

Abstract
Background
Onchocerciasis currently afflicts an estimated 15 million people and is the second leading infectious cause of blindness world-wide. The development of a macrofilaricide to cure the disease has been hindered by the lack of appropriate small laboratory animal models. This study therefore, was aimed at developing and validating the Mongolian gerbil, as an Onchocerca ochengi (the closest in phylogeny to O. volvulus) adult male worm model.
Methodology/Principal findings
Mongolian gerbils (Meriones unguiculatus) were each implanted with 20 O. ochengi male worms (collected from infected cattle), in the peritoneum. Following drug or placebo treatments, the implanted worms were recovered from the animals and analyzed for burden, motility and viability. Worm recovery in control gerbils was on average 35%, with 89% of the worms being 100% motile. Treatment of the gerbils implanted with male worms with flubendazole (FBZ) resulted in a significant reduction (p = 0.0021) in worm burden (6.0% versus 27.8% in the control animals); all recovered worms from the treated group had 0% worm motility versus 91.1% motility in control animals. FBZ treatment had similar results even after four different experiments. Using this model, we tested a related drug, oxfendazole (OFZ), and found it to also significantly (p = 0.0097) affect worm motility (22.7% versus 95.0% in the control group).
Conclusions/Significance
We have developed and validated a novel gerbil O. ochengi adult male worm model for testing new macrofilaricidal drugs in vivo. It was also used to determine the efficacy of oxfendazole in vivo.
Author summary
Onchocerciasis is a neglected tropical disease caused by the filarial nematode Onchocerca volvulus. The only drug currently used for the treatment of this disease and reduction of transmission is ivermectin. However, this drug kills only the microfilarial stage of the parasite, and not the adult worm, and cases of parasite resistance to the drug have been recorded. Therefore, there is an urgent need to find an efficacious macrofilaricidal drug that targets the adult worms. To identify an effective macrofilaricide, suitable small laboratory animal models are needed for routine drug screening and/or validation in vivo. To develop such a model, we implanted Mongolian gerbils with adult O. ochengi male worms; O. ochengi is the closest filarial relative of O. volvulus. Gerbils appeared to be permissive to O. ochengi male worms and when treated with flubendazole, a known macrofilaricidal drug, the drug significantly decreased worm burden, motility and viability when compared to vehicle-treated animals. This model was also used to test the efficacy of oxfendazole, a drug belonging to same class as flubendazole, and it also showed potent activity on O. ochengi male worms. The gerbil O. ochengi male worm model is now available to screen new candidate macrofilaricides.
Introduction
Onchocerciasis currently afflicts an estimated 15 million people worldwide, predominantly in Sub-Saharan Africa [1]. It is the second leading infectious cause of blindness globally [2]. Its causative agent, Onchocerca volvulus is transmitted by multiple bites from the infected black fly (of genus Simulium). The adult worms of this tissue dwelling filaria, establish in subcutaneous nodules (onchocercoma), producing millions of microfilariae (mf) which parasitize skin and eye tissues. Host inflammatory and immune responses to this parasite and antigens from the mf and adult worm stages result in major and sometimes severe pathologies; which include intense itching, dermatitis, atrophy, visual impairment and blindness [3, 4]. Host response to antigens of Wolbachia (the endosymbiont of Onchocerca) has also been associated with eye pathology [5].
Current efforts to control onchocerciasis are almost exclusively dependent on controlling transmission using mass distribution of ivermectin. This has also been successful in reducing the public health burden of both ocular and dermal manifestations of the disease with >65.3% global coverage [6]. However, sole reliance on ivermectin is not without its limitations as recent reports indicate continuous evolution of ocular onchocerciasis even after 17 years of consistent ivermectin treatment [7]. Furthermore, apart from being only a microfilaricide, unresponsiveness of O. volvulus to ivermectin treatment in some areas of West Africa have emerged [8]. As a result, attempts to use ivermectin in combination with the antibiotic doxycycline have been made, resulting in long-term sterility of adult female worms and a corresponding absence of mf [9, 10]. Other studies have shown the possibility of achieving macrofilaricidal activity after 6 weeks treatment with 200 mg/day doxycycline [11, 12]. Though significant macrofilaricidal activity has been observed with doxycycline [13–15], a prolonged (6 weeks) daily treatment serves as contraindication in pregnant or breastfeeding women, and children <8 years old [16]. This impedes the use of this class of anti-Wolbachia drugs in mass drug administration. Other drugs like albendazole, flubendazole, and oxytetracycline have been implicated in the treatment of onchocerciasis [17, 18] with oxytetracycline and flubendazole showing good macrofilaricidal activity against O. ochengi [13, 19]. However, wider testing of flubendazole in humans was restricted 30 years ago, as it was poorly tolerated and poorly absorbed from the gut given its formulation [2, 20]. In addition, its intramuscular route of administration results in painful sterile abscesses. Recently, Janssen announced the discontinuation of the development of an orally bioavailable formulation (amorphous solid dispersion) of flubendazole due to safety concerns [21, 22]. Given the aforementioned attributes and serious limitations of the current drugs, it is unlikely that eradication, or even stable elimination, can be achieved without a safe and effective macrofilaricide.
The potential development of such a macrofilaricide is mired by the lack of an appropriate pre-clinical small laboratory animal model to evaluate drug candidates in vivo [23]. The only small laboratory animal model developed and validated for adult O. ochengi reported till date is the SCID (Severe Combined Immuno-Deficient) mouse model, which is immunodeficient [24]. In this model, the role of the acquired immunity in parasite killing is not considered, necessitating more suitable models. In the SCID mice model, viable adult male worms were recovered 35 days post-implantation [24], indicating that the male worms can survive long enough to see the effect of slow acting compounds. For over 2 decades, the Mongolian gerbil (Meriones unguiculatus) has been known to have a uniquely high susceptibility to various parasitic infections, including filarial parasites [25]. It is currently used as an animal model for Brugia and Litomosoides sigmodontis [26–28] and has also been implicated in the study of giardiasis and schistosomiasis [29, 30]. Furthermore, immunocompetent gerbil has recently been shown to harbor adult male O. ochengi worms for 42 days but drugs which directly target the worms were not used in the study [31]. Given the long-standing use of gerbils as a model for other parasitic infections, we investigated it as a model for testing and validating new macrofilaricidal drugs against O. ochengi adult male worms.
Methods
Ethics statement
The animal protocol (UB-IACUC No 002/2017) was approved by the University of Buea Animal Care and Use Committee, following recommendations from the ‘Guide for the Care and Use of Laboratory Animals’, 8th edition by the National Research Council, USA. This in vivo study was reported in accordance with the ARRIVE Guidelines for reporting animal research.
Experimental animals
Male and female Mongolian gerbils (Meriones unguiculatus, (≥ 6 weeks old)) were purchased from Charles River (France). These animals were maintained in a conventional animal house at the Biotechnology Unit of the University of Buea, and given food and water ad libidum. Using the ClinCalc sample size calculator, the minimum number of animals suggested was 4 animals per group. However, the IACUC regulations require that the minimum number of animals able to give meaningful statistics be used and therefore we used at least 5 animals (3 exceptionally for the positive control group) per group in respect of these basic tenets.
Isolation of O. ochengi adult male worms from cattle skin and implantation in gerbils
Worms were isolated from umbilical cattle skin by the method of Cho-Ngwa et al., (2010) [32]. Fresh pieces of cattle skin with palpable nodules were purchased from a local slaughter house, washed repeatedly with tap water and rinsed with distilled water. The skin was towel-dried, sterilized with 70% ethanol, and allowed to dry in a laminar flow hood. Using a razor blade, the pale orange-yellow O. ochengi adult worm masses were excised with care not to hurt the adult worm. The entire mass was subsequently submerged in incomplete culture medium (ICM); RPMI-1640 (SIGMA, USA), supplemented with 25 mM HEPES, 2 g/L sodium bicarbonate, 2 mM L-glutamine, 200 units/ml penicillin, 200 μg/ml streptomycin and 0.25 μg/ml amphotericin B, pH 7.4 in 24-well tissue culture plates. Damaged worms and worms from putrefied nodules were discarded. The plates containing the worms were then incubated at 37°C in humidified air with 5% CO2 overnight allowing male worms to emerge. Thereafter, the viability of the male worms was evaluated microscopically using an inverted microscope (Nikon Eclipse TS100) before implantation. For implantation, the gerbils were sedated with 80/5 mg/kg body weight ketamine/xylazine. The abdominal area of each animal was shaved and sterilized with 70% ethanol. Using a size 23 sterile surgical blade (Everich, China), a small incision (~1 cm) was made on the skin and peritoneal membrane of each animal.
Twenty (20) male worms were implanted in the peritoneum of each gerbil. Using a sterile nylon monofilament non-absorbable suture needle from Medtrue Enterprise (Nigeria), interrupted stitches were made. The peritoneal membrane was stitched separately from skin, to avoid displacement of the worms from the peritoneum. The stitched area was sterilized with 70% ethanol; animals were placed in cages containing freshly sterilized beddings and monitored daily.
Drug administration
Flubendazole (FBZ) (cat# 34091) and oxfendazole (OFZ) (cat# 34176) were obtained from Sigma-Aldrich (Germany). Both drugs were administered subcutaneously, with FBZ given at a dose of 10 mg/kg body weight, SID, and OFZ given at 12 mg/kg body weight, BID, for 5 days starting 3 days post-implantation. The vehicle used for both drugs was composed of 0.5% HEC (hydroxyethyl cellulose), 0.1% Tween 80, and 99.4% water.
Recovery of O. ochengi adult worms from gerbils
Gerbils were sacrificed via cranial dislocation 35 days post-implantation of O. ochengi adult male worms. After opening the peritoneum, the cavity was thoroughly observed for the presence of adult male worms, either free or in nodules. Male nodules found in the peritoneum were introduced into collagenase B at a concentration of 0.5 mg/ml and incubated for at least 2 hours at 37°C. After incubation, all worms were observed under the microscope and observations recorded. For all confirmatory experiments, analyses of motility were performed in a blinded manner, where animal groups were only decoded after all the worms had been recovered and scored.
Assessing the viability of O. ochengi male worms recovered from gerbils
The motility of adult male worms was evaluated using an inverted microscope. Motility scores ranged from 100% (no observable reduction in motility), 90% (very minute reduction in motility), 75% (slight reduction in motility), 50% (general sluggish movement), and 25% (movement in only head or tail), to 0% (no movement).
Viability of male worms was assessed by MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide] staining. Each recovered male worm was incubated in 500 μl of 0.5 mg/ml MTT] for 30 min, transferred onto an absorbent paper, and electronic images captured against a white background. The individual male worms were then decolorized by incubating each at 37°C in 200 μl of DMSO for 30 min. The optical density (OD) was measured at 595 nm using a microplate reader (Molecular Devices, USA).
Data analyses
Data were analyzed using GraphPad Prism 5.0 software and statistical analyses were performed using the Mann-Whitney non-parametric test for two groups of data or Kruskal-Wallis test with Dunn’s multiple comparison post-test for three groups of data. Data presented show mean ± SEM. Data were considered significant if the differences were p <0.05.
Results
Ratio of O. ochengi adult male worms to worm masses
Cattle skin naturally infected with O. ochengi was obtained from slaughter houses in Douala, Littoral Region and Buea, South West Region of Cameroon. Each visit to the slaughter house yielded an average of 212.2 ± 28.0 (range: 111–544) worm masses from infected cattle skin, from which an average of 127.6 ± 12.7 (range: 55–250) adult male worms were recovered. Except for one case, the number of male worms obtained was always less than the number of worm masses recovered (Fig 1A). The average ratio of worm masses to liberated adult male worms was 1:0.6. The number of liberated male worms correlated with number of worm masses from which they emerged, Pearson r = 0.8171; p = 0.0002 (Fig 1B), although a single data point was much higher than the rest of the data. Based on our experience, on average, ~350 worm masses can be harvested per week which yields ~210 adult male worms.
Fig 1. Correlation between the number of worm masses and the number of liberated adult O. ochengi male worms.

Total number of worm masses obtained from cattle skin each time (n = 15) and the corresponding number of liberated male worms, A) Paired comparison of the number of worm masses and number of liberated male worms, B) Correlation between the number of worm masses to liberated male worms. Line shows linear regression best fit (Pearson r = 0.8171; p = 0.0002).
Comparable recovery of O. ochengi adult male worms in male and female gerbils
To see if there was a sex bias in the survival of adult O. ochengi male worms in gerbils, both male and female animals were infected with 20 male worms. Gerbils were analyzed 35 days post-implantation. It was found that 6.8 ± 1.2 adult male worms were recovered from male gerbils, while 7.3 ± 0.8 worms were recovered from female gerbils (Fig 2). Thus, the survival of O. ochengi adult male worms in male and female gerbils was not significantly different, allowing for both sexes to be used indiscriminately in this study. Considering both male and female gerbils, the average recovery was 7.0 ± 0.8 which is equivalent to 35%.
Fig 2. Comparison of O. ochengi male worm recovery in male and female gerbils.
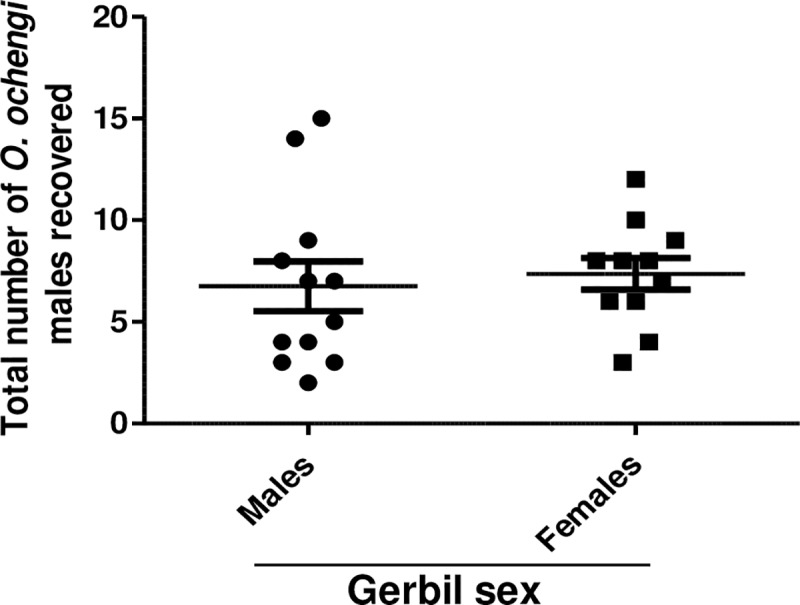
Gerbils were implanted with 20 O. ochengi males, sacrificed 35 days later and analyzed for total number of male worms recovered in male and female gerbils (n = 11–12 per group).
O. ochengi adult male worms in gerbil peritoneum were either moving freely or encapsulated in nodules
Fig 3A shows O. ochengi adult male worms that emerged in vitro from O. ochengi worm masses after 24 hours in culture. Upon sacrifice of untreated gerbils 35 days post-implantation, it was noticed that some of male worms were found free and could be seen moving in the peritoneum (Fig 3B) while other male worms were encapsulated within newly formed nodules (Fig 3C). The encapsulated male worms were released by digestion of the nodules with collagenase. Both free and nodular O. ochengi male worms had no preferred location and could be found anywhere in the peritoneum.
Fig 3. Recovery of free or nodular O. ochengi adult male worms within the peritoneum of gerbils.

A) Liberated O. ochengi adult male worms in vitro from worm masses removed from infected cattle skin, B) Cluster of free O. ochengi adult male worms in the peritoneum of gerbils (circle) 35 days post-implantation, C) Nodules containing O. ochengi adult male worms in the peritoneum of gerbils (arrow head) 35 days post-implantation, D) Percentage of free and nodular O. ochengi adult male worms in the peritoneum of gerbils 35 days post-implantation. Analyses showed that 61.2 ± 13.6% of the male worms recovered were free in the peritoneum while 38.8 ± 13.6% were in nodules (Fig 3D). There was no statistically significant difference in the motility of male worms recovered from nodules or found free in the peritoneum of control groups (S1 Table). Eighty nine (89)% of the O. ochengi male worms recovered from the exploratory gerbils scored 100% motility.
Validation of the gerbil O. ochengi adult male worm model with flubendazole (FBZ)
To validate the model, we treated gerbils implanted with O. ochengi adult male worms with the potent macrofilaricide; FBZ, and compared with vehicle-treated gerbils. FBZ treatment significantly decreased (p = 0.0021) the number of recovered male worms in gerbils (1.3 ± 0.5), compared to the number (5.5 ± 0.8) in control animals (Fig 4A). In addition, there was a complete inhibition of the motility of the recovered adult male worms from the FBZ-treated gerbils whereas the motility in control animals was 91.1 ± 3.9%, p = 0.0019 (Fig 4B). Pictures of MTT-stained adult male worms from control animals clearly showed that the worms were viable as they stained dark blue as opposed to worms from FBZ-treated animals which had a faint blue coloration (Fig 4C). Measuring the OD of DMSO used to decolorize MTT-stained male worms, showed a significantly reduced OD (p = 0.0007) for worms from FBZ-treated animals (0.07 ± 0.01) compared to worms from control gerbils (0.20 ± 0.02) (Fig 4D).
Fig 4. Flubendazole (FBZ) decreases O. ochengi male worm burden and motility in gerbils.

Gerbils were implanted with 20 O. ochengi male worms and treated for 5 days subcutaneously starting on day 3 post-implantation with 10 mg/kg FBZ, SID. Animals were sacrificed on day 35 post-implantation. A) Total number of male worms recovered, B) Percentage motility of the male worms recovered, C) Photo of O. ochengi male worms from control and FBZ-treated gerbils after MTT staining, D) OD of DMSO treated male worms after MTT staining, E) Blood clot in the lower peritoneal cavity of FBZ-treated animal (circle), and F) Picture of normal and swollen Peyer’s patches (arrow head). Results are representative of five independent experiments. N = 8 per group. Statistical significance was done by Mann Whitney test. **p <0.01; ***p <0.001.
More so, 80% (16/20) of FBZ-treated gerbils had blood clots in their abdominal cavities (Fig 4E) while such clots were not observed in control gerbils (0/18). Some gerbils had very prominent Peyer’s patches (Fig 4F). Though not significantly different (p = 0.4392), the occurrence of prominent Peyer’s patches was higher in the FBZ-treated group; 67% (6/9), compared with the control group; 43% (3/7).
Oxfendazole (OFZ) significantly reduces worm motility
Using the validated O. ochengi adult male worm gerbil model, we tested the effect of OFZ; a drug similar to FBZ, that is used as an anthelmintic in livestock. A total of 8.7 ± 1.6 male worms were retrieved from the control gerbils while 4.0 ± 1.0 were found in OFZ-treated animals (Fig 5A). Of the total male worms recovered, 7.8 ± 2.0 were motile in the control group, which was significantly higher (p = 0.0073), compared to the 1.4 ± 0.2 motile worms recovered from the OFZ-treated group (Fig 5B). The percent motility of worms (93.0 ± 4.0) in the control group was equally significantly higher (p = 0.0097), compared to the worms in the OFZ-treated group (20.0 ± 14.0) (Fig 5C). Similarly, the OD after MTT staining of O. ochengi male worms from control gerbils (0.19 ± 0.01) was higher (but not significant; p = 0.0519), compared to worms from OFZ-treated gerbils (0.10 ± 0.03) (Fig 5D). The percent male worms within nodules in the OFZ-treated group was 70.0 ± 12.2 as opposed to 38.8 ± 13.6 (p = 0.0530) in the control group (Fig 5E). FBZ was used as positive control and its effects on worm burden and viability were more severe, compared with OFZ-treated gerbils (Fig 5A–5D). Interestingly, all the worms recovered in the FBZ-treated group were retrieved from newly formed nodules (Fig 5E). The OFZ-treated gerbils equally had blood clots in the peritoneum, similar to FBZ-treated animals.
Fig 5. Oxfendazole (OFZ) decreases the number of O. ochengi motile male worms and their viability.

Gerbils were implanted with 20 O. ochengi male worms and treated for 5 days starting on day 3 post-implantation subcutaneously with 12 mg/kg OFZ, BID, or with 10 mg/kg FBZ, SID. Animals were sacrificed on day 35 post-implantation. A) Total number of male worms recovered, B) Number of viable male worms recovered, C) Percent motility of the male worms recovered, D) OD of DMSO treated male worms after MTT staining, and E) Percent of O. ochengi male worms within the nodules. FBZ was used as positive control. N = 3–6 per group. Statistical significance between groups was done using Kruskal-Wallis Test with Dunn’s multiple comparison. *p <0.05; **p <0.01.
Discussion
In this study a small animal model for the screening of macrofilaricide activity against onchocerciasis was developed. Mongolian gerbils were implanted with O. ochengi adult male worms and shown to be a validated model for testing known macrofilaricidal drugs such as flubendazole. Such a model represents a great contribution in onchocerciasis preclinical drug development, as drug candidates identified using in vitro screening assays can now be tested in this model. This will also allow a better prediction of the activity of new drugs against the human O. volvulus in vivo. The macrofilaricide; FBZ, was used to validate this model. The burden of adult male O. ochengi worms implanted in FBZ-treated gerbils was assessed on day 35 post-implantation and showed a significant decrease (78%); the number of recovered worms was 5.5 ± 0.8 in control gerbils and 1.3 ± 0.5 in FBZ-treated gerbils (Fig 4A). Remarkably, worm motility was completely inhibited in the FBZ-treated gerbils, but 91.1 ± 3.9% in the controls (Fig 4B). Using this model, OFZ used in livestock as an anthelmintic drug, was equally tested. Although there was no significant difference in the total number of worms recovered from the animals in each group (Fig 5A), in the OFZ-treated gerbils, the recovered viable worms had significantly reduced motility (p = 0.0073) (Fig 5C). This gerbil model is the first adult Onchocerca model described using immunocompetent animals and therefore promises to be a very robust model. Further analyses are required to better understand the role of the immune system in this model, which is probably also drug-dependent.
The effect of drugs on the parasite depends on many variables including the parasite species, the specific host, the drug concentration, formulation, route of administration, and other assay conditions. In addition, PK/PD varies from one host to another. Moreover, the difference in the location of the parasite within its natural human host and in this particular gerbil model (peritoneum) may also lead to some differences in pharmacokinetics and therefore, drug efficacy. Based on our results, OFZ is not as good as FBZ although it was given at a higher dose (twice daily at 12 mg/kg body weight while FBZ was given once daily at 10 mg/kg). Interestingly, in OFZ- and FBZ-treated gerbils, almost all the recovered worms were non-motile (Figs 4B and 5C).
Although a SCID mouse model of O. ochengi adult male worms has equally been described [24], our study used gerbils that are immunocompetent, and thus the possible synergistic involvement of the acquired immune response to the killing of the parasites can equally be evaluated in this model [31]. Ivermectin and diethylcarbamazine (DEC) have been shown to have no effect or require much higher concentrations for microfilaricidal activity in vitro than in vivo [33–35] supporting a synergistic role between the drug activity and the immune system. Interestingly, 43% of the control gerbils developed swollen Peyer’s patches after male O. ochengi implantation, indicating some involvement of the immune response; related to the effect of the drugs on the male worms, and/or because of the release of parasite proteins once the worms are killed. Gerbils have also been shown to have not only cellular, but also humoral immune responses to filarial antigens [36, 37].
The susceptibility of the gerbils to implanted male O. ochengi was not sex dependent as comparable numbers of worms were retrieved from both males and females (Fig 2). This makes it easier to get sufficient number of animals for experiments as both sexes can be used indiscriminately. However, survival of certain filarial nematodes has been shown to be animal sex dependent. For example, male gerbils are more susceptible to B. malayi than females [38]. Another nematode, L. sigmondontis, survives better in female than in male Balb/c mice [39].
This gerbil model with O. ochengi male worms we have developed is a good model for screening especially slow acting (requiring 30 or more days to act) drugs. The results obtained on day 35 post-implantation showed 89% of the male worms at 100% viability. This high viability indicates that the worms can survive beyond this time point. Killing the male worms will prevent the fertilization of female eggs, thereby preventing mf production. Although killing the adult female worm is preferred, such a model for O. ochengi is not yet available. Results from our laboratory showed that female O. ochengi worms do not survive for long in rodents commonly used in laboratory studies, including gerbils. The only published report of viable female O. ochengi worms was in SCID mice sacrificed 7 days post-implantation [24]. Even though female O. volvulus survives in SCID mice or athymic nude rats for up to 20 weeks post-implantation [40], such a model cannot be used for routine screening. Although closely related, O. ochengi and O. volvulus may present with different sensitivities. However, the closeness in phylogeny has proven to work in the past with ivermectin for example, clearing O. ochengi microfilariae in cattle, with the same efficiency with which it clears O. volvulus microfilariae in humans. O. ochengi has also been shown to share over 95% conserved regions with O. volvulus [41], this presents with a very slim chance of a potential drug molecule not to have similar effects in both species. More so, it will be too costly to get enough O. volvulus worms from humans for routine screens.
Interestingly, 38.8 ± 13.6% of the free male worms implanted in untreated animals were found in newly formed nodules, while 61.2 ± 13.6% of the male worms recovered were free in the peritoneum (Fig 3D). Although it is still unclear why some of these worms induce the formation of nodules in the peritoneum, the immune system would most likely play a role in worm recovery, and the formation of the nodule could be the result of an immune response to the wall of the infection, and/or response to the angiogenic factors the implanted worms secrete. The viable worms recovered from the nodules were generally as motile as the free worms found in the peritoneum (S1 Table). We did not find any reason that could suggest that formation of the nodules was linked to degradation of dead worms. It is therefore important to do a meticulous peritoneal search when analyzing the animals, to avoid missing any free males. Furthermore, the free males may be more exposed and thereby more susceptible to the drugs, as 100 ± 0.0% and 70.0 ± 12.2% of worms retrieved from FBZ- and OFZ-treated animals respectively, were recovered from nodules, as opposed to 38.8 ± 13.6% recovered from nodules in control animals (Fig 5E).
FBZ and OFZ belong to the class of benzimidazoles which are aromatic organic compounds used for decades as anthelmintics [42]. These β-tubulin binding drugs inhibit worm motility, reproduction and cell secretory processes [43, 44]. FBZ was very effective in validating this model (Fig 4A–4D). This is in line with observations by Halliday et al., (2014). In addition, a modified FBZ (UMF-078) was shown to be effective against infection with O. ochengi in naturally infected cattle [45]. Using the present gerbil model, it was found that OFZ had significant activity on O. ochengi male worms (Fig 5), howbeit less efficient than FBZ. OFZ was administered twice daily at 12 mg/kg body weight while FBZ was given once daily at 10 mg/kg, but both had similar activity, if not higher with FBZ. In recent studies [46] comparing the in vitro activity of FBZ and OFZ on O. volvulus, L5s have equally shown OFZ to be less effective. For instance, both drugs presented a similar IC50 (0.13 μM) for the inhibition of motility, which differed when the inhibition of viability was determined; 4 times higher with OFZ, compared with FBZ (0.54 μM vs. 0.13 μM). This further proves that the kinetics or activity is different for both drugs and may equally demand optimized in vivo timing. Perhaps for drugs like OFZ, a longer time point (>35 days) will result in complete killing of the male worms. With the recent good tolerance shown by OFZ in a Phase 1 study [47], it remains a promising drug to treat onchocerciasis in humans.
In conclusion, we have developed and validated an immunocompetent gerbil O. ochengi male worm model to aid the development of novel Onchocerca macrofilaricides. Using this model, we found that OFZ, a drug belonging to the same class as FBZ, was equally a potential macrofilaricidal drug.
Supporting information
All free worms were scored for motility before the nodular ones. The general motility score was similar in all the worms (almost always 100%). Gerbil SN 4 and 5 clearly illustrate this with 100% motility of all worms recovered, whether free or from nodules (p = 0.0931).
(PPTX)
Acknowledgments
We thank Gamua Stanley and the staff of Biotechnology Unit, Faculty of Science, University of Buea, Cameroon for their technical support.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported by grants from the Bill and Melinda Gates Foundation (www.gatesfoundation.org, grant numbers OPP1098475 and OPP1017584) awarded to FCN, JS, and SL. The funders had no role in the study design, data collection and analyses, decision to publish, or preparation of the manuscript.
References
- 1.Disease GBD, Injury I, Prevalence C. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet (London, England). 390(10100):1211–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Babalola EO. Ocular onchocerciasis: current management and future prospects. Clinical Ophthalmology. 2011; 5:1479–91. 10.2147/OPTH.S8372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wu Y, Egerton G, McCarthy JS, Nutman TB, Bianco AE. Human immune responses to infective stage larval-specific chitinase of filarial parasite, Onchocerca volvulus, Ov-CHI-1. Filaria journal. 2003; 2(1):1 10.1186/1475-2883-2-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Burnham G. Onchocerciasis. Lancet. 1998. May 2; 351(9112):1341–6. 10.1016/S0140-6736(97)12450-3 [DOI] [PubMed] [Google Scholar]
- 5.Saint André Av, Blackwell NM, Hall LR, Hoerauf A, Brattig NW, Volkmann L, et al. The role of endosymbiotic Wolbachia bacteria in the pathogenesis of river blindness. Science. 2002; 295(5561):1892–5. 10.1126/science.1068732 [DOI] [PubMed] [Google Scholar]
- 6.African Programme for Onchocerciasis Control: progress report, 2014–2015. Releve epidemiologique hebdomadaire / Section d'hygiene du Secretariat de la Societe des Nations = Weekly epidemiological record / Health Section of the Secretariat of the League of Nations. 2015. December 4; 90(49):661–74. [PubMed] [Google Scholar]
- 7.Katabarwa MN, Eyamba A, Nwane P, Enyong P, Yaya S, Baldiagaï J, et al. Seventeen years of annual distribution of ivermectin has not interrupted onchocerciasis transmission in North Region, Cameroon. The American journal of tropical medicine and hygiene. 2011; 85(6):1041–9. 10.4269/ajtmh.2011.11-0333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Osei-Atweneboana MY, Awadzi K, Attah SK, Boakye DA, Gyapong JO, Prichard RK. Phenotypic Evidence of Emerging Ivermectin Resistance in Onchocerca volvulus. PLoS neglected tropical diseases. 2011; 5(3):e998 10.1371/journal.pntd.0000998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hoerauf A, Mand S, Volkmann L, Büttner M, Marfo-Debrekyei Y, Taylor M, et al. Doxycycline in the treatment of human onchocerciasis: kinetics of Wolbachia endobacteria reduction and of inhibition of embryogenesis in female Onchocerca worms. Microbes and Infection. 2003; 5(4):261–73. [DOI] [PubMed] [Google Scholar]
- 10.Taylor MJ, Hoerauf A. A new approach to the treatment of filariasis. Current opinion in infectious diseases. 2001; 14(6):727–31. [DOI] [PubMed] [Google Scholar]
- 11.Hoerauf A, Specht S, Büttner M, Pfarr K, Mand S, Fimmers R, et al. Wolbachia endobacteria depletion by doxycycline as antifilarial therapy has macrofilaricidal activity in onchocerciasis: a randomized placebo-controlled study. Medical microbiology and immunology. 2008; 197(3):295–311. 10.1007/s00430-007-0062-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Turner JD, Tendongfor N, Esum M, Johnston KL, Langley RS, Ford L, et al. Macrofilaricidal activity after doxycycline only treatment of Onchocerca volvulus in an area of Loa loa co-endemicity: a randomized controlled trial. PLoS neglected tropical diseases. 2010; 4(4):e660 10.1371/journal.pntd.0000660 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Langworthy NG, Renz A, Mackenstedt U, Henkle–Dührsen K, Bronsvoort MBdC, Tanya VN, et al. Macrofilaricidal activity of tetracycline against the filarial nematode Onchocerca ochengi: elimination of Wolbachia precedes worm death and suggests a dependent relationship. Proceedings of the Royal Society of London B: Biological Sciences. 2000; 267(1448):1063–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nfon CK, Makepeace BL, Njongmeta LM, Tanya VN, Bain O, Trees AJ. Eosinophils contribute to killing of adult Onchocerca ochengi within onchocercomata following elimination of Wolbachia. Microbes and infection. 2006; 8(12):2698–705. [DOI] [PubMed] [Google Scholar]
- 15.Taylor MJ, Hoerauf A, Bockarie M. Lymphatic filariasis and onchocerciasis. The Lancet. 2010; 376(9747):1175–85. [DOI] [PubMed] [Google Scholar]
- 16.Shovlin JP. Doxy: use with caution: here are the things we must consider prior to prescribing therapy for rosacea. Review of Optometry. 2011; 148(5):99–100. [Google Scholar]
- 17.Townson S, Hutton D, Siemienska J, Hollick L, Scanlon T, Tagboto S, et al. Antibiotics and Wolbachia in filarial nematodes: antifilarial activity of rifampicin, oxytetracycline and chloramphenicol against Onchocerca gutturosa, Onchocerca lienalis and Brugia pahangi. Annals of tropical medicine and parasitology. 2000; 94(8):801–16. 10.1080/00034980020027988 [DOI] [PubMed] [Google Scholar]
- 18.Cline BL, Hernandez JL, Mather FJ, Bartholomew R, De Maza SN, Rodulfo S, et al. Albendazole in the treatment of onchocerciasis: double-blind clinical trial in Venezuela. The American journal of tropical medicine and hygiene. 1992; 47(4):512–20. 10.4269/ajtmh.1992.47.512 [DOI] [PubMed] [Google Scholar]
- 19.deC Bronsvoort BM, Makepeace BL, Renz A, Tanya VN, Fleckenstein L, Ekale D, et al. UMF-078: A modified flubendazole with potent macrofilaricidal activity against Onchocerca ochengi in African cattle. Parasites Vectors. journal article]. 2008. June 20; 1(1):18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mackenzie CD, Geary TG. Flubendazole: a candidate macrofilaricide for lymphatic filariasis and onchocerciasis field programs. Expert Rev Anti Infect Ther. 2011. May; 9(5):497–501. 10.1586/eri.11.30 [DOI] [PubMed] [Google Scholar]
- 21.Janssen Discontinues Development of Flubendazole Formulation to Treat Onchocerciasis. TITUSVILLE2017 [cited 2018 3/27/ 2018]; Available from: https://www.jnj.com/media-center/press-releases/janssen-discontinues-development-of-flubendazole-formulation-to-treat-onchocerciasis.
- 22.Fischer C, Ibiricu Urriza I, Bulman CA, Lim KC, Gut J, Lachau-Durand S, et al. Efficacy of subcutaneous doses and a new oral amorphous solid dispersion formulation of flubendazole on male jirds (Meriones unguiculatus) infected with the filarial nematode Brugia pahangi. PLoS neglected tropical diseases. 2019; 13(1):e0006787 10.1371/journal.pntd.0006787 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cupp E, Sauerbrey M, Richards F. Elimination of human onchocerciasis: history of progress and current feasibility using ivermectin (Mectizan) monotherapy. Acta tropica. 2011; 120:S100–S8. 10.1016/j.actatropica.2010.08.009 [DOI] [PubMed] [Google Scholar]
- 24.Halliday A, Guimaraes A, Tyrer H, Metuge H, Patrick C, Arnaud K-O, et al. A murine macrofilaricide pre-clinical screening model for onchocerciasis and lymphatic filariasis. Parasites Vectors. 2014. 2014/10/24; 7(1):1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Morris CP, Evans H, Larsen SE, Mitre E. A Comprehensive, Model-Based Review of Vaccine and Repeat Infection Trials for Filariasis. Clinical Microbiology Reviews. 2013; 26(3):381 10.1128/CMR.00002-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mutafchiev Y, Bain O, Williams Z, McCall JW, Michalski ML. Intraperitoneal development of the filarial nematode Brugia malayi in the Mongolian jird (Meriones unguiculatus). Parasitology research. 2014; 113(5):1827–35. 10.1007/s00436-014-3829-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Globisch D, Specht S, Pfarr KM, Eubanks LM, Hoerauf A, Janda KD. Litomosoides sigmodontis: A jird urine metabolome study. Bioorganic & medicinal chemistry letters. 2015; 25(24):5804–7. [DOI] [PubMed] [Google Scholar]
- 28.Bulman CA, Bidlow CM, Lustigman S, Cho-Ngwa F, Williams D, Rascón AA Jr, et al. Repurposing auranofin as a lead candidate for treatment of lymphatic filariasis and onchocerciasis. PLoS neglected tropical diseases. 2015; 9(2):e0003534 10.1371/journal.pntd.0003534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hardin J, Olson M, Gall D. Pathophysiology of small intestinal malabsorption in gerbils infected with G. lamblia. Gastroenterology. 1992; 103:506–13. [DOI] [PubMed] [Google Scholar]
- 30.Cheever AW. A Comparative Study of Schistosoma mansoni Infections in Mice, Gerbils, Multimammate Rats and Hamsters. I. The Relation of Portal Hypertension to Size of Hepatic Granulomas. American Journal of Tropical Medicine and Hygiene. 1965; 14(2):211–26. [DOI] [PubMed] [Google Scholar]
- 31.Taylor MJ, von Geldern TW, Ford L, Hübner MP, Marsh K, Johnston KL, et al. Preclinical development of an oral anti-Wolbachia macrolide drug for the treatment of lymphatic filariasis and onchocerciasis. Science translational medicine. 2019; 11(483):eaau2086 10.1126/scitranslmed.aau2086 [DOI] [PubMed] [Google Scholar]
- 32.Cho-Ngwa F, Abongwa M, Ngemenya MN, Nyongbela KD. Selective activity of extracts of Margaritaria discoidea and Homalium africanum on Onchocerca ochengi. BMC complementary and alternative medicine. 2010; 10(1):62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Devaney E, Howells RE. The microfilaricidal activity of ivermectin in vitro and in vivo. Tropenmed Parasitol. 1984. March; 35(1):47–9. [PubMed] [Google Scholar]
- 34.Carithers DS. Examining the role of macrolides and host immunity in combatting filarial parasites. Parasites Vectors. 2017; 10(1):182–. 10.1186/s13071-017-2116-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Maclean MJ, Savadelis MD, Coates R, Dzimianski MT, Jones C, Benbow C, et al. Does evaluation of in vitro microfilarial motility reflect the resistance status of Dirofilaria immitis isolates to macrocyclic lactones? Parasit Vectors. 2017. November 9; 10(Suppl 2):480 10.1186/s13071-017-2436-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Abraham D, Weiner DJ, Farrell JP. Cellular and humoral immune responses of jirds resistant to Dipetalonema viteae infection. Infect Immun. 1986. June; 52(3):742–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Arumugam S, Wei J, Liu Z, Abraham D, Bell A, Bottazzi ME, et al. Vaccination of Gerbils with Bm-103 and Bm-RAL-2 Concurrently or as a Fusion Protein Confers Consistent and Improved Protection against Brugia malayi Infection. PLoS neglected tropical diseases. 2016; 10(4):e0004586 10.1371/journal.pntd.0004586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Faye Schrater A, Spielman A, Piessens WF. Predisposition to Brugia Malayi Microfilaremia in Progeny of Infected Gerbils*. The American Journal of Tropical Medicine and Hygiene. 1983; 32(6):1306–8. 10.4269/ajtmh.1983.32.1306 [DOI] [PubMed] [Google Scholar]
- 39.Graham AL, Taylor MD, Le Goff L, Lamb TJ, Magennis M, Allen JE. Quantitative appraisal of murine filariasis confirms host strain differences but reveals that BALB/c females are more susceptible than males to Litomosoides sigmodontis. Microbes Infect. 2005. April; 7(4):612–8. 10.1016/j.micinf.2004.12.019 [DOI] [PubMed] [Google Scholar]
- 40.Rajan TV, Nelson FK, Cupp E, Schultz LD, Greiner DL. Survival of Onchocerca volvulus in nodules implanted in immunodeficient rodents. J Parasitol. 1992. February; 78(1):160–3. [PubMed] [Google Scholar]
- 41.Armstrong SD, Xia D, Bah GS, Krishna R, Ngangyung HF, LaCourse EJ, et al. Stage-specific Proteomes from Onchocerca ochengi, Sister Species of the Human River Blindness Parasite, Uncover Adaptations to a Nodular Lifestyle. Mol Cell Proteomics. 2016. August; 15(8):2554–75. 10.1074/mcp.M115.055640 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.McKellar QA, Scott EW. The benzimidazole anthelmintic agents-a review. Journal of Veterinary Pharmacology and Therapeutics. 1990; 13(3):223–47. [DOI] [PubMed] [Google Scholar]
- 43.Borgers M, De Nollin S. Ultrastructural changes in Ascaris suum intestine after mebendazole treatment in vivo. J Parasitol. 1975. February; 61(1):110–22. [PubMed] [Google Scholar]
- 44.Driscoll M, Dean E, Reilly E, Bergholz E, Chalfie M. Genetic and molecular analysis of a Caenorhabditis elegans beta-tubulin that conveys benzimidazole sensitivity. J Cell Biol. 1989. December; 109(6 Pt 1):2993–3003. 10.1083/jcb.109.6.2993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.deC Bronsvoort BM, Makepeace BL, Renz A, Tanya VN, Fleckenstein L, Ekale D, et al. UMF-078: A modified flubendazole with potent macrofilaricidal activity against Onchocerca ochengi in African cattle. Parasites Vectors. 2008; 1:18–. 10.1186/1756-3305-1-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Voronin D, Tricoche N, Jawahar S, Shlossman M, Bulman CA, Fischer C, et al. Development of a preliminary in vitro drug screening assay based on a newly established culturing system for pre-adult fifth-stage Onchocerca volvulus worms. PLoS neglected tropical diseases. 2019; 13(1):e0007108 10.1371/journal.pntd.0007108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.An G, Murry DJ, Gajurel K, Bach T, Deye G, Stebounova LV, et al. Pharmacokinetics, Safety, and Tolerability of Oxfendazole in Healthy Volunteers: a Randomized, Placebo-Controlled First-in-Human Single-Dose Escalation Study. Antimicrobial Agents and Chemotherapy. 2019:AAC.02255-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
All free worms were scored for motility before the nodular ones. The general motility score was similar in all the worms (almost always 100%). Gerbil SN 4 and 5 clearly illustrate this with 100% motility of all worms recovered, whether free or from nodules (p = 0.0931).
(PPTX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.