevens, A.M. (2015) OpaR controls a network of downstream transcription factors in Vibrio parahaemolyticus BB22OP. PLoS One 10: e0121863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keyhani, N.O. , and Roseman, S. (1999) Physiological aspects of chitin catabolism in marine bacteria. Biochim Biophys Acta 1473: 108–122. [DOI] [PubMed] [Google Scholar]
- Kimbrough, J.H. , and Stabb, E.V. (2015) Antisocial luxO mutants provide a stationary‐phase survival advantage in Vibrio fischeri ES114. J Bacteriol 198: 673–687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kühn, J. , Finger, F. , Bertuzzo, E. , Borgeaud, S. , Gatto, M. , Rinaldo, A. , and Blokesch, M. (2014) Glucose‐ but not Rice‐based oral rehydration therapy enhances the production of virulence determinants in the human pathogen Vibrio cholerae . PLoS Negl Trop Dis 8: e3347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Roux, F. , and Blokesch, M. (2018) Eco‐evolutionary dynamics linked to horizontal gene transfer in vibrios. Annu Rev Microbiol 72: 89–110. [DOI] [PubMed] [Google Scholar]
- Leiman, P.G. , Basler, M. , Ramagopal, U.A. , Bonanno, J.B. , Sauder, J.M. , Pukatzki, S. , et al (2009) Type VI secretion apparatus and phage tail‐associated protein complexes share a common evolutionary origin. Proc Natl Acad Sci USA 106: 4154–4159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, X. , and Roseman, S. (2004) The chitinolytic cascade in vibrios is regulated by chitin oligosaccharides and a two‐component chitin catabolic sensor/kinase. Proc Natl Acad Sci USA 101: 627–631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livny, J. , Zhou, X. , Mandlik, A. , Hubbard, T. , Davis, B.M. , and Waldor, M.K. (2014) Comparative RNA‐Seq based dissection of the regulatory networks and environmental stimuli underlying Vibrio parahaemolyticus gene expression during infection. Nucleic Acids Res 42: 12212–12223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lo Scrudato, M. , and Blokesch, M. (2012) The regulatory network of natural competence and transformation of Vibrio cholerae . PLoS Genet 8: e1002778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lo Scrudato, M. , and Blokesch, M. (2013) A transcriptional regulator linking quorum sensing and chitin induction to render Vibrio cholerae naturally transformable. Nucleic Acids Res 41: 3644–3658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marvig, R.L. , and Blokesch, M. (2010) Natural transformation of Vibrio cholerae as a tool‐optimizing the procedure. BMC Microbiol 10: 155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthey, N. , and Blokesch, M. (2016) The DNA‐uptake process of naturally competent Vibrio cholerae . Trends Microbiol 24: 98–110. [DOI] [PubMed] [Google Scholar]
- Matthey, N. , Drebes Dörr, N.C. , and Blokesch, M. (2018) Long‐read‐based genome sequences of pandemic and environmental Vibrio cholerae strains. Microbiol Resour Announc 7: e01574‐18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCarter, L.L. (2001) Polar flagellar motility of the Vibrionaceae. Microbiol Mol Biol Rev 65: 445–462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meibom, K.L. , Blokesch, M. , Dolganov, N.A. , Wu, C.‐Y. , and Schoolnik, G.K. (2005) Chitin induces natural competence in Vibrio cholerae . Science 310: 1824–1827. [DOI] [PubMed] [Google Scholar]
- Meibom, K.L. , Li, X.B. , Nielsen, A.T. , Wu, C.Y. , Roseman, S. , and Schoolnik, G.K. (2004) The Vibrio cholerae chitin utilization program. Proc Natl Acad Sci USA 101: 2524–2529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metzger, L.C. , and Blokesch, M. (2016) Regulation of competence‐mediated horizontal gene transfer in the natural habitat of Vibrio cholerae . Curr Opin Microbiol 30: 1–7. [DOI] [PubMed] [Google Scholar]
- Metzger, L.C. , Stutzmann, S. , Scrignari, T. , Van der Henst, C. , Matthey, N. , and Blokesch, M. (2016) Independent regulation of type VI secretion in Vibrio cholerae by TfoX and TfoY. Cell Rep 15: 951–958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milton, D.L. (2006) Quorum sensing in vibrios: complexity for diversification. Int J Med Microbiol 296: 61–71. [DOI] [PubMed] [Google Scholar]
- Miyata, S.T. , Bachmann, V. , and Pukatzki, S. (2013) Type VI secretion system regulation as a consequence of evolutionary pressure. J Med Microbiol 62: 663–676. [DOI] [PubMed] [Google Scholar]
- Nielsen, A.T. , Dolganov, N.A. , Otto, G. , Miller, M.C. , Wu, C.Y. , and Schoolnik, G.K. (2006) RpoS controls the Vibrio cholerae mucosal escape response. PLoS Pathog 2: e109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollack‐Berti, A. , Wollenberg, M.S. , and Ruby, E.G. (2010) Natural transformation of Vibrio fischeri requires tfoX and tfoY . Environ Microbiol 12: 2302–2311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pukatzki, S. , Ma, A.T. , Sturtevant, D. , Krastins, B. , Sarracino, D. , Nelson, W.C. , et al (2006) Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc Natl Acad Sci USA 103: 1528–1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pursley, B.R. , Maiden, M.M. , Hsieh, M.L. , Fernandez, N.L. , Severin, G.B. , and Waters, C.M. (2018) Cyclic di‐GMP regulates TfoY in Vibrio cholerae to control motility by both transcriptional and posttranscriptional mechanisms. J Bacteriol 200: e00578‐17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redfield, R.J. (1991) sxy‐1, a Haemophilus influenzae mutation causing greatly enhanced spontaneous competence. J Bacteriol 173: 5612–5618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell, A.B. , Peterson, S.B. , and Mougous, J.D. (2014) Type VI secretion system effectors: poisons with a purpose. Nat Rev Microbiol 12: 137–148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salomon, D. , Klimko, J.A. , and Orth, K. (2014) H‐NS regulates the Vibrio parahaemolyticus type VI secretion system 1. Microbiology 160: 1867–1873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salomon, D. , Gonzalez, H. , Updegraff, B.L. , and Orth, K. (2013) Vibrio parahaemolyticus type VI secretion system 1 is activated in marine conditions to target bacteria, and is differentially regulated from system 2. PLoS One 8: e61086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salomon, D. , Klimko, J.A. , Trudgian, D.C. , Kinch, L.N. , Grishin, N.V. , Mirzaei, H. , and Orth, K. (2015) Type VI secretion system toxins horizontally shared between marine bacteria. PLoS Pathog 11: e1005128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook, J. , Fritsch, E.F. , and Maniatis, T. (1982) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. [Google Scholar]
- Seitz, P. , and Blokesch, M. (2013) DNA‐uptake machinery of naturally competent Vibrio cholerae . Proc Natl Acad Sci USA 110: 17987–17992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seitz, P. , and Blokesch, M. (2014) DNA transport across the outer and inner membranes of naturally transformable Vibrio cholerae is spatially but not temporally coupled. MBio 5: e01409‐14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seitz, P. , Pezeshgi Modarres, H. , Borgeaud, S. , Bulushev, R.D. , Steinbock, L.J. , Radenovic, A. , et al (2014) ComEA is essential for the transfer of external DNA into the Periplasm in naturally transformable Vibrio cholerae cells. PLoS Genet 10: e1004066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simon, R. , Priefer, U. , and Pühler, A. (1983) A broad host range mobilization system for in vivo genetic engineering: transposon mutagenesis in Gram negative bacteria. Nat Biotechnol 1: 784–791. [Google Scholar]
- Speare, L. , Cecere, A.G. , Guckes, K.R. , Smith, S. , Wollenberg, M. , Mandel, M. , et al (2018) Bacterial symbionts use a type VI secretion system to eliminate competitors in their natural host. Proc Natl Acad Sci USA 115: E8528–E8537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stutzmann, S. , and Blokesch, M. (2016) Circulation of a quorum‐sensing‐impaired variant of Vibrio cholerae strain C6706 masks important phenotypes. mSphere 1: e00098‐16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suckow, G. , Seitz, P. , and Blokesch, M. (2011) Quorum sensing contributes to natural transformation of Vibrio cholerae in a species‐specific manner. J Bacteriol 193: 4914–4924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor, N.M.I. , van Raaij, M.J. , and Leiman, P.G. (2018) Contractile injection systems of bacteriophages and related systems. Mol Microbiol 108: 6–15. [DOI] [PubMed] [Google Scholar]
- Unterweger, D. , Kitaoka, M. , Miyata, S.T. , Bachmann, V. , Brooks, T.M. , Moloney, J. , et al (2012) Constitutive type VI secretion system expression gives Vibrio cholerae intra‐ and interspecific competitive advantages. PLoS One 7: e48320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Henst, C. , Vanhove, A.S. , Drebes Dörr, N.C. , Stutzmann, S. , Stoudmann, C. , Clerc, S. , et al (2018) Molecular insights into Vibrio cholerae's intra‐amoebal host‐pathogen interactions. Nature Commun 9: 3460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veening, J.W. , and Blokesch, M. (2017) Interbacterial predation as a strategy for DNA acquisition in naturally competent bacteria. Nat Rev Microbiol 15: 621–629. [DOI] [PubMed] [Google Scholar]
- Verma, S.C. , and Miyashiro, T. (2013) Quorum sensing in the squid‐Vibrio symbiosis. Int J Mol Sci 14: 16386–16401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto, S. , Izumiya, H. , Mitobe, J. , Morita, M. , Arakawa, E. , Ohnishi, M. , and Watanabe, H. (2011) Identification of a chitin‐induced small RNA that regulates translation of the tfoX gene, encoding a positive regulator of natural competence in Vibrio cholerae . J Bacteriol 193: 1953–1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto, S. , Mitobe, J. , Ishikawa, T. , Wai, S.N. , Ohnishi, M. , Watanabe, H. , and Izumiya, H. (2014) Regulation of natural competence by the orphan two‐component system sensor kinase ChiS involves a non‐canonical transmembrane regulator in Vibrio cholerae . Mol Microbiol 91: 326–347. [DOI] [PubMed] [Google Scholar]
- Yang, Z. , Zhou, X. , Ma, Y. , Zhou, M. , Waldor, M.K. , Zhang, Y. , and Wang, Q. (2018) Serine/threonine kinase PpkA coordinates the interplay between T6SS2 activation and quorum sensing in the marine pathogen Vibrio alginolyticus . Environ Microbiol 20: 903–919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yanisch‐Perron, C. , Vieira, J. , and Messing, J. (1985) Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 33: 103–119. [DOI] [PubMed] [Google Scholar]
- Yildiz, F.H. , and Schoolnik, G.K. (1998) Role of rpoS in stress survival and virulence of Vibrio cholerae . J Bacteriol 180: 773–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yildiz, F.H. , Dolganov, N.A. , and Schoolnik, G.K. (2001) VpsR, a member of the response regulators of the two‐component regulatory systems, is required for expression of vps biosynthesis genes and EPS(ETr)‐associated phenotypes in Vibrio cholerae O1 El tor. J Bacteriol 183: 1716–1726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, Y. , Yang, H. , Li, J. , Zhang, P. , Wu, B. , Zhu, B. , et al (2012) Putative type VI secretion systems of Vibrio parahaemolyticus contribute to adhesion to cultured cell monolayers. Arch Microbiol 194: 827–835. [DOI] [PubMed] [Google Scholar]
- Zoued, A. , Brunet, Y.R. , Durand, E. , Aschtgen, M.S. , Logger, L. , Douzi, B. , et al (2014) Architecture and assembly of the type VI secretion system. Biochim Biophys Acta 1843: 1664–1673. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Bacterial strains and plasmids used in this study.
Fig. S1. TfoX‐ and TfoY‐induced T6SS production is conserved in pandemic V. cholerae strains. Interspecies killing assay between diverse V. cholerae strains and E. coli as prey. The V. cholerae O1 El Tor strains tested are as follows: A1552, N16961rep (with repaired frameshift mutation in hapR), C6709, E7946 and P27459 as indicated below the graph. Co‐culturing with E. coli occurred on LB agar plates supplemented with arabinose to induce tfoX or tfoY where indicated. The parental strains without inducible copies of the regulatory genes served as a control. Prey recovery is indicated as CFU/ml on the Y‐axis. Bar plots represent the average of three independent biological replicates (± SD). Statistical significance is indicated (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001).
Fig. S2. TfoY production is translationally but not transcriptionally controlled by c‐di‐GMP in V. cholerae. V. cholerae reporter strains carrying a gene encoding for a translational fusion between TfoY and mCherry (tfoY‐mCherry) with or without a downstream transcriptional reporter gene (gfp) at the gene's native chromosomal locus were genetically manipulated to insert inducible copies vdcA or cdpA into their genome (inside a mini‐Tn7 transposon). Cells were then grown under inducible conditions to increase or decrease intracellular c‐di‐GMP concentrations, as shown by the arrows above the images. Detection of TfoY‐mCherry and GFP occurred through western blotting. The reporter strain lacking the transposon as well as the parental WT served as controls. Detection of σ70 served as a loading control.