
Figure 1.
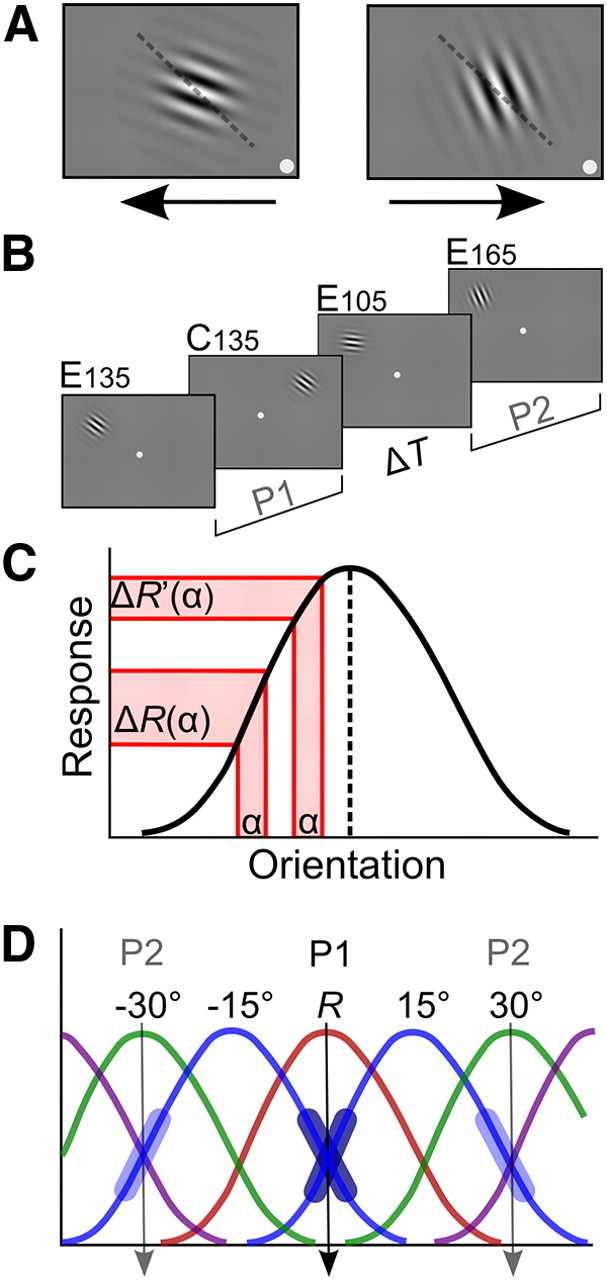
ODT and training-induced tuning changes in single neurons. A, Gabor stimuli and task. Participants fixated in the center of the screen while covertly attending the Gabor patch in an upper (left/right) quadrant of the visual field. Participants indicated the clockwise or anticlockwise deviation of the stimulus relative to a never-presented reference (in this case 135°; dashed line shows reference for illustrative purposes) by pressing the right or left arrow key on a computer keyboard. Feedback was given by a change in color of the fixation dot (green, correct; red, incorrect). Trials with fixation errors (>1.5° from fixation) were aborted (Arrington Research), and 84% correct staircase JNDs were determined (implemented in CORTEX 5.9.6, NIH freeware). B, Testing paradigm. Participants were tested in four conditions (4 JNDs of ∼100 trials per condition). In training period P1 participants were trained at a 135° reference orientation in upper left and right quadrant. One upper quadrant served as a control (C135). The other was the experimental quadrant (E135), where P1 training after a variable time interval of rest (ΔT) was followed by training in a second period P2 at reference orientations 105° (E105) and 165° (E165). C, Tuning curve of an orientation selective cell is most discriminative at its flanks. A difference α between two orientations on the flanks of the tuning curve will yield a differential response ΔR(α) that is larger than the differential response ΔR′(α) yielded by the same difference between two orientations closer to the neuron's preferred orientation (dashed line). D, Our experimental design is informed by prior research showing that extensive training in orientation discrimination invoking asymptotic performance (Vogels and Orban, 1985; Schoups et al., 1995) selectively modified flank slopes of tuning curves in V1 and V4 neurons (Schoups et al., 2001; Raiguel et al., 2006). As V1 neurons are more narrowly tuned to orientation (David et al., 2006), and are probably more crucial during final asymptotic performance, we used V1 tuning properties to predict conditions of behavioral interference. In a V1 population neurons with tuning curve peaks about 15° away (blue curves) from a trained reference orientation R (135°) (red curve) sharpen their tuning curve flanks (fat dark blue line segments) overlapping with the reference orientation. Extrapolating this idea, we predicted that following up P1 training at the reference orientation R with subsequent, P2 training at R ± 30° (green curves) would cause interference with training at R. P1 training at R and subsequent P2 training at R ± 30° both require plasticity in neurons with preferred orientations at R ± 15°, P1 training should increase the slope of flanks in tuning curves overlapping with R, and P2 training the slope of flanks overlapping with the +30° and −30° orientations (fat light blue line segments). We hypothesized that one requirement would counteract the other, leading to behavioral interference at R [P2 training is also expected to lead to steepening of flanks of ±45° neurons' tuning curves (in purple)].