
Figure 5.
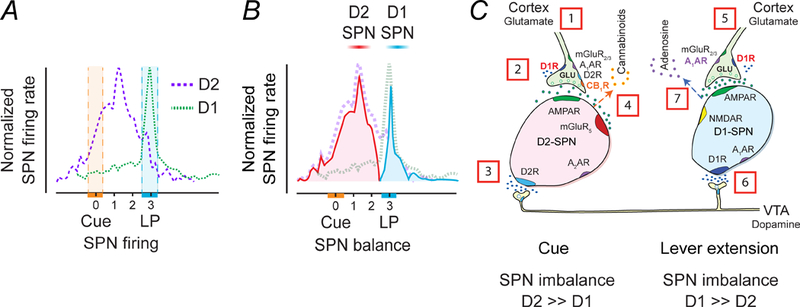
Time course of D1-SPN and D2-SPN responses in the rat NAc during an ICSS trial consisting of a cue presentation followed two seconds later by lever extension and then a subsequent lever press (LP), which delivers an electrical stimulation to dopamine cell bodies in the VTA. (A) Time course of D2-SPNs that are cue excitatory and D1-SPNs that are LP-excitatory. The hatched areas indicate the time variation of either cue or LP. (B) Changes in the balance between D2- and D1-SPNs during the cue and the LP. The cue temporarily heightens D2R-SPN activity to create an imbalance in striatal output. The imbalance may alert the animal, perhaps by promoting an urge to move. The lever extension increases D1R-SPN activity, culminating in the lever press (LP). (C) Temporal sequence of chemical activity at SPN synapses. The resting activity of SPNs is low because mature SPNs have prominent potassium currents (Wilson and Kawaguchi 1996) and cortical inputs to D2-SPNs are tonically inhibited by endocannabinoids (Wang and others 2012). [1] The sensory cue triggers excitatory inputs to D2-SPNs, which [2] activate further via presynaptic D1Rs in response to [3] dopamine release. [4] Balance is restored following the release of endocannabinoids from D2-SPNs. [5] In animals that have learned to associate sensory cue to the lever press, another set of glutamatergic inputs subsequently activate with [6] convergent dopamine release to stimulate D1-SPNs. Coactivation of post-synaptic D1Rs, NMDA and AMPA receptors on D1-SPNs release [7] adenosine to inhibit presynaptic activity to both D1- and D2-SPNs and rebalances the circuits.