
Fig. 1.
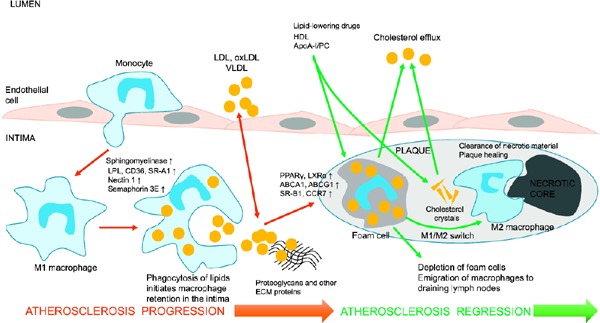
Atherosclerosis progression and reversal by enhancing cholesterol efflux and emigration of macrophages from the plaque
The proinflammatory recruitment of monocytes is followed by their subendothelial trafficking to the arterial intima, where monocytes differentiate into proinflammatory macrophages. The macrophages phagocytize proatherogenic low-density lipoprotein (LDL), oxidized LDL (oxLDL), and very low-density lipoprotein enriched with cholesterol. The accumulation of lipids in macrophages leads to their loss of mobility, retention in the vascular wall, and transformation to foam cells. Foam cells contribute to the formation of the intraplaque lipid pool and then the necrotic core. The increased production of matrix metalloproteinases (MMPs) by foam cells and plaque macrophages leads to plaque destabilization and rupture. Potent improvements in plasma lipoprotein levels by lowering LDL cholesterol and increasing high-density lipoprotein cholesterol can induce plaque regression, characterized by the enhancement of the reverse cholesterol transport, reduction of foam cell numbers, macrophage emigration, and phenotypic switch of retained macrophages from proinflammatory cells to anti-inflammatory cells that deal with the clearance of necrotic debris and plaque material and tissue repair. The increased mobility of macrophages is associated with up-regulation of liver X receptor and peroxisome proliferator-activated receptor gamma (PPARγ), which mediate cholesterol efflux in macrophages, activation of anti-inflammatory genes and cell mobility genes such as C-C chemokine receptor 7 (CCR7), and down-regulation of genes that inhibit cell migration such as semaphore in 3E and netrin 1.
Abbreviations: ABCA1, ATP-binding cassette transporter A1; ECM, extracellular matrix; LPL, lipoprotein lipase; PC, phosphatidylcholine; SR-B1, scavenger receptor.
Reprinted from “The phenomenon of atherosclerosis reversal and regression”: Lessons from animal models, Experimental and Molecular Pathology 102 (2017) 138–145. Copyright (2017), with permission from Elsevier “48